
The Probiotic Bacillus subtilis MB40 Improves Immunity in a Porcine Model of Listeriosis

 ,
,  ,
,
Abstract
:1. Introduction
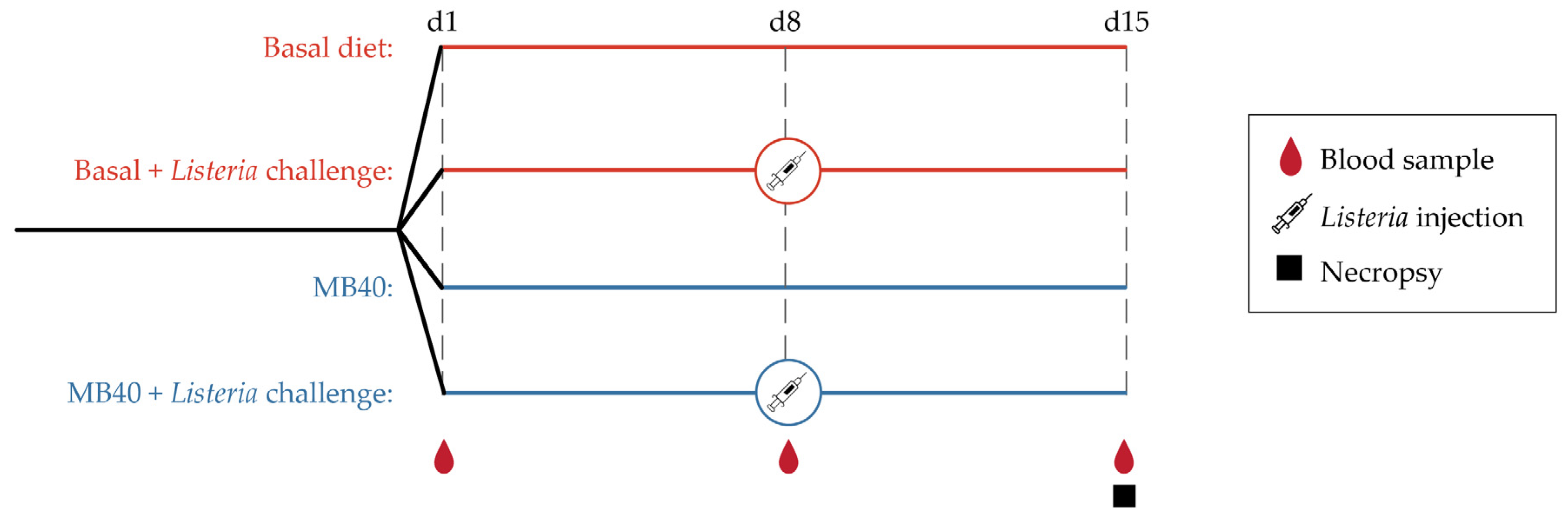
2. Materials and Methods
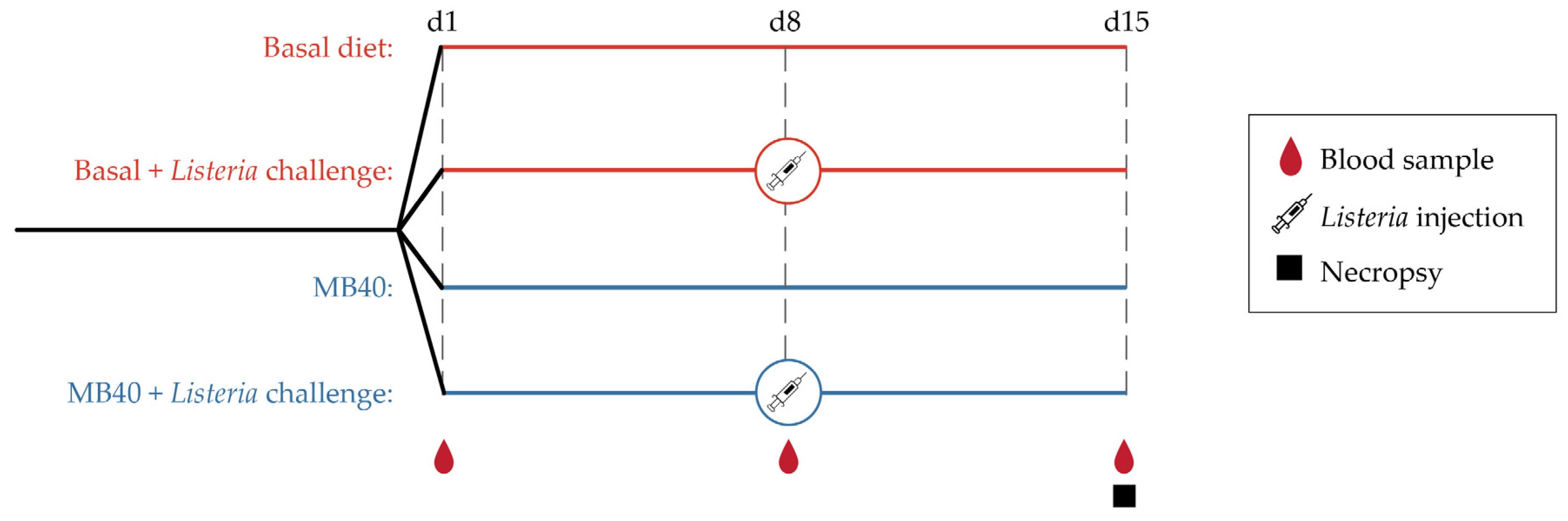
2.1. Animals and Diets
2.2. Bacterial Challenge
2.3. Necropsy and Sample Collection
2.4. Plasma Cytokine Analysis
2.5. Intestinal Morphology
2.6. Statistical Analysis
3. Results
3.1. Growth Parameters
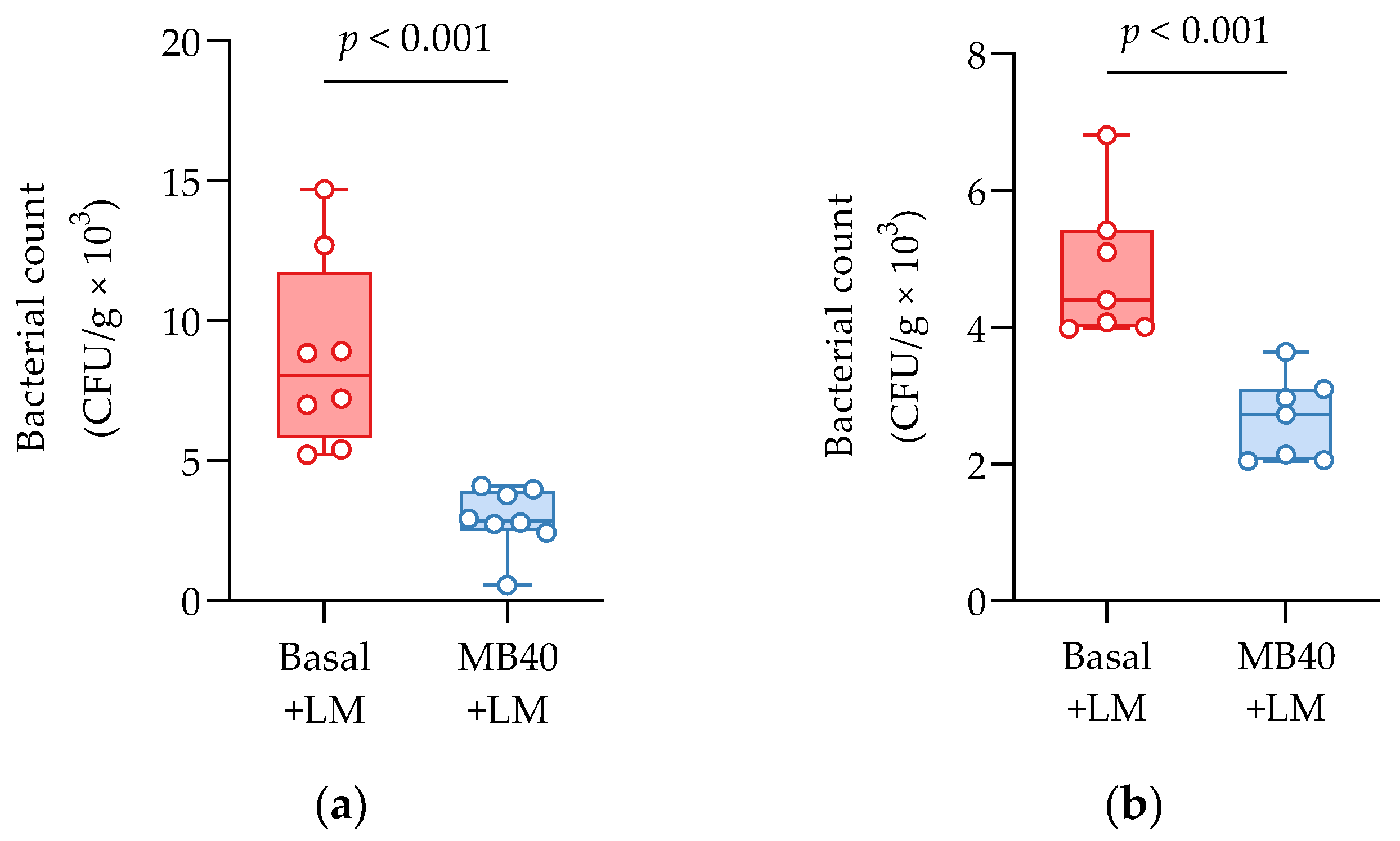
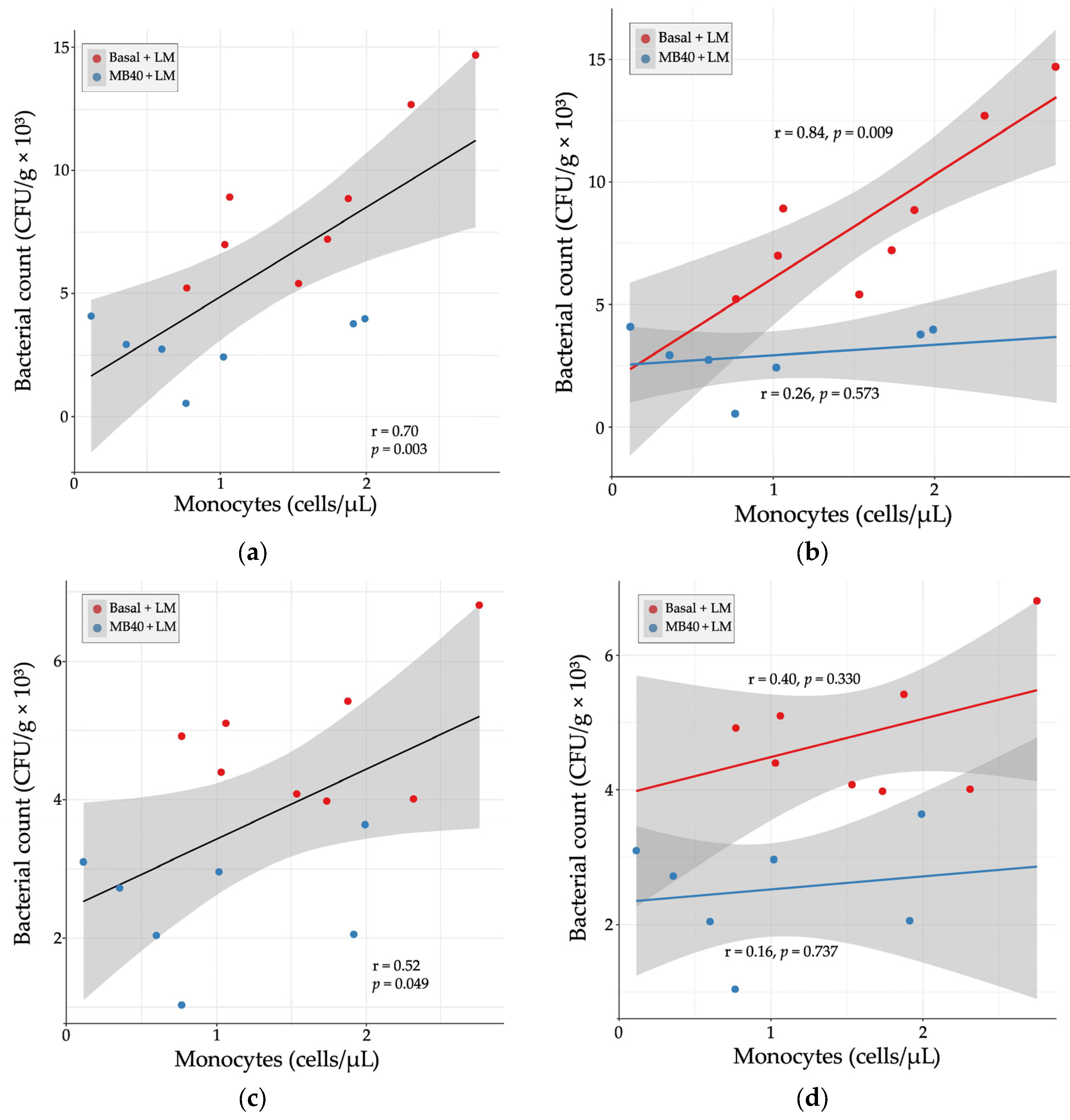
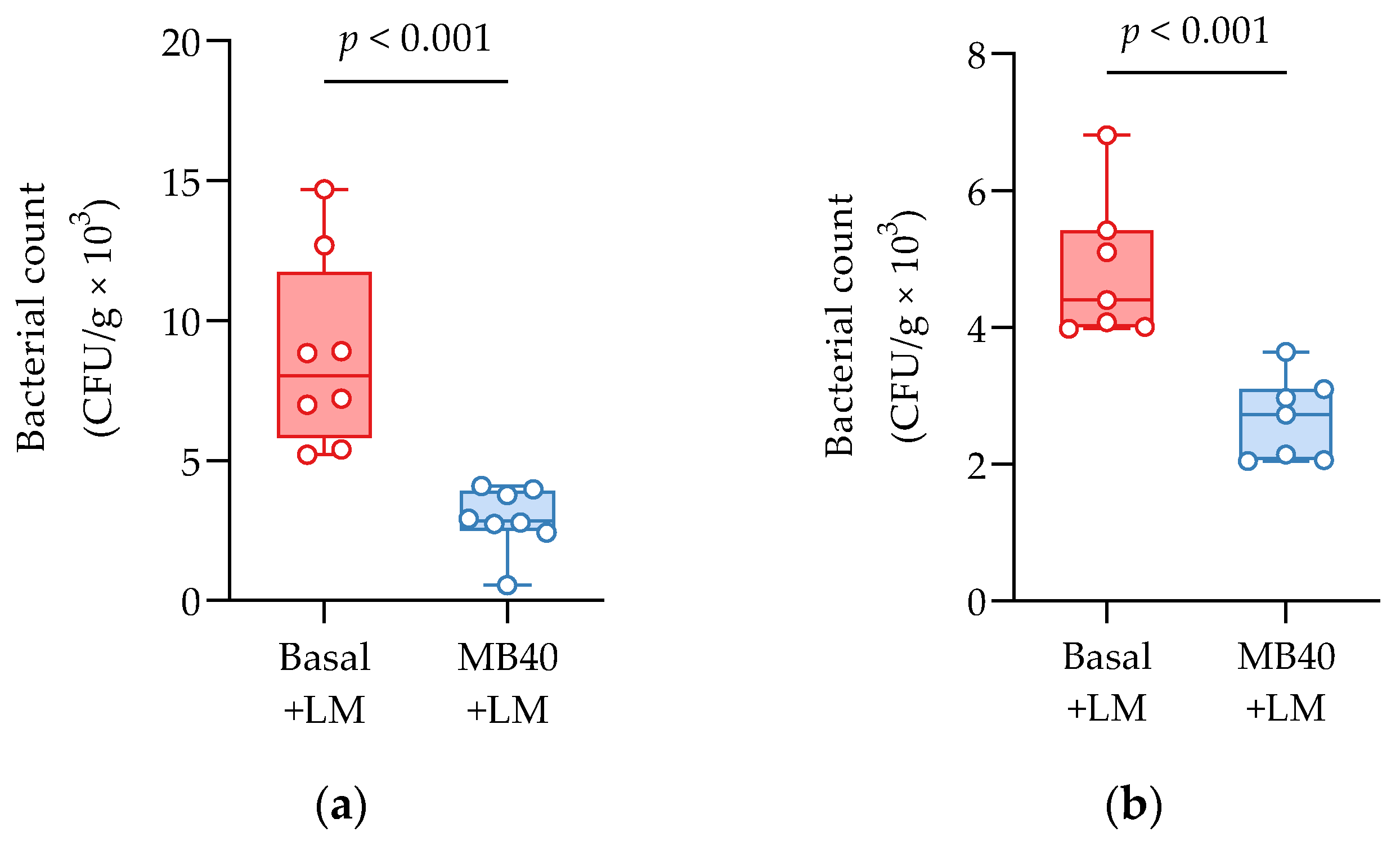
3.2. Organ Weights and Bacterial Counts
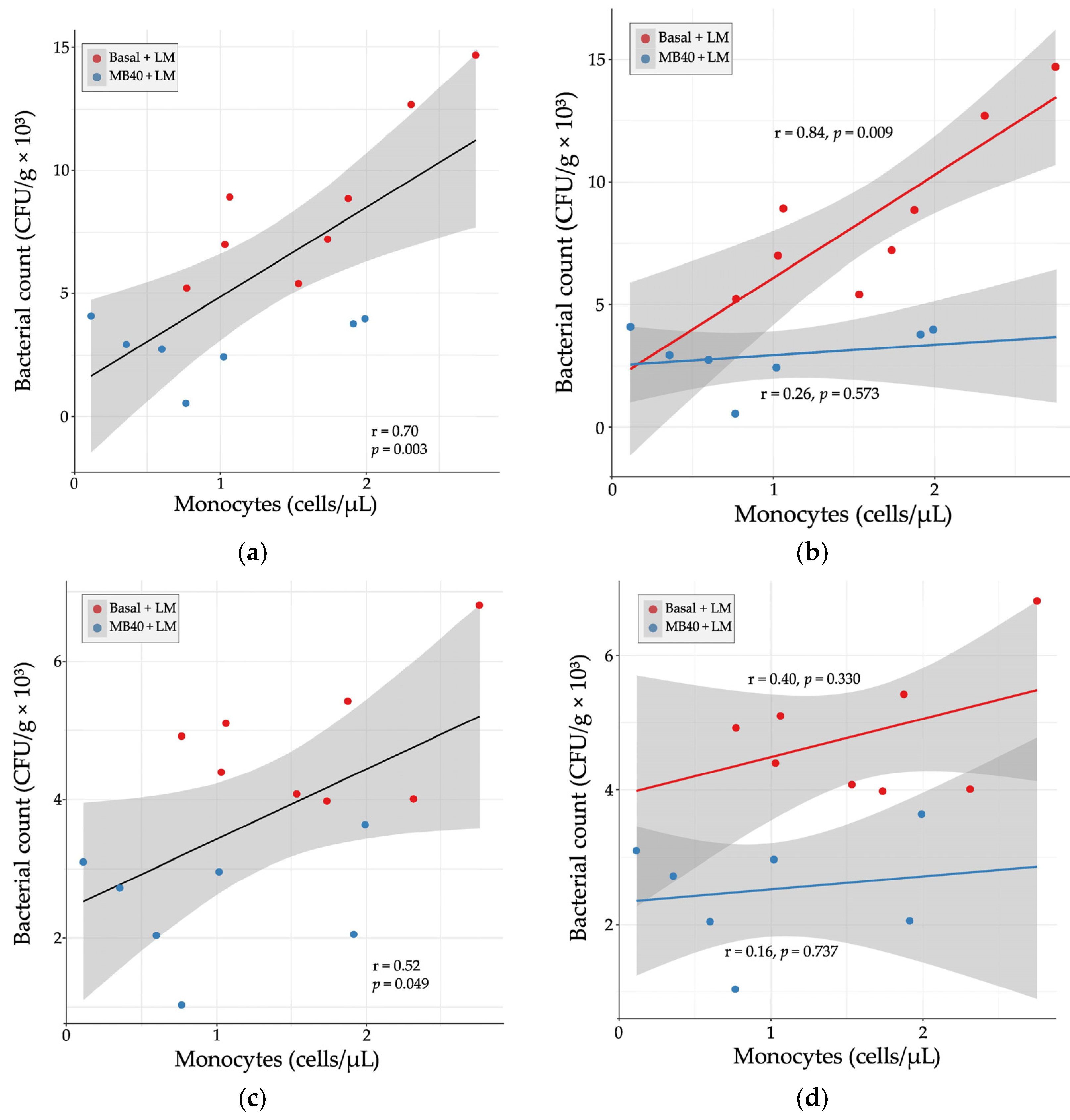
3.3. Hematology and Plasma Cytokines

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | p Value | ||||||
|---|---|---|---|---|---|---|---|
| Parameters (Units) | Basal | Basal + LM | MB40 | MB40 + LM | Diet | Challenge | D × C |
| Erythrocytes (cells/µL) * | |||||||
| d1 | 6.7 ± 0.62 | 6.6 ± 0.69 | 6.5 ± 0.42 | 6.8 ± 0.39 | 0.926 | 0.593 | 0.331 |
| d8 | 6.3 ± 0.66 | 6.2 ± 0.88 | 6.0 ± 0.78 | 6.5 ± 0.37 | 0.880 | 0.502 | 0.334 |
| d15 | 6.5 ± 0.52 | 6.3 ± 0.68 | 6.3 ± 0.53 | 6.5 ± 0.59 | 0.796 | 0.964 | 0.429 |
| Hemoglobin (g/dL) * | |||||||
| d1 | 12.0 ± 0.80 | 11.9 ± 0.91 | 11.9 ± 0.91 | 12.6 ± 0.66 | 0.358 | 0.437 | 0.207 |
| d8 | 10.9 ± 0.50 | 11.1 ± 1.50 | 10.6 ± 1.39 | 11.5 ± 0.38 | 0.847 | 0.191 | 0.415 |
| d15 | 11.4 ± 0.59 | 11.3 ± 0.56 | 11.2 ± 0.92 | 11.3 ± 0.92 | 0.674 | 0.971 | 0.605 |
| Hematocrit (%) * | |||||||
| d1 | 39.1 ± 2.90 | 38.3 ± 2.56 | 38.9 ± 2.91 | 40.7 ± 2.38 | 0.289 | 0.610 | 0.205 |
| d8 | 35.6 ± 2.11 | 36.0 ± 4.89 | 34.5 ± 4.06 | 37.0 ± 1.22 | 0.989 | 0.275 | 0.444 |
| d15 | 37.2 ± 1.73 | 35.8 ± 3.28 | 35.4 ± 2.81 | 36.9 ± 2.62 | 0.721 | 0.985 | 0.176 |
| Packed cell volume (%) * | |||||||
| d1 | 37.1 ± 1.66 | 36.7 ± 2.81 | 37.8 ± 3.17 | 39.0 ± 2.22 | 0.132 | 0.658 | 0.376 |
| d8 | 35.1 ± 3.40 | 35.3 ± 4.41 | 34.3 ± 5.16 | 36.0 ± 1.78 | 0.974 | 0.547 | 0.650 |
| d15 | 36.4 ± 1.74 | 33.8 ± 2.93 | 36.0 ± 3.50 | 35.6 ± 2.39 | 0.508 | 0.146 | 0.297 |
| Leukocytes (cells/µL) ‡ | |||||||
| d1 ‡ | 15.0 ± 1.98 A | 15.5 ± 3.10 B | 12.4 ± 2.29 A | 16.8 ± 3.95 B | 0.564 | 0.047 | 0.104 |
| d8 | 18.4 ± 5.13 | 17.7 ± 6.39 | 15.1 ± 3.79 | 16.6 ± 2.68 | 0.233 | 0.835 | 0.561 |
| d15 ‡ | 13.6 ± 3.35 A | 16.9 ± 3.03 B | 12.0 ± 3.22 A | 17.2 ± 4.93 B | 0.647 | 0.005 | 0.480 |
| Segmented neutrophils (cells/µL) *,‡ | |||||||
| d1 | 7.3 ± 2.32 | 9.1 ± 2.54 | 6.6 ± 2.58 | 8.9 ± 3.45 | 0.689 | 0.062 | 0.806 |
| d8 | 9.4 ± 4.20 | 10.4 ± 4.92 | 7.2 ± 3.39 | 9.1 ± 1.35 | 0.242 | 0.313 | 0.773 |
| d15 ‡ | 4.6 ± 1.70 A | 8.2 ± 2.33 B | 5.5 ± 1.03 A | 7.5 ± 2.45 B | 0.921 | 0.001 | 0.285 |
| Lymphocytes (cells/µL) | |||||||
| d1 | 6.5 ± 1.83 | 5.1 ± 1.00 | 4.9 ± 1.35 | 5.0 ± 1.46 | 0.159 | 0.267 | 0.191 |
| d8 | 7.4 ± 1.20 | 6.1 ± 2.46 | 6.3 ± 1.70 | 6.0 ± 1.67 | 0.417 | 0.255 | 0.468 |
| d15 | 7.8 ± 1.93 | 6.7 ± 2.08 | 5.8 ± 3.17 | 7.0 ± 2.07 | 0.363 | 0.999 | 0.190 |
| Eosinophils (cells/µL) ** | |||||||
| d1 | 0.17 ± 0.23 | 0.04 ± 0.07 | 0.20 ± 0.16 | 0.26 ± 0.20 | 0.072 | 0.645 | 0.152 |
| d8 † | 0.21 ± 0.28 a | 0.07 ± 0.08 a | 0.17 ± 0.19 b | 0.25 ± 0.17 b | 0.040 | 0.690 | 0.369 |
| d15 | 0.16 ± 0.09 | 0.29 ± 0.26 | 0.11 ± 0.16 | 0.21 ± 0.18 | 0.350 | 0.130 | 0.825 |
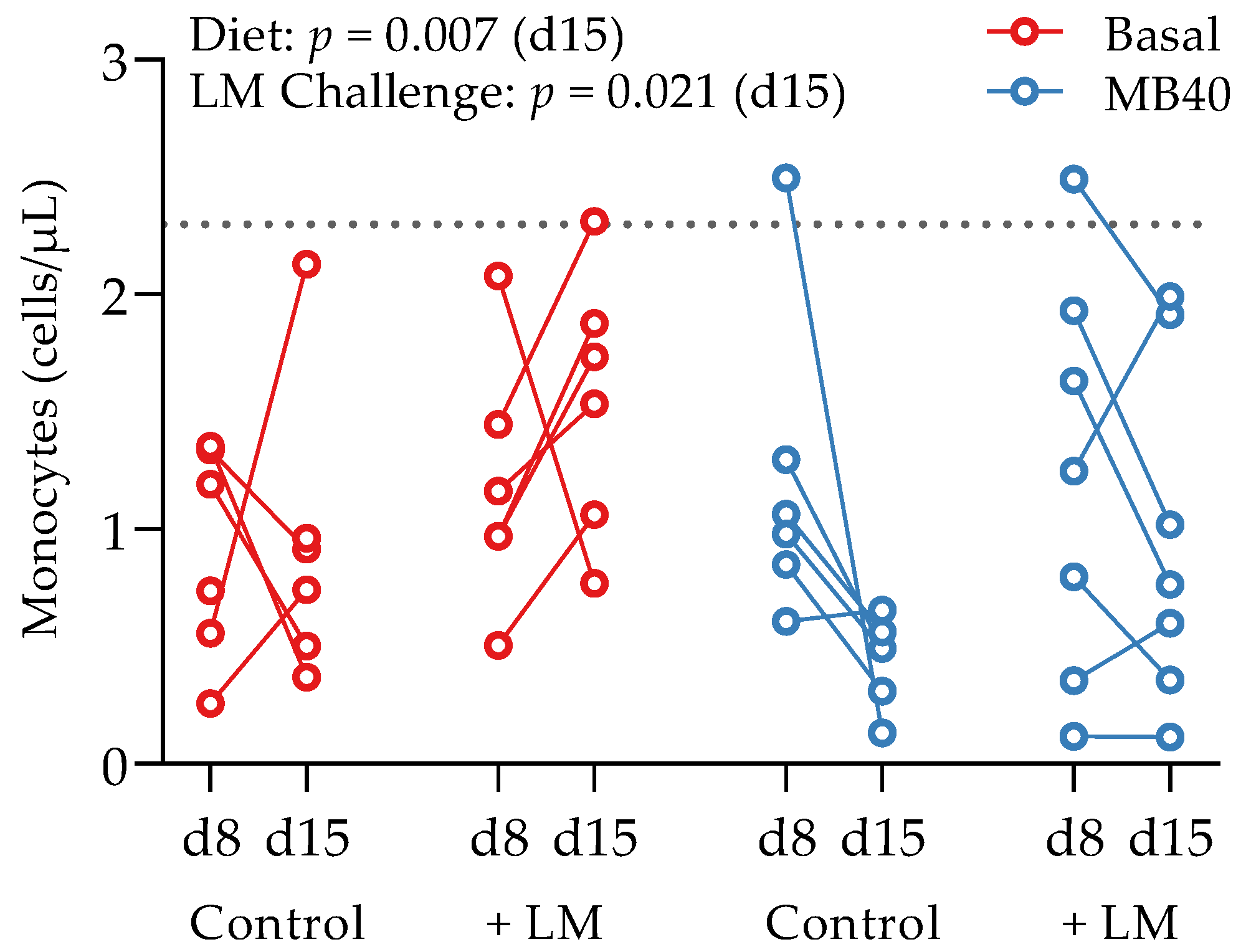
| Monocytes (cells/µL) ‡,** | |||||||
| d1 ‡ | 1.0 ± 0.43 A | 1.2 ± 0.65 B | 0.75 ± 0.29 A | 1.5 ± 0.72 B | 0.870 | 0.047 | 0.234 |
| d8 | 0.91 ± 0.46 | 1.2 ± 0.53 | 1.5 ± 0.88 | 1.2 ± 0.86 | 0.316 | 0.928 | 0.377 |
| d15 †,‡ | 0.89 ± 0.59 a,A | 1.6 ± 0.68 b,A | 0.44 ± 0.17 a,B | 0.97 ± 0.73 b,B | 0.007 | 0.021 | 0.801 |
| Interleukin-10 (pg/mL) * | |||||||
| d1 | 326 ± 104.4 | 401 ± 153.0 | 354 ± 148.1 | 361 ± 154.5 | 0.903 | 0.425 | 0.500 |
| d8 | 285 ± 73.6 | 343 ± 106.2 | 360 ± 66.5 | 317 ± 87.1 | 0.430 | 0.803 | 0.103 |
| d15 | 283 ± 70.1 | 235 ± 54.4 | 287 ± 62.0 | 276 ± 54.1 | 0.299 | 0.190 | 0.399 |
| TNF-α (pg/mL) * | |||||||
| d1 | 512 ± 156.1 | 595 ± 243.0 | 435 ± 102.0 | 452 ± 163.5 | 0.092 | 0.434 | 0.605 |
| d8 | 433 ± 220.8 | 410 ± 151.1 | 458 ± 177.5 | 330 ± 123.8 | 0.659 | 0.228 | 0.398 |
| d15 | 275 ± 123.5 | 337 ± 147.0 | 407 ± 194.0 | 403 ± 207.8 | 0.123 | 0.646 | 0.599 |
| Interleukin-6 (pg/mL) * | |||||||
| d1 | 736 ± 152.8 | 854 ± 152.6 | 756 ± 300.7 | 775 ± 281.6 | 0.726 | 0.409 | 0.551 |
| d8 | 336 ± 91.3 | 427 ± 171.7 | 439 ± 102.0 | 421 ± 137.7 | 0.299 | 0.435 | 0.242 |
| d15 | 505 ± 70.0 | 474 ± 141.2 | 504 ± 134.5 | 492 ± 106.3 | 0.837 | 0.608 | 0.818 |
3.4. Plasma Biochemistry
3.5. Intestinal Morphology
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert Consensus Document. The International Scientific Association for Probiotics and Prebiotics Consensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Probiotics and Prebiotics in Intestinal Health and Disease: From Biology to the Clinic. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 605–616. [Google Scholar] [CrossRef] [PubMed]
- Su, G.L.; Ko, C.W.; Bercik, P.; Falck-Ytter, Y.; Sultan, S.; Weizman, A.V.; Morgan, R.L. AGA Clinical Practice Guidelines on the Role of Probiotics in the Management of Gastrointestinal Disorders. Gastroenterology 2020, 159, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Goodman, C.; Keating, G.; Georgousopoulou, E.; Hespe, C.; Levett, K. Probiotics for the Prevention of Antibiotic-Associated Diarrhoea: A Systematic Review and Meta-Analysis. BMJ Open 2021, 11, e043054. [Google Scholar] [CrossRef]
- Liao, W.; Chen, C.; Wen, T.; Zhao, Q. Probiotics for the Prevention of Antibiotic-Associated Diarrhea in Adults: A Meta-Analysis of Randomized Placebo-Controlled Trials. J. Clin. Gastroenterol. 2021, 55, 469–480. [Google Scholar] [CrossRef]
- Pastor-Villaescusa, B.; Blanco-Rojo, R.; Olivares, M. Evaluation of the Effect of Limosilactobacillus fermentum CECT5716 on Gastrointestinal Infections in Infants: A Systematic Review and Meta-Analysis. Microorganisms 2021, 9, 1412. [Google Scholar] [CrossRef]
- Ahmad, H.H.; Peck, B.; Terry, D. The Influence of Probiotics on Gastrointestinal Tract Infections Among Children Attending Childcare: A Systematic Review and Meta-Analysis. J. Appl. Microbiol. 2022, 132, 1636–1651. [Google Scholar] [CrossRef]
- CDC, Centers for Disease Control and Prevention. Estimates of Foodborne Illness in the United States. Available online: https://www.cdc.gov/foodborneburden/2011-foodborne-estimates.html (accessed on 4 August 2023).
- Farber, J.M.; Peterkin, P.I. Listeria monocytogenes, a Food-Borne Pathogen. Microbiol. Rev. 1991, 55, 476–511. [Google Scholar] [CrossRef]
- Swaminathan, B.; Gerner-Smidt, P. The Epidemiology of Human Listeriosis. Microbes Infect. 2007, 9, 1236–1243. [Google Scholar] [CrossRef]
- Ogawa, M.; Yoshikawa, Y.; Mimuro, H.; Hain, T.; Chakraborty, T.; Sasakawa, C. Autophagy Targeting of Listeria monocytogenes and the Bacterial Countermeasure. Autophagy 2011, 7, 310–314. [Google Scholar] [CrossRef]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne Illness Acquired in the United States—Major Pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Ward, T.J.; Ducey, T.F.; Usgaard, T.; Dunn, K.A.; Bielawski, J.P. Multilocus Genotyping Assays for Single Nucleotide Polymorphism-Based Subtyping of Listeria monocytogenes Isolates. Appl. Environ. Microbiol. 2008, 74, 7629–7642. [Google Scholar] [CrossRef] [PubMed]
- Orsi, R.H.; den Bakker, H.C.; Wiedmann, M. Listeria monocytogenes Lineages: Genomics, Evolution, Ecology, and Phenotypic Characteristics. Int. J. Med. Microbiol. 2011, 301, 79–96. [Google Scholar] [CrossRef] [PubMed]
- Carstens, C.K.; Salazar, J.K.; Darkoh, C. Multistate Outbreaks of Foodborne Illness in the United States Associated with Fresh Produce from 2010 to 2017. Front. Microbiol. 2019, 10, 2667. [Google Scholar] [CrossRef] [PubMed]
- Cherifi, T.; Arsenault, J.; Pagotto, F.; Quessy, S.; Côté, J.C.; Neira, K.; Fournaise, S.; Bekal, S.; Fravalo, P. Distribution, Diversity and Persistence of Listeria monocytogenes in Swine Slaughterhouses and Their Association with Food and Human Listeriosis Strains. PLoS ONE 2020, 15, e0236807:1–e0236807:19. [Google Scholar] [CrossRef]
- Wiktorczyk-Kapischke, N.; Skowron, K.; Grudlewska-Buda, K.; Wałecka-Zacharska, E.; Korkus, J.; Gospodarek-Komkowska, E. Adaptive Response of Listeria monocytogenes to the Stress Factors in the Food Processing Environment. Front. Microbiol. 2021, 12, 710085. [Google Scholar] [CrossRef]
- Port, G.C.; Miner, M.D.; Freitag, N.E. Listeria monocytogenes–From Saprophyte to Intracellular Pathogen. Nat. Rev. Microbiol. 2009, 7, 623–628. [Google Scholar] [CrossRef]
- Jones, F.T.; Ricke, S.C. Observations on the History of the Development of Antimicrobials and Their Use in Poultry Feeds. Poult. Sci. 2003, 82, 613–617. [Google Scholar] [CrossRef]
- Dibner, J.J.; Richards, J.D. Antibiotic Growth Promoters in Agriculture: History and Mode of Action. Poult. Sci. 2005, 84, 634–643. [Google Scholar] [CrossRef]
- Castanon, J.I. History of the Use of Antibiotic as Growth Promoters in European Poultry Feeds. Poult. Sci. 2007, 86, 2466–2471. [Google Scholar] [CrossRef]
- EFSA, European Food Safety Authority. Foodborne Antimicrobial Resistance as a Biological Hazard–Scientific Opinion of the Panel on Biological Hazards. EFSA J. 2008, 765, 1–87. [Google Scholar] [CrossRef]
- FDA, Food and Drug Administration. Guide for Industry (GFI) #213. New Animal Drugs and New Animal Drug Combination Products Administered in or on Medicated Feed or Drinking Water of Food-Producing Animals: Recommendations for Drug Sponsors for Voluntarily Aligning Product Use Conditions with GFI #209. 2013. Available online: https://www.fda.gov/media/83488/download (accessed on 4 August 2023).
- Grant, A.; Gay, C.G.; Lillehoj, H.S. Bacillus spp. as Direct-Fed Microbial Antibiotic Alternatives to Enhance Growth, Immunity, and Gut Health in Poultry. Avian Pathol. 2018, 47, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Espinosa, C.D.; Abelilla, J.J.; Casas, G.A.; Lagos, L.V.; Lee, S.A.; Kwon, W.B.; Mathai, J.K.; Navarro, D.; Jaworski, N.W.; et al. Non-Antibiotic Feed Additives in Diets for Pigs: A Review. Anim. Nutr. 2018, 4, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Markowiak, P.; Śliżewska, K. The Role of Probiotics, Prebiotics and Synbiotics in Animal Nutrition. Gut Pathog. 2018, 10, 21. [Google Scholar] [CrossRef] [PubMed]
- Pollmann, D.S.; Danielson, D.M.; Peo, E.R., Jr. Effects of Microbial Feed Additives on Performance of Starter and Growing-Finishing Pigs. J. Anim. Sci. 1980, 51, 577–581. [Google Scholar] [CrossRef]
- Abe, F.; Ishibashi, N.; Shimamura, S. Effect of Administration of Bifidobacteria and Lactic Acid Bacteria to Newborn Calves and Piglets. J. Dairy Sci. 1995, 78, 2838–2846. [Google Scholar] [CrossRef]
- Alexopoulos, C.; Georgoulakis, I.E.; Tzivara, A.; Kyriakis, C.S.; Govaris, A.; Kyriakis, S.C. Field Evaluation of the Effect of a Probiotic-Containing Bacillus licheniformis and Bacillus subtilis Spores on the Health Status, Performance, and Carcass Quality of Grower and Finisher Pigs. J. Vet. Med. A Physiol. Pathol. Clin. Med. 2004, 51, 306–312. [Google Scholar] [CrossRef]
- Bahaddad, S.A.; Almalki, M.; Alghamdi, O.A.; Sohrab, S.S.; Yasir, M.; Azhar, E.I.; Chouayekh, H. Bacillus Species as Direct-Fed Microbial Antibiotic Alternatives for Monogastric Production. Probiotics Antimicrob. Proteins 2023, 15, 1–16. [Google Scholar] [CrossRef]
- Luise, D.; Bosi, P.; Raff, L.; Amatucci, L.; Virdis, S.; Trevisi, P. Bacillus spp. Probiotic Strains as a Potential Tool for Limiting the Use of Antibiotics, and Improving the Growth and Health of Pigs and Chickens. Front. Microbiol. 2022, 13, 801827. [Google Scholar] [CrossRef]
- Ilinskaya, O.N.; Ulyanova, V.V.; Yarullina, D.R.; Gataullin, I.G. Secretome of Intestinal Bacilli: A Natural Guard Against Pathologies. Front. Microbiol. 2018, 8, 1666. [Google Scholar] [CrossRef]
- Kaspar, F.; Neubauer, P.; Gimpel, M. Bioactive Secondary Metabolites from Bacillus subtilis: A Comprehensive Review. J. Nat. Prod. 2019, 82, 2038–2053. [Google Scholar] [CrossRef] [PubMed]
- Tran, C.; Cock, I.E.; Chen, X.; Feng, Y. Antimicrobial Bacillus: Metabolites and Their Mode of Action. Antibiotics 2022, 11, 88. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Qian, Y.; Yu, B.; Zhang, T.; Gao, J.; He, J.; Huang, Z.; Zheng, P.; Mao, X.; Luo, J.; et al. Effects of Bacillus subtilis DSM32315 Supplementation and Dietary Crude Protein Level on Performance, Gut Barrier Function and Microbiota Profile in Weaned Piglets. J. Anim. Sci. 2019, 97, 2125–2138. [Google Scholar] [CrossRef]
- Xu, Y.; Yu, Y.; Shen, Y.; Li, Q.; Lan, J.; Wu, Y.; Zhang, R.; Cao, G.; Yang, C. Effects of Bacillus subtilis and Bacillus licheniformis on Growth Performance, Immunity, Short Chain Fatty Acid Production, Antioxidant Capacity, and Cecal Microflora in Broilers. Poult. Sci. 2021, 100, 101358. [Google Scholar] [CrossRef]
- Huang, J.M.; La Ragione, R.M.; Nunez, A.; Cutting, S.M. Immunostimulatory Activity of Bacillus Spores. FEMS Immunol. Med. Microbiol. 2008, 53, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Li, Y.; Zhang, J.; Yang, Q. Co-administration of Bacillus subtilis RJGP16 and Lactobacillus salivarius B1 Strongly Enhances the Intestinal Mucosal Immunity of Piglets. Res. Vet. Sci. 2013, 94, 62–68. [Google Scholar] [CrossRef]
- Zhang, P.; Huang, L.; Zhang, E.; Yuan, C.; Yang, Q. Oral Administration of Bacillus subtilis Promotes Homing of CD3+ T Cells and IgA-Secreting Cells to the Respiratory Tract in Piglets. Res. Vet. Sci. 2021, 136, 310–317. [Google Scholar] [CrossRef]
- Hyronimus, B.; Le Marrec, C.; Sassi, A.H.; Deschamps, A. Acid and Bile Tolerance of Spore-Forming Lactic Acid Bacteria. Int. J. Food Microbiol. 2000, 61, 193–197. [Google Scholar] [CrossRef]
- Hong, H.A.; Duc, l.; Cutting, S.M. The Use of Bacterial Spore Formers as Probiotics. FEMS Microbiol. Rev. 2005, 29, 813–835. [Google Scholar] [CrossRef]
- Leser, T.D.; Knarreborg, A.; Worm, J. Germination and outgrowth of Bacillus subtilis and Bacillus licheniformis spores in the gastrointestinal tract of pigs. J. Appl. Microbiol. 2008, 104, 1025–1033. [Google Scholar] [CrossRef]
- Cutting, S.M. Bacillus Probiotics. Food Microbiol. 2011, 28, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Colom, J.; Freitas, D.; Simon, A.; Brodkorb, A.; Buckley, M.; Deaton, J.; Winger, A.M. Presence and Germination of the Probiotic Bacillus subtilis DE111® in the Human Small Intestinal Tract: A Randomized, Crossover, Double-Blind, and Placebo-Controlled Study. Front. Microbiol. 2021, 12, 715863. [Google Scholar] [CrossRef] [PubMed]
- Penet, C.; Kramer, R.; Little, R.; Spears, J.L.; Parker, J.; Iyer, J.K.; Guthrie, N.; Evans, M. A Randomized, Double-Blind, Placebo-Controlled, Parallel Study Evaluating the Efficacy of Bacillus subtilis MB40 to Reduce Abdominal Discomfort, Gas, and Bloating. Altern. Ther. Health Med. 2021, 27, 146–157. [Google Scholar] [PubMed]
- Garvey, S.M.; Mah, E.; Blonquist, T.M.; Kaden, V.N.; Spears, J.L. The Probiotic Bacillus subtilis BS50 Decreases Gastrointestinal Symptoms in Healthy Adults: A Randomized, Double-Blind, Placebo-Controlled Trial. Gut Microbes 2022, 14, 2122668. [Google Scholar] [CrossRef] [PubMed]
- Walden, K.E.; Hagele, A.M.; Orr, L.S.; Gross, K.N.; Krieger, J.M.; Jäger, R.; Kerksick, C.M. Probiotic BC30 Improves Amino Acid Absorption from Plant Protein Concentrate in Older Women. Probiotics Antimicrob. Proteins 2022, 1–13. [Google Scholar] [CrossRef]
- Dolin, B.J. Effects of a Proprietary Bacillus coagulans Preparation on Symptoms of Diarrhea-Predominant Irritable Bowel Syndrome. Methods Find. Exp. Clin. Pharmacol. 2009, 31, 655–659. [Google Scholar] [CrossRef]
- Hun, L. Bacillus coagulans Significantly Improved Abdominal Pain and Bloating in Patients with IBS. Postgrad. Med. 2009, 121, 119–124. [Google Scholar] [CrossRef]
- Kalman, D.S.; Schwartz, H.I.; Alvarez, P.; Feldman, S.; Pezzullo, J.C.; Krieger, D.R. A Prospective, Randomized, Double-Blind, Placebo-Controlled Parallel-Group Dual Site Trial to Evaluate the Effects of a Bacillus coagulans-Based Product on Functional Intestinal Gas Symptoms. BMC Gastroenterol. 2009, 9, 85. [Google Scholar] [CrossRef]
- Majeed, M.; Nagabhushanam, K.; Natarajan, S.; Sivakumar, A.; Ali, F.; Pande, A.; Majeed, S.; Karri, S.K. Bacillus coagulans MTCC 5856 Supplementation in the Management of Diarrhea Predominant Irritable Bowel Syndrome: A Double Blind Randomized Placebo Controlled Pilot Clinical Study. Nutr. J. 2016, 15, 21. [Google Scholar] [CrossRef]
- Hatanaka, M.; Yamamoto, K.; Suzuki, N.; Iio, S.; Takara, T.; Morita, H.; Takimoto, T.; Nakamura, T. Effect of Bacillus subtilis C-3102 on Loose Stools in Healthy Volunteers. Benef. Microbes 2018, 9, 357–365. [Google Scholar] [CrossRef]
- Madempudi, R.S.; Ahire, J.J.; Neelamraju, J.; Tripathi, A.; Nanal, S. Randomized Clinical Trial: The Effect of Probiotic Bacillus coagulans Unique IS2 vs. Placebo on the Symptoms Management of Irritable Bowel Syndrome in Adults. Sci. Rep. 2019, 9, 12210. [Google Scholar] [CrossRef] [PubMed]
- Maity, C.; Gupta, A.K. A Prospective, Interventional, Randomized, Double-Blind, Placebo-Controlled Clinical Study to Evaluate the Efficacy and Safety of Bacillus coagulans LBSC in the Treatment of Acute Diarrhea with Abdominal Discomfort. Eur. J. Clin. Pharmacol. 2019, 75, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Maity, C. Efficacy and Safety of Bacillus coagulans LBSC in Irritable Bowel Syndrome: A Prospective, Interventional, Randomized, Double-Blind, Placebo-Controlled Clinical Study [CONSORT Compliant]. Medicine 2021, 100, e23641. [Google Scholar] [CrossRef] [PubMed]
- Wauters, L.; Slaets, H.; De Paepe, K.; Ceulemans, M.; Wetzels, S.; Geboers, K.; Toth, J.; Thys, W.; Dybajlo, R.; Walgraeve, D.; et al. Efficacy and Safety of Spore-Forming Probiotics in the Treatment of Functional Dyspepsia: A Pilot Randomised, Double-Blind, Placebo-Controlled Trial. Lancet Gastroenterol. Hepatol. 2021, 6, 784–792. [Google Scholar] [CrossRef] [PubMed]
- Kyriakis, S.C.; Tsiloyiannis, V.K.; Vlemmas, J.; Sarris, K.; Tsinas, A.C.; Alexopoulos, C.; Jansegers, L. The Effect of Probiotic LSP 122 on the Control of Post-Weaning Diarrhoea Syndrome of Piglets. Res. Vet. Sci. 1999, 67, 223–228. [Google Scholar] [CrossRef]
- Kantas, D.; Papatsiros, V.G.; Tassis, P.D.; Giavasis, I.; Bouki, P.; Tzika, E.D. A Feed Additive Containing Bacillus toyonensis (Toyocerin®) Protects Against Enteric Pathogens in Postweaning Piglets. J. Appl. Microbiol. 2015, 118, 727–738. [Google Scholar] [CrossRef]
- Amalaradjou, M.A.; Bhunia, A.K. Modern Approaches in Probiotics Research to Control Foodborne Pathogens. Adv. Food Nutr. Res. 2012, 67, 185–239. [Google Scholar] [CrossRef]
- Becker, S.L.; Li, Q.; Burrough, E.R.; Kenne, D.; Sahin, O.; Gould, S.A.; Patience, J.F. Effects of an F18 Enterotoxigenic Escherichia coli Challenge on Growth Performance, Immunological Status, and Gastrointestinal Structure of Weaned Pigs and the Potential Protective Effect of Direct-Fed Microbial Blends. J. Anim. Sci. 2020, 98, skaa113. [Google Scholar] [CrossRef]
- He, Y.; Jinno, C.; Kim, K.; Wu, Z.; Tan, B.; Li, X.; Whelan, R.; Liu, Y. Dietary Bacillus spp. Enhanced Growth and Disease Resistance of Weaned Pigs by Modulating Intestinal Microbiota and Systemic Immunity. J. Anim. Sci. Biotechnol. 2020, 11, 101. [Google Scholar] [CrossRef]
- Sabaté, D.C.; Audisio, M.C. Inhibitory Activity of Surfactin, Produced by Different Bacillus subtilis subsp. subtilis Strains, Against Listeria monocytogenes Sensitive and Bacteriocin-Resistant Strains. Microbiol. Res. 2013, 168, 125–129. [Google Scholar] [CrossRef]
- Das, G.; Park, S.; Baek, K.H. Diversity of Endophytic Bacteria in a Fern Species Dryopteris uniformis (Makino) Makino and Evaluation of Their Antibacterial Potential Against Five Foodborne Pathogenic Bacteria. Foodborne Pathog. Dis. 2017, 14, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.Z.; Zheng, Q.W.; Wei, T.; Zhang, Z.Q.; Zhao, C.F.; Zhong, H.; Xu, Q.Y.; Lin, J.F.; Guo, L.Q. Isolation and Characterization of Fengycins Produced by Bacillus amyloliquefaciens JFL21 and Its Broad-Spectrum Antimicrobial Potential Against Multidrug-Resistant Foodborne Pathogens. Front. Microbiol. 2020, 11, 579621. [Google Scholar] [CrossRef] [PubMed]
- Zilelidou, E.A.; Milina, V.; Paramithiotis, S.; Zoumpopoulou, G.; Poimenidou, S.V.; Mavrogonatou, E.; Kletsas, D.; Papadimitriou, K.; Tsakalidou, E.; Skandamis, P.N. Differential Modulation of Listeria monocytogenes Fitness, In Vitro Virulence, and Transcription of Virulence-Associated Genes in Response to the Presence of Different Microorganisms. Appl. Environ. Microbiol. 2020, 86, e01165-20. [Google Scholar] [CrossRef] [PubMed]
- Hudson, L.K.; Orellana, L.; Bryan, D.W.; Moore, A.; Munafo, J.P.; den Bakker, H.C.; Denes, T.G. Phylogeny of the Bacillus altitudinis Complex and Characterization of a Newly Isolated Strain with Antilisterial Activity. J. Food Prot. 2021, 84, 1321–1332. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Woo, C.; Fugaban, J.; Vazquez Bucheli, J.E.; Holzapfel, W.H.; Todorov, S.D. Bacteriocinogenic Potential of Bacillus amyloliquefaciens Isolated from Kimchi, a Traditional Korean Fermented Cabbage. Probiotics Antimicrob. Proteins 2021, 13, 1195–1212. [Google Scholar] [CrossRef]
- Johny, L.C.; Suresh, P.V. Complete Genome Sequencing and Strain Characterization of a Novel Marine Bacillus velezensis FTL7 with a Potential Broad Inhibitory Spectrum Against Foodborne Pathogens. World J. Microbiol. Biotechnol. 2022, 38, 164. [Google Scholar] [CrossRef]
- Saggese, A.; De Luca, Y.; Baccigalupi, L.; Ricca, E. An Antimicrobial Peptide Specifically Active Against Listeria monocytogenes is Secreted by Bacillus pumilus SF214. BMC Microbiol. 2022, 22, 3. [Google Scholar] [CrossRef]
- Wang, H.; Jiang, X. Isolation and Characterization of Competitive Exclusion Microorganisms from Animal Wastes-Based Composts Against Listeria monocytogenes. J. Appl. Microbiol. 2022, 132, 4531–4543. [Google Scholar] [CrossRef]
- Spears, J.L.; Kramer, R.; Nikiforov, A.I.; Rihner, M.O.; Lambert, E.A. Safety Assessment of Bacillus subtilis MB40 for Use in Foods and Dietary Supplements. Nutrients 2021, 13, 733. [Google Scholar] [CrossRef]
- FDA. GRN No. 955. Bacillus subtilis Strain BS-MB40 PTA-122264 Spore Preparation. 2021. Available online: https://www.cfsanappsexternal.fda.gov/scripts/fdcc/index.cfm?set=GRASNotices&id=955 (accessed on 4 August 2023).
- National Research Council. Nutrient Requirements of Swine, 11th ed.; The National Academies Press: Washington, DC, USA, 2012. [Google Scholar] [CrossRef]
- Hof, H.; Hefner, P. Pathogenicity of Listeria monocytogenes in Comparison to Other Listeria Species. Infection 1988, 16, S141–S144. [Google Scholar] [CrossRef]
- Pine, L.; Malcolm, G.B.; Plikaytis, B.D. Listeria monocytogenes Intragastric and Intraperitoneal Approximate 50% Lethal Doses for Mice are Comparable, but Death Occurs Earlier by Intragastric Feeding. Infect. Immun. 1990, 58, 2940–2945. [Google Scholar] [CrossRef] [PubMed]
- Reference Intervals from the Clinical Pathology Laboratory, Department of Veterinary Pathology, College of Veterinary Medicine, Iowa State University. Available online: https://vetmed.iastate.edu/vpath/services/diagnostic-services/clinical-pathology/testing-and-fees/reference-intervals (accessed on 4 August 2023).
- Vázquez-Boland, J.A.; Kuhn, M.; Berche, P.; Chakraborty, T.; Domínguez-Bernal, G.; Goebel, W.; González-Zorn, B.; Wehland, J.; Kreft, J. Listeria Pathogenesis and Molecular Virulence Determinants. Clin. Microbiol. Rev. 2001, 14, 584–640. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Lee, S.N.; Chang, S.Y.; Ko, H.J.; Ryu, S.; Kweon, M.N. A Mouse Model of Shigellosis by Intraperitoneal Infection. J. Infect. Dis. 2014, 209, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Piewngam, P.; Zheng, Y.; Nguyen, T.H.; Dickey, S.W.; Joo, H.S.; Villaruz, A.E.; Glose, K.A.; Fisher, E.L.; Hunt, R.L.; Li, B.; et al. Pathogen Elimination by Probiotic Bacillus via Signalling Interference. Nature 2018, 562, 532–537. [Google Scholar] [CrossRef]
- Mercado, V.; Olmos, J. Bacteriocin Production by Bacillus Species: Isolation, Characterization, and Application. Probiotics Antimicrob. Proteins 2022, 14, 1151–1169. [Google Scholar] [CrossRef]
- Wang, J.; Xu, H.; Liu, S.; Song, B.; Liu, H.; Li, F.; Deng, S.; Wang, G.; Zeng, H.; Zeng, X.; et al. Toyoncin, a Novel Leaderless Bacteriocin That Is Produced by Bacillus toyonensis XIN-YC13 and Specifically Targets B. cereus and Listeria monocytogenes. Appl. Environ. Microbiol. 2021, 87, e00185-21. [Google Scholar] [CrossRef]
- Xin, B.; Xu, H.; Liu, H.; Liu, S.; Wang, J.; Xue, J.; Zhang, F.; Deng, S.; Zeng, H.; Zeng, X.; et al. Identification and Characterization of a Novel Circular Bacteriocin, Bacicyclicin XIN-1, from Bacillus sp. Xin1. Food Control 2021, 121, 107696. [Google Scholar] [CrossRef]
- Cruz Mendoza, I.; Villavicencio-Vasquez, M.; Aguayo, P.; Coello Montoya, D.; Plaza, L.; Romero-Peña, M.; Marqués, A.M.; Coronel-León, J. Biosurfactant from Bacillus subtilis DS03: Properties and Application in Cleaning Out Place System in a Pilot Sausages Processing. Microorganisms 2022, 10, 1518. [Google Scholar] [CrossRef]
- Deng, S.; Liu, S.; Li, X.; Liu, H.; Li, F.; Liu, K.; Zeng, H.; Zeng, X.; Xin, B. Thuricins: Novel Leaderless Bacteriocins with Potent Antimicrobial Activity Against Gram-Positive Foodborne Pathogens. J. Agric. Food Chem. 2022, 70, 9990–9999. [Google Scholar] [CrossRef]
- Hong, S.W.; Kim, J.H.; Cha, H.A.; Chung, K.S.; Bae, H.J.; Park, W.S.; Ham, J.S.; Park, B.Y.; Oh, M.H. Identification and Characterization of a Bacteriocin from the Newly Isolated Bacillus subtilis HD15 with Inhibitory Effects against Bacillus cereus. J. Microbiol. Biotechnol. 2022, 32, 1462–1470. [Google Scholar] [CrossRef]
- Stincone, P.; Fonseca Veras, F.; Micalizzi, G.; Donnarumma, D.; Vitale Celano, G.; Petras, D.; de Angelis, M.; Mondello, L.; Brandelli, A. Listeria monocytogenes Exposed to Antimicrobial Peptides Displays Differential Regulation of Lipids and Proteins Associated to Stress Response. Cell. Mol. Life Sci. 2022, 79, 263. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xin, N.; Zhu, Z.; Li, X.; Dai, D.; Pan, C.; Peng, D.; Sun, M. Three Novel Leaderless Bacteriocins Have Antimicrobial Activity Against Gram-Positive Bacteria to Serve as Promising Food Biopreservative. Microb. Cell Fact. 2022, 21, 194. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gu, Z.; Zhang, S.; Li, P. Complete Genome Sequencing Revealed the Potential Application of a Novel Weizmannia coagulans PL-W Production with Promising Bacteriocins in Food Preservative. Foods 2023, 12, 216. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Sha, C.; Zhang, L.; Ge, D.; Wang, Y.; Xia, X.; Liu, X.; Zhou, J. A Novel Subtilin-Like Lantibiotics Subtilin JS-4 Produced by Bacillus subtilis JS-4, and its Antibacterial Mechanism Against Listeria monocytogenes. LWT 2021, 142, 110993. [Google Scholar] [CrossRef]
- de Waard, R.; Garssen, J.; Bokken, G.C.; Vos, J.G. Antagonistic Activity of Lactobacillus casei Strain Shirota Against Gastrointestinal Listeria monocytogenes Infection in Rats. Int. J. Food Microbiol. 2002, 73, 93–100. [Google Scholar] [CrossRef]
- Corr, S.C.; Li, Y.; Riedel, C.U.; O’Toole, P.W.; Hill, C.; Gahan, C.G. Bacteriocin Production as a Mechanism for the Antiinfective Activity of Lactobacillus salivarius UCC118. Proc. Natl. Acad. Sci. USA 2007, 104, 7617–7621. [Google Scholar] [CrossRef]
- dos Santos, L.M.; Santos, M.M.; de Souza Silva, H.P.; Arantes, R.M.; Nicoli, J.R.; Vieira, L.Q. Monoassociation with Probiotic Lactobacillus delbrueckii UFV-H2b20 Stimulates the Immune System and Protects Germfree Mice Against Listeria monocytogenes Infection. Med. Microbiol. Immunol. 2011, 200, 29–38. [Google Scholar] [CrossRef]
- Archambaud, C.; Nahori, M.A.; Soubigou, G.; Bécavin, C.; Laval, L.; Lechat, P.; Smokvina, T.; Langella, P.; Lecuit, M.; Cossart, P. Impact of Lactobacilli on Orally Acquired Listeriosis. Proc. Natl. Acad. Sci. USA 2012, 109, 16684–16689. [Google Scholar] [CrossRef]
- Lukic, J.; Jancic, I.; Mirkovic, N.; Bufan, B.; Djokic, J.; Milenkovic, M.; Begovic, J.; Strahinic, I.; Lozo, J. Lactococcus lactis and Lactobacillus salivarius Differently Modulate Early Immunological Response of Wistar Rats Co-Administered with Listeria monocytogenes. Benef. Microbes 2017, 8, 809–822. [Google Scholar] [CrossRef]
- Li, Y.; Liu, M.; Zhou, J.; Hou, B.; Su, X.; Liu, Z.; Yuan, J.; Li, M. Bacillus licheniformis Zhengchangsheng® Attenuates DSS-Induced Colitis and Modulates the Gut Microbiota in Mice. Benef. Microbes 2019, 10, 543–553. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, J.; Cao, M.; Li, Y.; Zhuo, Y.; Fang, Z.; Che, L.; Xu, S.; Feng, B.; Lin, Y.; et al. Dietary Supplementation of Bacillus subtilis PB6 Improves Sow Reproductive Performance and Reduces Piglet Birth Intervals. Anim. Nutr. 2020, 6, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Heng, C.; Zhou, X.; Cao, G.; Jiang, L.; Wang, J.; Li, K.; Wang, D.; Zhan, X. Supplemental Bacillus subtilis DSM 29784 and Enzymes, Alone or in Combination, as Alternatives for Antibiotics to Improve Growth Performance, Digestive Enzyme Activity, Anti-Oxidative Status, Immune Response and the Intestinal Barrier of Broiler Chickens. Br. J. Nutr. 2021, 125, 494–507. [Google Scholar] [CrossRef]
- Richter, J.F.; Gitter, A.H.; Günzel, D.; Weiss, S.; Mohamed, W.; Chakraborty, T.; Fromm, M.; Schulzke, J.D. Listeriolysin O Affects Barrier Function and Induces Chloride Secretion in HT-29/B6 Colon Epithelial Cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G1350–G1359. [Google Scholar] [CrossRef] [PubMed]
- Cajnko, M.M.; Marušić, M.; Kisovec, M.; Rojko, N.; Benčina, M.; Caserman, S.; Anderluh, G. Listeriolysin O Affects the Permeability of Caco-2 Monolayer in a Pore-Dependent and Ca2+-Independent Manner. PLoS ONE 2015, 10, e0130471. [Google Scholar] [CrossRef] [PubMed]
- Tingting, W.; Tianqi, F.; Xinyu, W.; Can, Z.; Xue, S.; Xuming, D.; Jianfeng, W. Amentoflavone Attenuates Listeria monocytogenes Pathogenicity Through an LLO-Dependent Mechanism. Br. J. Pharmacol. 2022, 179, 3839–3858. [Google Scholar] [CrossRef]
- Albers, R.; Antoine, J.M.; Bourdet-Sicard, R.; Calder, P.C.; Gleeson, M.; Lesourd, B.; Samartín, S.; Sanderson, I.R.; Van Loo, J.; Vas Dias, F.W.; et al. Markers to Measure Immunomodulation in Human Nutrition Intervention Studies. Br. J. Nutr. 2005, 94, 452–481. [Google Scholar] [CrossRef]
- Fiorini, G.; Cimminiello, C.; Chianese, R.; Visconti, G.P.; Cova, D.; Uberti, T.; Gibelli, A. Bacillus subtilis Selectively Stimulates the Synthesis of Membrane Bound and Secreted IgA. Chemioterapia 1985, 4, 310–312. [Google Scholar]
- Marseglia, G.L.; Tosca, M.; Cirillo, I.; Licari, A.; Leone, M.; Marseglia, A.; Castellazzi, A.M.; Ciprandi, G. Efficacy of Bacillus clausii Spores in the Prevention of Recurrent Respiratory Infections in Children: A Pilot Study. Ther. Clin. Risk Manag. 2007, 3, 13–17. [Google Scholar] [CrossRef]
- Lefevre, M.; Racedo, S.M.; Ripert, G.; Housez, B.; Cazaubiel, M.; Maudet, C.; Jüsten, P.; Marteau, P.; Urdaci, M.C. Probiotic Strain Bacillus subtilis CU1 Stimulates Immune System of Elderly During Common Infectious Disease Period: A Randomized, Double-Blind Placebo-Controlled Study. Immun. Ageing 2015, 12, 24. [Google Scholar] [CrossRef]
- Burgueño, J.F.; Abreu, M.T. Epithelial Toll-like Receptors and Their Role in Gut Homeostasis and Disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 263–278. [Google Scholar] [CrossRef]
- Tian, Z.; Wang, X.; Duan, Y.; Zhao, Y.; Zhang, W.; Azad, M.; Wang, Z.; Blachier, F.; Kong, X. Dietary Supplementation with Bacillus subtilis Promotes Growth and Gut Health of Weaned Piglets. Front. Vet. Sci. 2021, 7, 600772. [Google Scholar] [CrossRef] [PubMed]
- Piewngam, P.; Khongthong, S.; Roekngam, N.; Theapparat, Y.; Sunpaweravong, S.; Faroongsarng, D.; Otto, M. Probiotic for Pathogen-Specific Staphylococcus aureus Decolonisation in Thailand: A Phase 2, Double-Blind, Randomised, Placebo-Controlled Trial. Lancet Gastroenterol. Hepatol. 2023, 4, e75–e83. [Google Scholar] [CrossRef] [PubMed]
| Condition | p Value | ||||||
|---|---|---|---|---|---|---|---|
| Parameter (Unit) | Basal | Basal + LM | MB40 | MB40 + LM | Diet | Challenge | D × C |
| Body weight (kg) * | |||||||
| d1 | 7.46 ± 1.67 | 6.96 ± 1.97 | 6.92 ± 1.64 | 7.16 ± 1.78 | 0.785 | 0.840 | 0.561 |
| d8 | 9.07 ± 2.01 | 8.60 ± 2.49 | 8.74 ± 2.03 | 8.96 ± 2.90 | 0.985 | 0.881 | 0.692 |
| d15 | 12.83 ± 2.92 | 12.04 ± 3.37 | 11.94 ± 2.98 | 11.98 ± 3.30 | 0.672 | 0.739 | 0.717 |
| Body weight gain (kg) * | |||||||
| d1–d8 | 1.61 ± 0.67 | 1.64 ± 1.10 | 1.83 ± 0.70 | 1.80 ± 1.21 | 0.580 | 0.999 | 0.929 |
| d8–d15 | 3.76 ± 1.87 | 3.44 ± 1.46 | 3.20 ± 1.11 | 3.02 ± 0.64 | 0.311 | 0.608 | 0.885 |
| d1–d15 | 5.37 ± 2.24 | 5.08 ± 1.50 | 5.03 ± 1.51 | 4.82 ± 1.66 | 0.628 | 0.693 | 0.950 |
| ADG (kg) * | |||||||
| d1–d8 | 0.18 ± 0.07 | 0.20 ± 0.11 | 0.19 ± 0.12 | 0.17 ± 0.09 | 0.727 | 0.963 | 0.647 |
| d8–d15 | 0.18 ± 0.07 | 0.18 ± 0.12 | 0.20 ± 0.08 | 0.20 ± 0.13 | 0.579 | 0.995 | 0.929 |
| d1–d15 | 0.42 ± 0.21 | 0.38 ± 0.16 | 0.36 ± 0.12 | 0.34 ± 0.07 | 0.309 | 0.608 | 0.886 |
| ADFI (kg) * | |||||||
| d1–d8 | 0.50 ± 0.14 | 0.54 ± 0.17 | 0.45 ± 0.13 | 0.47 ± 0.18 | 0.313 | 0.253 | 0.919 |
| d8–d15 | 0.81 ± 0.16 | 0.76 ± 0.17 | 0.73 ± 0.17 | 0.73 ± 0.19 | 0.953 | 0.898 | 0.871 |
| Total feed intake (kg) | |||||||
| d1–d15 | 9.95 ± 1.83 | 9.85 ± 2.44 | 9.00 ± 2.12 | 9.14 ± 2.73 | 0.316 | 0.978 | 0.879 |
| FCR (kg/kg) | |||||||
| d1–d8 | 2.44 ± 0.96 | 2.57 ± 1.29 | 1.89 ± 0.60 | 1.97 ± 0.63 | 0.096 | 0.765 | 0.932 |
| d8–d15 | 1.69 ± 0.53 | 1.97 ± 0.69 | 1.97 ± 0.63 | 1.93 ± 0.31 | 0.550 | 0.539 | 0.437 |
| d1–d15 ‡ | 1.76 ± 0.31 A | 2.00 ± 0.36 B | 1.72 ± 0.12 A | 1.93 ± 0.21 B | 0.593 | 0.031 | 0.902 |
| Condition | p Value | ||||||
|---|---|---|---|---|---|---|---|
| Parameter (Unit) | Basal | Basal + LM | MB40 | MB40 + LM | Diet | Challenge | D × C |
| Small intestine weight (g) † | 535 ± 111 a | 620 ± 81 a | 505 ± 70 b | 510 ± 83 b | 0.036 | 0.166 | 0.219 |
| Liver weight (g) | 401 ± 75 | 405 ± 107 | 410 ± 113 | 405 ± 68 | 0.894 | 0.999 | 0.901 |
| Kidney weight (g) | 36 ± 9 | 40 ± 10 | 35 ± 14 | 38 ± 6 | 0.635 | 0.305 | 0.851 |
| Spleen weight (g) | 27 ± 10 | 33 ± 10 | 26 ± 14 | 31 ± 10 | 0.664 | 0.207 | 0.934 |
| Normalized small intestine weight (% bw) | 6.3 ± 0.61 | 6.5 ± 0.98 | 5.9 ± 1.71 | 6.1 ± 0.97 | 0.294 | 0.687 | 0.896 |
| Normalized liver weight (% bw) | 3.3 ± 0.46 | 3.2 ± 0.53 | 3.3 ± 0.88 | 3.4 ± 0.76 | 0.651 | 0.976 | 0.726 |
| Normalized kidney weight (% bw) | 0.29 ± 0.02 | 0.33 ± 0.03 | 0.27 ± 0.08 | 0.32 ± 0.1 | 0.510 | 0.052 | 0.801 |
| Normalized spleen weight (% bw) ‡ | 0.22 ± 0.07 A | 0.26 ± 0.04 B | 0.20 ± 0.09 A | 0.27 ± 0.03 B | 0.795 | 0.017 | 0.585 |
| Condition | p Value | ||||||
|---|---|---|---|---|---|---|---|
| Parameter (Unit) | Basal | Basal + LM | MB40 | MB40 + LM | Diet | Challenge | D × C |
| Jejunum | |||||||
| Villus height (µM) | 490 ± 67 | 473 ± 51 | 437 ± 98 | 471 ± 66 | 0.298 | 0.746 | 0.334 |
| Crypt depth (µM) | 210 ± 50 | 227 ± 54 | 195 ± 17 | 202 ± 17 | 0.156 | 0.402 | 0.721 |
| VH:CD ratio | 2.46 ± 0.77 | 2.19 ± 0.59 | 2.25 ± 0.55 | 2.44 ± 0.26 | 0.919 | 0.845 | 0.271 |
| Ileum | |||||||
| Villus height (µM) | 454 ± 71 | 436 ± 38 | 432 ± 108 | 400 ± 114 | 0.360 | 0.417 | 0.822 |
| Crypt depth (µM) ‡ | 189 ± 25 A | 244 ± 48 B | 185 ± 30 A | 231 ± 57 B | 0.548 | 0.002 | 0.773 |
| VH:CD ratio ‡ | 2.43 ± 0.35 A | 1.86 ± 0.48 B | 2.35 ± 0.48 A | 1.85 ± 0.74 B | 0.818 | 0.009 | 0.870 |
| Large intestine | |||||||
| Crypt depth (µM) | 324 ± 59 | 324 ± 41 | 293 ± 74 | 298 ± 51 | 0.178 | 0.895 | 0.885 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garvey, S.M.; Emami, N.K.; Guice, J.L.; Sriranganathan, N.; Penet, C.; Rhoads, R.P.; Spears, J.L.; Dalloul, R.A.; El-Kadi, S.W. The Probiotic Bacillus subtilis MB40 Improves Immunity in a Porcine Model of Listeriosis. Microorganisms 2023, 11, 2110. https://doi.org/10.3390/microorganisms11082110
Garvey SM, Emami NK, Guice JL, Sriranganathan N, Penet C, Rhoads RP, Spears JL, Dalloul RA, El-Kadi SW. The Probiotic Bacillus subtilis MB40 Improves Immunity in a Porcine Model of Listeriosis. Microorganisms. 2023; 11(8):2110. https://doi.org/10.3390/microorganisms11082110
Chicago/Turabian StyleGarvey, Sean M., Nima K. Emami, Justin L. Guice, Nammalwar Sriranganathan, Christopher Penet, Robert P. Rhoads, Jessica L. Spears, Rami A. Dalloul, and Samer W. El-Kadi. 2023. "The Probiotic Bacillus subtilis MB40 Improves Immunity in a Porcine Model of Listeriosis" Microorganisms 11, no. 8: 2110. https://doi.org/10.3390/microorganisms11082110
APA StyleGarvey, S. M., Emami, N. K., Guice, J. L., Sriranganathan, N., Penet, C., Rhoads, R. P., Spears, J. L., Dalloul, R. A., & El-Kadi, S. W. (2023). The Probiotic Bacillus subtilis MB40 Improves Immunity in a Porcine Model of Listeriosis. Microorganisms, 11(8), 2110. https://doi.org/10.3390/microorganisms11082110