
Flavonoid Synthesis by Deinococcus sp. 43 Isolated from the Ginkgo Rhizosphere

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Genome Extraction
2.2. Genome Sequencing, Assembly and Functional Annotation for Flavonoid Synthesis
2.3. HPLC Sample Preparation and Detection Conditions
2.4. LC-MS/MS Sample Preparation and Detection Conditions
2.5. Metabolite Data Processing and Analysis
2.6. Gene Expression Analysis by RT-qPCR
3. Results
3.1. Strain Identification
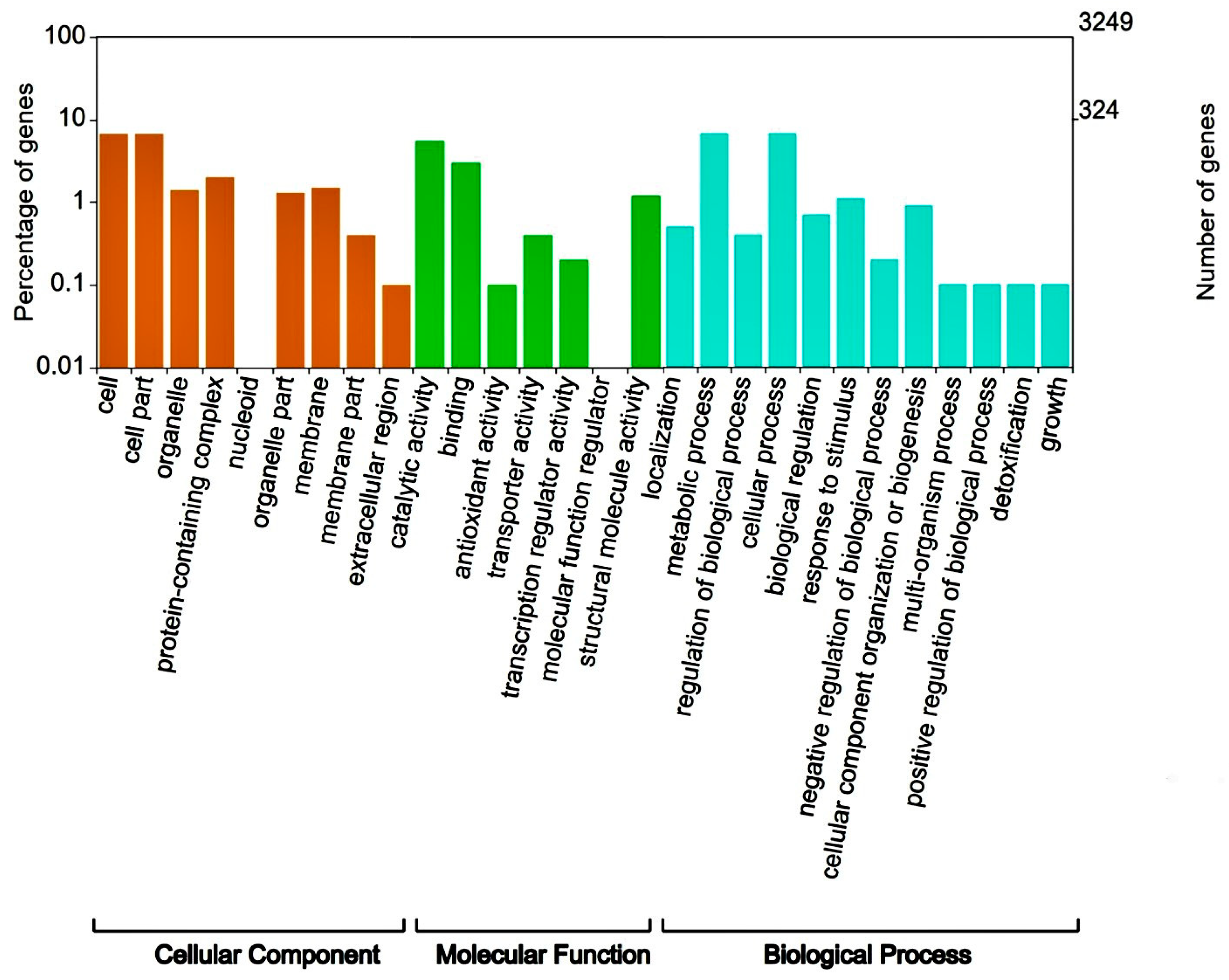
3.2. Genome Sequencing, Assembly and Functional Annotation for Flavonoid Synthesis
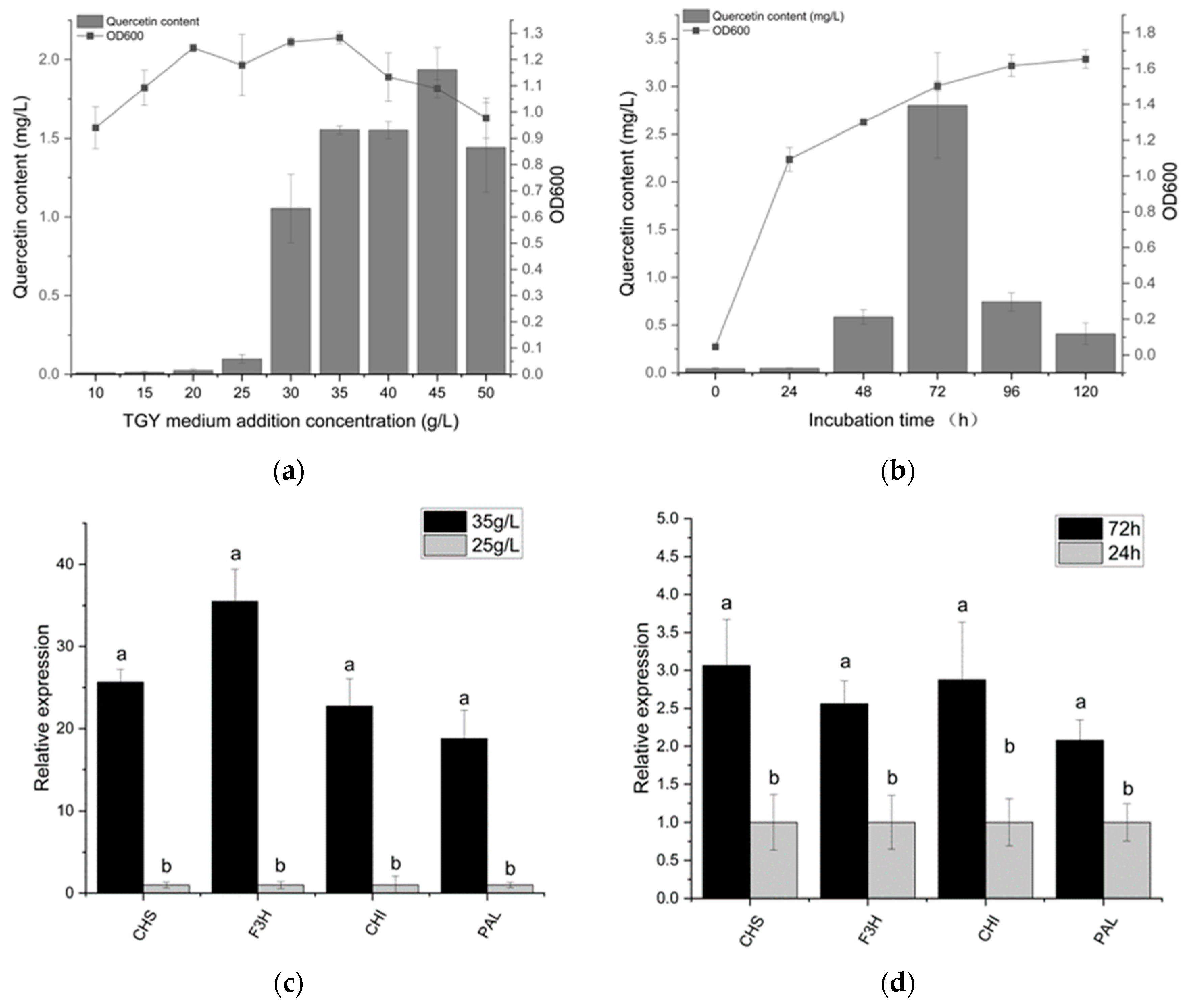
3.3. Quercetin Production Analysis and Optimization
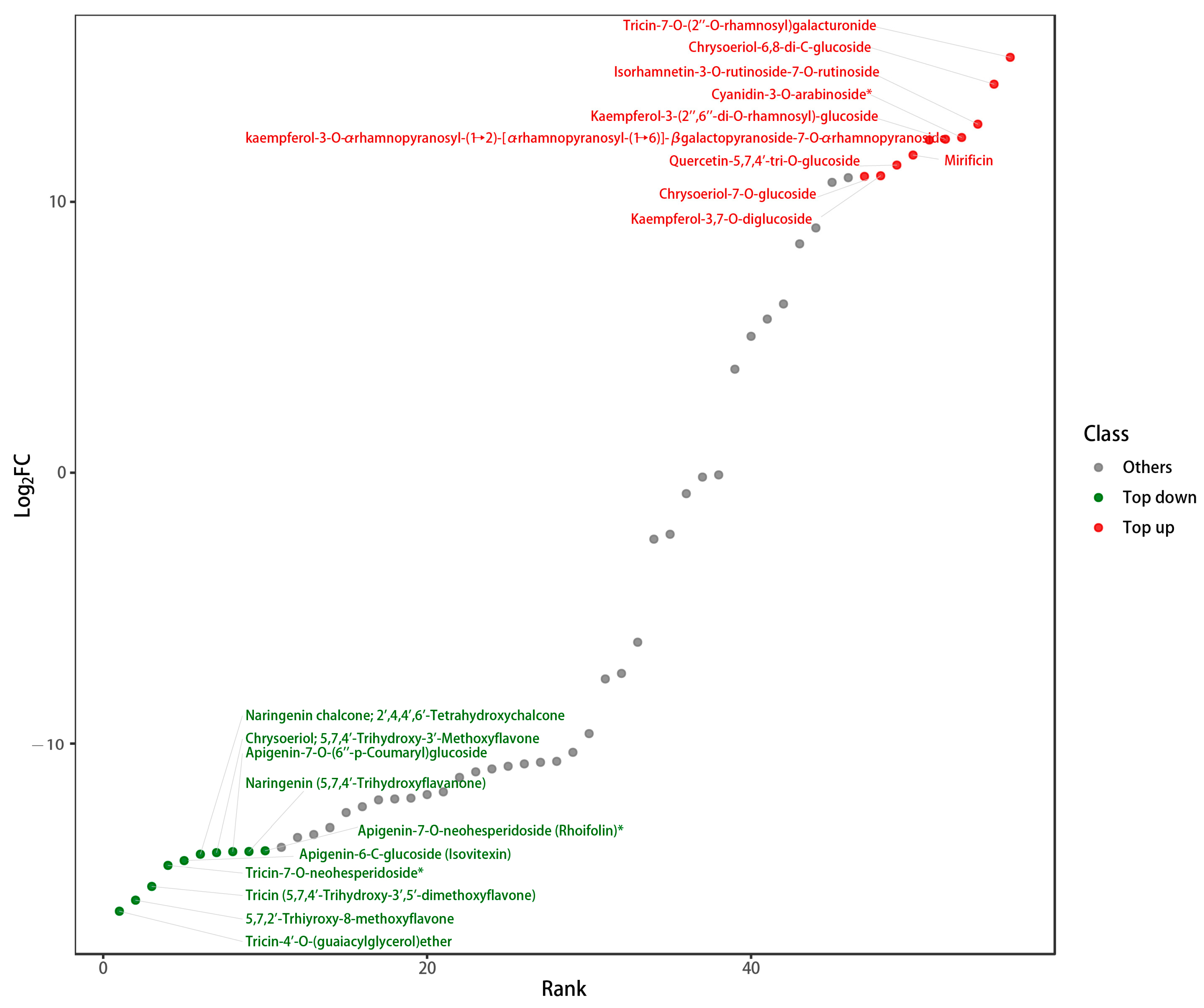
3.4. Analysis of Flavonoid Metabolites
3.5. Construction of the Genome-Scale Metabolic Model of Deinococcus sp. 43
4. Discussion
4.1. Deinococcus Becomes a Potential Source for Flavonoid Production
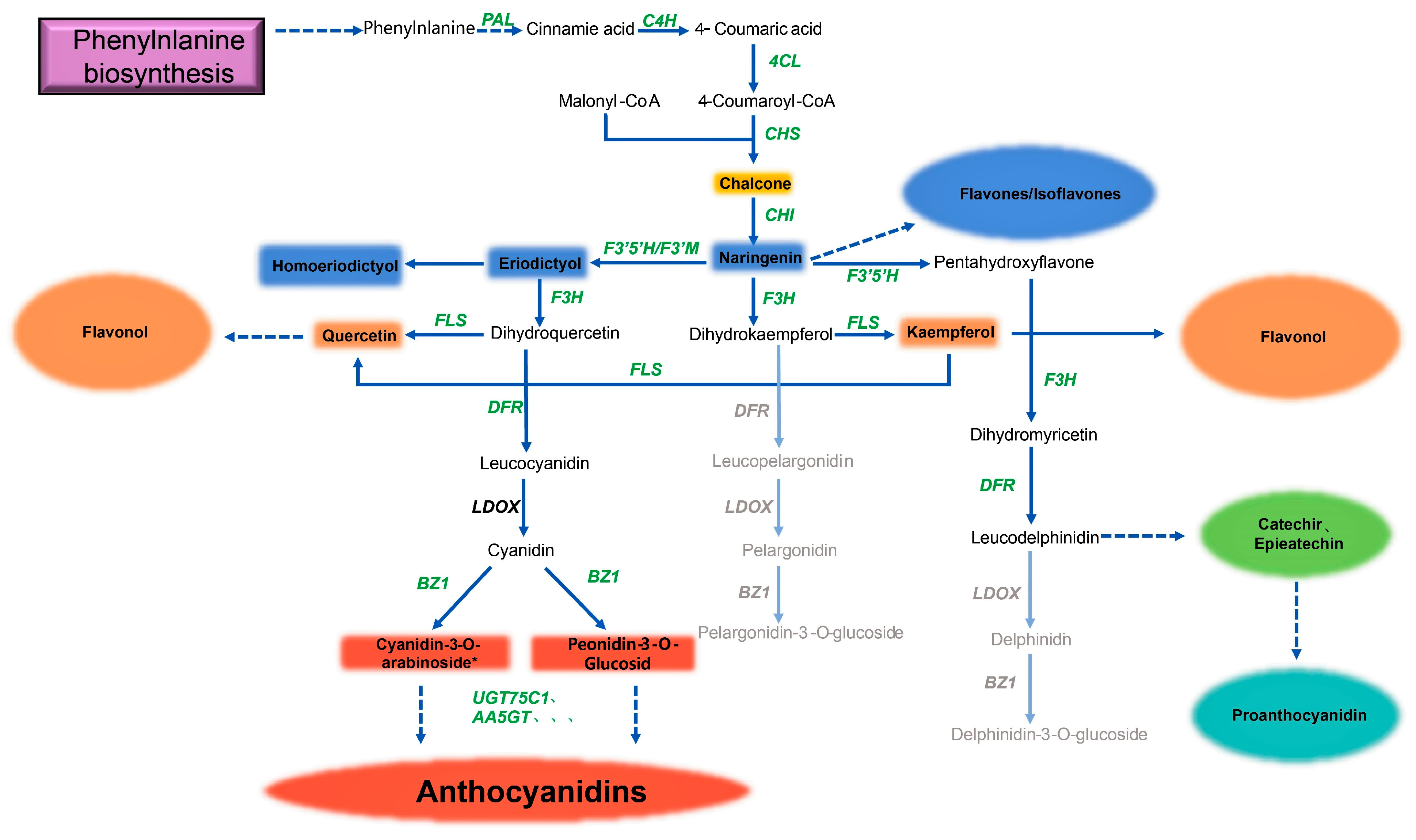
4.2. The Intricate Mechanism of Deinococcus’ Production of Flavonoids
4.3. The Variations in the Intracellular and Extracellular Distribution of Flavonoids Are Correlated with Their Distinct Biological Roles
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Forkmann, G.; Heller, W. Biosynthesis of flavonoids. In Comprehensive Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 1999; pp. 713–748. [Google Scholar]
- Wang, Y.C.; Chen, S.; Yu, O. Metabolic engineering of flavonoids in plants and microorganisms. Appl. Microbiol. Biot. 2011, 91, 949–956. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as Anticancer Agents. Nutrients 2020, 12, 457. [Google Scholar] [PubMed]
- Spagnuolo, C.; Moccia, S.; Russo, G.L. Anti-inflammatory effects of flavonoids in neurodegenerative disorders. Eur. J. Med. Chem. 2018, 153, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Chograni, H.; Riahi, L.; Zaouali, Y.; Boussaid, M. Polyphenols, flavonoids, antioxidant activity in leaves and flowers of Tunisian Globularia alypum L. (Globulariaceae). Afr. J. Ecol. 2013, 51, 343–347. [Google Scholar] [CrossRef]
- Hassan, Y.I.; Kosir, V.; Yin, X.; Ross, K.; Diarra, M.S. Grape pomace as a promising antimicrobial alternative in feed: A critical review. J. Agric. Food Chem. 2019, 67, 9705–9718. [Google Scholar] [CrossRef]
- Flesch, V.; Jacques, M.; Cosson, L.; Teng, B.; Petiard, V.; Balz, J. Relative importance of growth and light level on terpene content of Ginkgo biloba. Phytochemistry 1992, 31, 1941–1945. [Google Scholar] [CrossRef]
- Isah, T. Rethinking Ginkgo biloba L.: Medicinal uses and conservation. Pharmacogn. Rev. 2015, 9, 140. [Google Scholar] [CrossRef]
- Shen, N.; Wang, T.F.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef]
- Alseekh, S.; de Souza, L.P.; Benina, M.; Fernie, A.R. The style and substance of plant flavonoid decoration; towards defining both structure and function. Phytochemistry 2020, 174, 112347. [Google Scholar] [CrossRef]
- Brader, G.; Compant, S.; Mitter, B.; Trognitz, F.; Sessitsch, A. Metabolic potential of endophytic bacteria. Curr. Opin. Biotechnol. 2014, 27, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.H.; Bernard, S.M.; Andersen, G.L.; Chen, W. Developing microbe-plant interactions for applications in plant-growth promotion and disease control, production of useful compounds, remediation and carbon sequestration. Microb. Biotechnol. 2009, 2, 428–440. [Google Scholar] [CrossRef] [PubMed]
- Ludwig-Müller, J. Interplay between endophyte and host plant in the synthesis and modification of metabolites. In Endophyte Biotechnology: Potential for Agriculture and Pharmacology; CABI: Wallingford, UK, 2019; pp. 180–197. [Google Scholar]
- Ferreira, M.C.; de Carvalho, C.R.; Bahia, M.; Barreto, D.L.C.; Azevedo, R.N.; Cota, B.B.; Zani, C.L.; de Oliveira Santos, A.R.; Rosa, C.A.; Rosa, L.H. Plant-associated fungi: Methods for taxonomy, diversity, and bioactive secondary metabolite bioprospecting. Plant Microbiome: Methods Protoc. 2021, 2232, 85–112. [Google Scholar]
- Li, D.; Fu, D.W.; Zhang, Y.; Ma, X.L.; Gao, L.G.; Wang, X.H.; Zhou, D.P.; Zhao, K. Isolation, Purification, and Identification of Taxol and Related Taxanes from Taxol-Producing Fungus Aspergillus niger subsp taxi. J. Microbiol. Biotechnol. 2017, 27, 1379–1385. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Zeng, Q.C.; Shao, Y.B.; Zhou, H.X.; Li, T.J.; Song, F.; Liu, W. Anti-cervical cancer activity of secondary metabolites of endophytic fungi from Ginkgo biloba. Cancer Biomark. 2020, 28, 371–379. [Google Scholar] [CrossRef]
- Hacquard, S.; Garrido-Oter, R.; Gonzalez, A.; Spaepen, S.; Ackermann, G.; Lebeis, S.; McHardy, A.C.; Dangl, J.L.; Knight, R.; Ley, R.; et al. Microbiota and Host Nutrition across Plant and Animal Kingdoms. Cell Host Microbe 2015, 17, 603–616. [Google Scholar] [CrossRef]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttila, A.M.; Compant, S.; Campisano, A.; Doring, M.; Sessitsch, A. The Hidden World within Plants: Ecological and Evolutionary Considerations for Defining Functioning of Microbial Endophytes. Microbiol. Mol. Biol. R. 2015, 79, 293–320. [Google Scholar] [CrossRef]
- Provorov, N.A.; Vorobyov, N.I. Host plant as an organizer of microbial evolution in the beneficial symbioses. Phytochem. Rev. 2009, 8, 519–534. [Google Scholar] [CrossRef]
- Wang, H.; Guo, Y.J.; Luo, Z.; Gao, L.W.; Li, R.; Zhang, Y.X.; Kalaji, H.M.; Qiang, S.; Chen, S.G. Recent Advances in Alternaria Phytotoxins: A Review of Their Occurrence, Structure, Bioactivity, and Biosynthesis. J. Fungi 2022, 8, 168. [Google Scholar] [CrossRef]
- Qiu, M.; Xie, R.S.; Shi, Y.; Zhang, H.H.; Chen, H.M. Isolation and identification of two flavonoid-producing endophytic fungi from Ginkgo biloba L. Ann. Microbiol. 2010, 60, 143–150. [Google Scholar] [CrossRef]
- Ma, J.; Zhang, X.L.; Wang, Y.; Zheng, J.Y.; Wang, C.Y.; Shao, C.L. Aspergivones A and B, two new flavones isolated from a gorgonian-derived Aspergillus candidus fungus. Nat. Prod. Res. 2017, 31, 32–36. [Google Scholar] [CrossRef]
- Xu, D.B.; Ye, W.W.; Han, Y.; Deng, Z.X.; Hong, K. Natural Products from Mangrove Actinomycetes. Mar. Drugs 2014, 12, 2590–2613. [Google Scholar] [CrossRef]
- Cao, D.D.; Do, T.Q.; Mai, H.D.T.; Thi, Q.V.; Nguyen, M.A.; Thi, H.M.L.; Tran, D.T.; Chau, V.M.; Thung, D.C.; Pham, V.C. Antimicrobial lavandulylated flavonoids from a sponge-derived actinomycete. Nat. Prod. Res. 2020, 34, 413–420. [Google Scholar] [CrossRef]
- Rha, C.-S.; Kim, H.G.; Baek, N.-I.; Kim, D.-O.; Park, C.-S. Amylosucrase from Deinococcus geothermalis can be modulated under different reaction conditions to produce novel quercetin 4′-O-α-D-isomaltoside. Enzym. Microb. Technol. 2020, 141, 109648. [Google Scholar] [CrossRef]
- Deng, Y.; Huang, H.N.; Lei, F.Y.; Fu, S.D.; Zou, K.; Zhang, S.F.; Liu, X.D.; Jiang, L.H.; Liu, H.W.; Miao, B.; et al. Endophytic Bacterial Communities of Ginkgo biloba Leaves During Leaf Developmental Period. Front. Microbiol. 2021, 12, 698703. [Google Scholar] [CrossRef] [PubMed]
- Prjibelski, A.; Antipov, D.; Meleshko, D.; Lapidus, A.; Korobeynikov, A. Using SPAdes de novo assembler. Curr. Protoc. Bioinform. 2020, 70, e102. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Moriya, Y.; Itoh, M.; Okuda, S.; Yoshizawa, A.C.; Kanehisa, M. KAAS: An automatic genome annotation and pathway reconstruction server. Nucleic Acids Res. 2007, 35, W182–W185. [Google Scholar] [CrossRef]
- Huerta-Cepas, J.; Forslund, K.; Coelho, L.P.; Szklarczyk, D.; Jensen, L.J.; von Mering, C.; Bork, P. Fast Genome-Wide Functional Annotation through Orthology Assignment by eggNOG-Mapper. Mol. Biol. Evol. 2017, 34, 2115–2122. [Google Scholar] [CrossRef]
- Huerta-Cepas, J.; Szklarczyk, D.; Heller, D.; Hernandez-Plaza, A.; Forslund, S.K.; Cook, H.; Mende, D.R.; Letunic, I.; Rattei, T.; Jensen, L.J.; et al. eggNOG 5.0: A hierarchical, functionally and phylogenetically annotated orthology resource based on 5090 organisms and 2502 viruses. Nucleic Acids Res. 2019, 47, D309–D314. [Google Scholar] [CrossRef] [PubMed]
- Darzi, Y.; Letunic, I.; Bork, P.; Yamada, T. iPath3.0: Interactive pathways explorer v3. Nucleic Acids Res. 2018, 46, W510–W513. [Google Scholar] [CrossRef] [PubMed]
- Keshri, P.K.; Rai, N.; Verma, A.; Kamble, S.C.; Barik, S.; Mishra, P.; Singh, S.K.; Salvi, P.; Gautam, V. Biological potential of bioactive metabolites derived from fungal endophytes associated with medicinal plants. Mycol. Prog. 2021, 20, 577–594. [Google Scholar] [CrossRef]
- Zou, K.; Liu, X.D.; Hu, Q.; Zhang, D.; Fu, S.D.; Zhang, S.F.; Huang, H.N.; Lei, F.Y.; Zhang, G.Q.; Miao, B.; et al. Root Endophytes and Ginkgo biloba Are Likely to Share and Compensate Secondary Metabolic Processes, and Potentially Exchange Genetic Information by LTR-RTs. Front. Plant Sci. 2021, 12, 704985. [Google Scholar] [CrossRef]
- Jian, Z.Y.; Meng, L.; Hu, X.Q. An endophytic fungus efficiently producing paclitaxel isolated from Taxus wallichiana var. mairei. Medicine 2017, 96, e7406. [Google Scholar]
- Xiong, Z.Q.; Yang, Y.Y.; Zhao, N.; Wang, Y. Diversity of endophytic fungi and screening of fungal paclitaxel producer from Anglojap yew, Taxus x media. BMC Microbiol. 2013, 13, 71. [Google Scholar] [CrossRef]
- Daly, M.J.; Gaidamakova, E.K.; Matrosova, V.Y.; Vasilenko, A.; Zhai, M.; Leapman, R.D.; Lai, B.; Ravel, B.; Li, S.M.W.; Kemner, K.M.; et al. Protein oxidation implicated as the primary determinant of bacterial radioresistance. PLoS Biol. 2007, 5, 769–779. [Google Scholar] [CrossRef]
- Tian, B.; Hua, Y.J. Carotenoid biosynthesis in extremophilic Deinococcus-Thermus bacteria. Trends Microbiol. 2010, 18, 512–520. [Google Scholar] [CrossRef]
- Krinsky, N.I.; Johnson, E.J. Carotenoid actions and their relation to health and disease. Mol. Asp. Med. 2005, 26, 459–516. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.L.; Pei, Y.C.; Huang, W.; Ding, J.Q.; Siemann, E. Increasing flavonoid concentrations in root exudates enhance associations between arbuscular mycorrhizal fungi and an invasive plant. ISME J. 2021, 15, 1919–1930. [Google Scholar] [CrossRef] [PubMed]
- Barrera, A.; Hereme, R.; Ruiz-Lara, S.; Larrondo, L.F.; Gundel, P.E.; Pollmann, S.; Molina-Montenegro, M.A.; Ramos, P. Fungal Endophytes Enhance the Photoprotective Mechanisms and Photochemical Efficiency in the Antarctic Colobanthus quitensis (Kunth) Bartl. Exposed to UV-B Radiation. Front. Ecol. Evol. 2020, 8, 122. [Google Scholar] [CrossRef]
- Jia, M.; Chen, L.; Xin, H.L.; Zheng, C.J.; Rahman, K.; Han, T.; Qin, L.P. A Friendly Relationship between Endophytic Fungi and Medicinal Plants: A Systematic Review. Front. Microbiol. 2016, 7, 906. [Google Scholar] [CrossRef]
- Zhao, J.; Shan, T.; Mou, Y.; Zhou, L. Plant-Derived Bioactive Compounds Produced by Endophytic Fungi. Mini-Rev. Med. Chem. 2011, 11, 159–168. [Google Scholar] [CrossRef]
- Wang, X. Regulation and Switching in Bacterial Gene Expression Networks in Response to Nutrients; University of Illinois at Urbana-Champaign: Champaign, IL, USA, 2020. [Google Scholar]
- Schvartzman, J.M.; Thompson, C.B.; Finley, L.W.S. Metabolic regulation of chromatin modifications and gene expression. J. Cell Biol. 2018, 217, 2247–2259. [Google Scholar] [CrossRef]
- Lillo, C.; Lea, U.S.; Ruoff, P. Nutrient depletion as a key factor for manipulating gene expression and product formation in different branches of the flavonoid pathway. Plant Cell Environ. 2008, 31, 587–601. [Google Scholar] [CrossRef] [PubMed]
- Naghavi, F.S.; Hanachi, P.; Soudi, M.R.; Saboora, A.; Ghorbani, A. Evaluation of the Relationship between the Incubation Time and Carotenoid Production in Rhodotorula slooffiae and R. mucilaginosa Isolated from Leather Tanning Wastewater. Iran. J. Basic. Med. Sci. 2013, 16, 1114–1118. [Google Scholar]
- Steed, A.L.; Christophi, G.P.; Kaiko, G.E.; Sun, L.L.; Goodwin, V.M.; Jain, U.; Esaulova, E.; Artyomov, M.N.; Morales, D.J.; Holtzman, M.J.; et al. The microbial metabolite desaminotyrosine protects from influenza through type I interferon. Science 2017, 357, 498–502. [Google Scholar] [CrossRef]
- Westlake, D.W.; Roxburgh, J.M.; Talbot, G. Microbial production of carbon monoxide from flavonoids. Nature 1961, 189, 510–511. [Google Scholar] [CrossRef]
- Westlake, D.W.; Spencer, J.F. The utilization of flavonoid compounds by yeasts and yeast-like fungi. Can. J. Microbiol. 1966, 12, 165–174. [Google Scholar] [CrossRef]
- Schoefer, L.; Mohan, R.; Schwiertz, A.; Braune, A.; Blaut, M. Anaerobic degradation of flavonoids by Clostridium orbiscindens. Appl. Environ. Microbiol. 2003, 69, 5849–5854. [Google Scholar] [CrossRef] [PubMed]
- Koes, R.E.; Quattrocchio, F.; Mol, J.N. The flavonoid biosynthetic pathway in plants: Function and evolution. BioEssays 1994, 16, 123–132. [Google Scholar] [CrossRef]
- Dixon, R.A.; Paiva, N.L. Stress-Induced Phenylpropanoid Metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef]
- Dooner, H.K.; Robbins, T.P.; Jorgensen, R.A. Genetic and developmental control of anthocyanin biosynthesis. Annu. Rev. Genet. 1991, 25, 173–199. [Google Scholar] [CrossRef]
- Shaw, L.J.; Morris, P.; Hooker, J.E. Perception and modification of plant flavonoid signals by rhizosphere microorganisms. Environ. Microbiol. 2006, 8, 1867–1880. [Google Scholar] [CrossRef] [PubMed]
- Heller, W.; Forkmann, G. Biosynthesis of flavonoids. In The Flavonoids; Routledge: London, UK, 2017; pp. 499–535. [Google Scholar]
- Chen, L.; Chang, S.; Zhao, L.; Li, B.; Zhang, S.; Yun, C.; Wu, X.; Meng, J.; Li, G.; Guo, S.; et al. Biosynthesis of a water solubility-enhanced succinyl glucoside derivative of luteolin and its neuroprotective effect. Microb. Biotechnol. 2022, 15, 2401–2410. [Google Scholar] [CrossRef]
- Husain, S.R.; Cillard, J.; Cillard, P. Hydroxyl radical scavenging activity of flavonoids. Phytochemistry 1987, 26, 2489–2491. [Google Scholar] [CrossRef]
- Burda, S.; Oleszek, W. Antioxidant and antiradical activities of flavonoids. J. Agric. Food Chem. 2001, 49, 2774–2779. [Google Scholar] [CrossRef]
- Banjarnahor, S.D.; Artanti, N. Antioxidant properties of flavonoids. Med. J. Indones. 2014, 23, 239–244. [Google Scholar] [CrossRef]
- Kejik, Z.; Kaplanek, R.; Masarik, M.; Babula, P.; Matkowski, A.; Filipensky, P.; Vesela, K.; Gburek, J.; Sykora, D.; Martasek, P.; et al. Iron Complexes of Flavonoids-Antioxidant Capacity and Beyond. Int. J. Mol. Sci. 2021, 22, 646. [Google Scholar] [CrossRef]
- Zhao, J.Y.; Ren, F.L. Influence of hydroxylation and glycosylation in ring A of soybean isoflavones on interaction with BSA. Spectrochim. Acta A 2009, 72, 682–685. [Google Scholar] [CrossRef] [PubMed]
- Rha, C.S.; Jung, Y.S.; Seo, D.H.; Kim, D.O.; Park, C.S. Site-specific alpha-glycosylation of hydroxyflavones and hydroxyflavanones by amylosucrase from Deinococcus geothermalis. Enzyme Microb. Technol. 2019, 129, 109361. [Google Scholar] [CrossRef] [PubMed]
- He, Y. High cell density production of Deinococcus radiodurans under optimized conditions. J. Ind. Microbiol. Biotechnol. 2009, 36, 539–546. [Google Scholar] [CrossRef]
- Hostetler, G.; Riedl, K.; Cardenas, H.; Diosa-Toro, M.; Arango, D.; Schwartz, S.; Doseff, A.I. Flavone deglycosylation increases their anti-inflammatory activity and absorption. Mol. Nutr. Food Res. 2012, 56, 558–569. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.Y.; Sun, C.X.; Mao, L.K.; Ma, P.H.; Liu, F.G.; Yang, J.; Gao, Y.X. The biological activities, chemical stability, metabolism and delivery systems of quercetin: A review. Trends Food Sci. Tech. 2016, 56, 21–38. [Google Scholar] [CrossRef]
- Ren, S.C.; Qiao, Q.Q.; Ding, X.L. Antioxidative Activity of Five Flavones Glycosides from Corn Silk (Stigma maydis). Czech J. Food Sci. 2013, 31, 148–155. [Google Scholar] [CrossRef]
- Skogman, M.E.; Kanerva, S.; Manner, S.; Vuorela, P.M.; Fallarero, A. Flavones as Quorum Sensing Inhibitors Identified by a Newly Optimized Screening Platform Using Chromobacterium violaceum as Reporter Bacteria. Molecules 2016, 21, 1211. [Google Scholar] [CrossRef]
- Xie, Y.X.; Chen, J.X.; Wang, B.; Peng, A.Y.; Mao, Z.W.; Xia, W. Inhibition of Quorum-Sensing Regulator from Pseudomonas aeruginosa Using a Flavone Derivative. Molecules 2022, 27, 2439. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene | Sequence of Primers (5′–3′) |
|---|---|---|
| PAL-F | PAL | AACCCTCTGATCTTCCCCGA |
| PAL-R | ACTTTCAGCGTGTCGATGGT | |
| CHS-F | CHS | ACATGAGTAGCGTCACCGTC |
| CHS-R | AGTTCAGGAGGACGTGTTCG | |
| F3H-F | F3H | GGAACGGGATCGAGGTCTAC |
| F3H-R | CACGGCTTTCGTGTCCATCT | |
| CHI-F | CHI | GGCGTCGGTCGCCTATTT |
| CHI-R | TCAGCCACGCGTGATTCAG | |
| RLPL32-F | RLPL | GTCCCCAAGAAGAAGACCAGC |
| RLPL32-R | CCTGGCGGCCATCGTAGTAA |
| Attributes | Characteristic |
|---|---|
| Clean reads | 4,017,618 |
| Clean base | 1,205,285,400 |
| Q20 (%) | 96.28% |
| N50 | 203,832 bp |
| Total base | 4,314,008 bp |
| GC content (%) | 69.53% |
| CDS | 4116 |
| tRNA genes | 56 |
| rRNA genes | 2 |
| Genes assigned to NR | 4036 (96.67%) |
| Genes assigned to COG | 3252 (77.89%) |
| Genes assigned to KEGG | 1808 (43.31%) |
| Compound | DI | DE | p Value |
|---|---|---|---|
| Hesperetin | 1.28 × 104 | 6.99 × 104 | 0.001178 |
| Quercetin | 2.62 × 104 | 2.77 × 104 | 0.238109 |
| 7-O-Methylnaringenin | 0 | 1.76 × 104 | 0.004591 |
| 3,4′-Dihydroxyflavone | 3.16 × 105 | 1.52 × 106 | 0.001678 |
| Kaempferol-3-O-(6″-Acetyl) glucosyl -(1→3)-Galactoside | 2.22 × 105 | 6.75 × 103 | 6.36 × 10−5 |
| 7-O-Methyleriodictyol | 0 | 4.63 × 104 | 0.033842 |
| Tricin (5,7,4′-Trihydroxy-3′,5′-dimethoxyflavone) | 0 | 3.52 × 105 | 0.012179 |
| 5,7,2′-Trhiyroxy-8-methoxy flavone | 0 | 5.00 × 105 | 0.008064 |
| Naringenin (5,7,4′-Trihydroxyflavanone) | 0 | 1.44 × 105 | 0.03777 |
| Tricin-4′-O-(guaiacylglycerol) ether | 0 | 6.62 × 105 | 0.008651 |
| Kaempferol-3-O-rutinoside-7-O-rhamnoside | 3.15 × 103 | 0 | 0.000327 |
| Gnetifolin B | 0 | 9.34 × 104 | 0.009861 |
| Chrysoeriol; 5,7,4′-Trihydroxy-3′-Methoxyflavone | 0 | 1.48 × 105 | 0.004042 |
| Naringenin chalcone; 2′,4,4′,6′-Tetrahydroxychalcone | 0 | 1.55 × 105 | 0.039474 |
| Chrysoeriol-7-O-homovanillic acid | 0 | 1.55 × 104 | 0.004311 |
| Tricin-4′-O-syringic acid | 0 | 2.18 × 104 | 0.110615 |
| Tricin-4′-O-oxalic acid-7-O-(p-coumaroyl) shikimic acid | 0 | 3.39 × 104 | 0.141641 |
| Kaempferol-3,7-O-diglucoside | 1.80 × 104 | 0 | 0.035479 |
| Tricin-4′-O-eudesmic acid | 0 | 3.17 × 104 | 0.040095 |
| Apigenin; 4′,5,7-Trihydroxyflavone | 0 | 1.90 × 104 | 0.015337 |
| Psoralenol | 4.76 × 104 | 8.14 × 104 | 0.003094 |
| Tricin-7-O-(2″-O-rhamnosyl) galacturonide | 3.72 × 105 | 0 | 0.147061 |
| Kaempferol-3-(2″,6″-di-O-rhamnosyl)-glucoside | 4.58 × 104 | 0 | 0.000393 |
| Cyanidin-3-O-arabinoside * | 4.78 × 104 | 0 | 0.005653 |
| Eriodictyol (5,7,3′,4′-Tetrahydroxyflavanone) | 0 | 3.89 × 104 | 0.016676 |
| Apigenin-6-C-glucoside (Isovitexin) | 0 | 1.82 × 105 | 0.017304 |
| Quercetagetin; 3,3′,4′,5,6,7-Hexahydroxyflavone | 0 | 5.37 × 104 | 0.003139 |
| Tricin-4′-O-glycerol | 0 | 3.79 × 104 | 0.011644 |
| Tricin-7-O-(2″-Malonyl) rhamnoside | 0 | 3.72 × 104 | 0.146934 |
| Isorhamnetin-3-O-rutinoside-7-O-rutinoside | 6.72 × 104 | 0 | 0.000119 |
| Apigenin-8-C-Glucoside (Vitexin) | 0 | 1.01 × 105 | 0.026334 |
| Tricin-4′-O-syringyl alcohol | 0 | 7.86 × 104 | 0.003272 |
| Kaempferol (3,5,7,4′-Tetrahydroxyflavone) | 0 | 7.13 × 103 | 0.051729 |
| Malvidin-3,5-di-O-glucoside | 4.73 × 103 | 0 | 0.005893 |
| Chrysoeriol-7-O-(6″-acetyl) glucoside | 0 | 1.76 × 103 | 0.017605 |
| Tricin-7-O-rutinoside * | 3.96 × 103 | 3.02 × 105 | 0.039741 |
| Gallocatechin-(4α→8)-gallocatechin | 9.73 × 104 | 1.30 × 103 | 0.037396 |
| Apigenin-7-O-neohesperidoside (Rhoifolin) * | 0 | 1.42 × 105 | 0.007817 |
| Kaempferol-3-O-(2-O-Xylosyl-6-O-Rhamnosyl) Glucoside | 2.00 × 104 | 1.41 × 103 | 0.026179 |
| Chrysoeriol-7-O-glucoside | 1.77 × 104 | 0 | 0.020361 |
| Mirificin | 3.05 × 104 | 0 | 0.004285 |
| Chrysoeriol-5,7-di-O-rutinoside | 1.52 × 104 | 0 | 0.014933 |
| Homoeriodictyol | 0 | 1.15 × 104 | 0.004614 |
| Chrysoeriol-6,8-di-C-glucoside | 1.87 × 105 | 0 | 0.002926 |
| Apigenin-7-O-(6″-p-Coumaryl) glucoside | 0 | 1.45 × 105 | 0.019662 |
| Nepetin (5,7,3′,4′-Tetrahydroxy-6-methoxyflavone) | 0 | 1.64 × 104 | 0.002265 |
| Procyanidin B1 | 0 | 1.53 × 103 | 0.000211 |
| Quercetin-5,7,4′-tri-O-glucoside | 2.36 × 104 | 0 | 0.002187 |
| Tricin-7-O-neohesperidoside * | 0 | 2.06 × 105 | 1.21 × 10−6 |
| Luteolin-7-O-gentiobioside | 7.24 × 104 | 1.42 × 103 | 0.03487 |
| Apiin | 3.06 × 104 | 3.42 × 104 | 0.285151 |
| Eriodictyol-3′-O-glucoside | 0 | 1.49 × 104 | 0.027668 |
| Apigenin-7-O-rutinoside (Isorhoifolin) * | 0 | 1.30 × 105 | 1.61 × 10−5 |
| Dihydrokaempferol-7-O-glucoside | 0 | 1.45 × 104 | 0.012045 |
| Catechin gallate | 1.71 × 104 | 0 | 0.011896 |
| kaempferol-3-O-α-rhamnopyranosyl-(1→2)- [α-rhamnopyranosyl-(1→6)]-β-galactopyranoside-7-O-α-rhamnopyranoside | 4.48 × 104 | 0 | 0.002381 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, J.; Zou, K.; Fu, S.; Duan, Z.; Zhang, G.; Wu, X.; Huang, J.; Li, S.; Liu, X.; Zhang, S.; et al. Flavonoid Synthesis by Deinococcus sp. 43 Isolated from the Ginkgo Rhizosphere. Microorganisms 2023, 11, 1848. https://doi.org/10.3390/microorganisms11071848
Zhou J, Zou K, Fu S, Duan Z, Zhang G, Wu X, Huang J, Li S, Liu X, Zhang S, et al. Flavonoid Synthesis by Deinococcus sp. 43 Isolated from the Ginkgo Rhizosphere. Microorganisms. 2023; 11(7):1848. https://doi.org/10.3390/microorganisms11071848
Chicago/Turabian StyleZhou, Jin, Kai Zou, Shaodong Fu, Zhenchun Duan, Guoqing Zhang, Xinhong Wu, Jingwen Huang, Shihui Li, Xueduan Liu, Shuangfei Zhang, and et al. 2023. "Flavonoid Synthesis by Deinococcus sp. 43 Isolated from the Ginkgo Rhizosphere" Microorganisms 11, no. 7: 1848. https://doi.org/10.3390/microorganisms11071848
APA StyleZhou, J., Zou, K., Fu, S., Duan, Z., Zhang, G., Wu, X., Huang, J., Li, S., Liu, X., Zhang, S., & Liang, Y. (2023). Flavonoid Synthesis by Deinococcus sp. 43 Isolated from the Ginkgo Rhizosphere. Microorganisms, 11(7), 1848. https://doi.org/10.3390/microorganisms11071848