
Codon Usage Bias Analysis in Macronuclear Genomes of Ciliated Protozoa


Abstract
1. Introduction
2. Materials and Methods
2.1. Data Sources
2.2. Nucleotide Composition Analysis
2.3. Effective Number of Codons
2.4. Codon Adaptation Index (CAI)
2.5. Relative Synonymous Codon Usage (RSCU) and Putative Optimal Codons
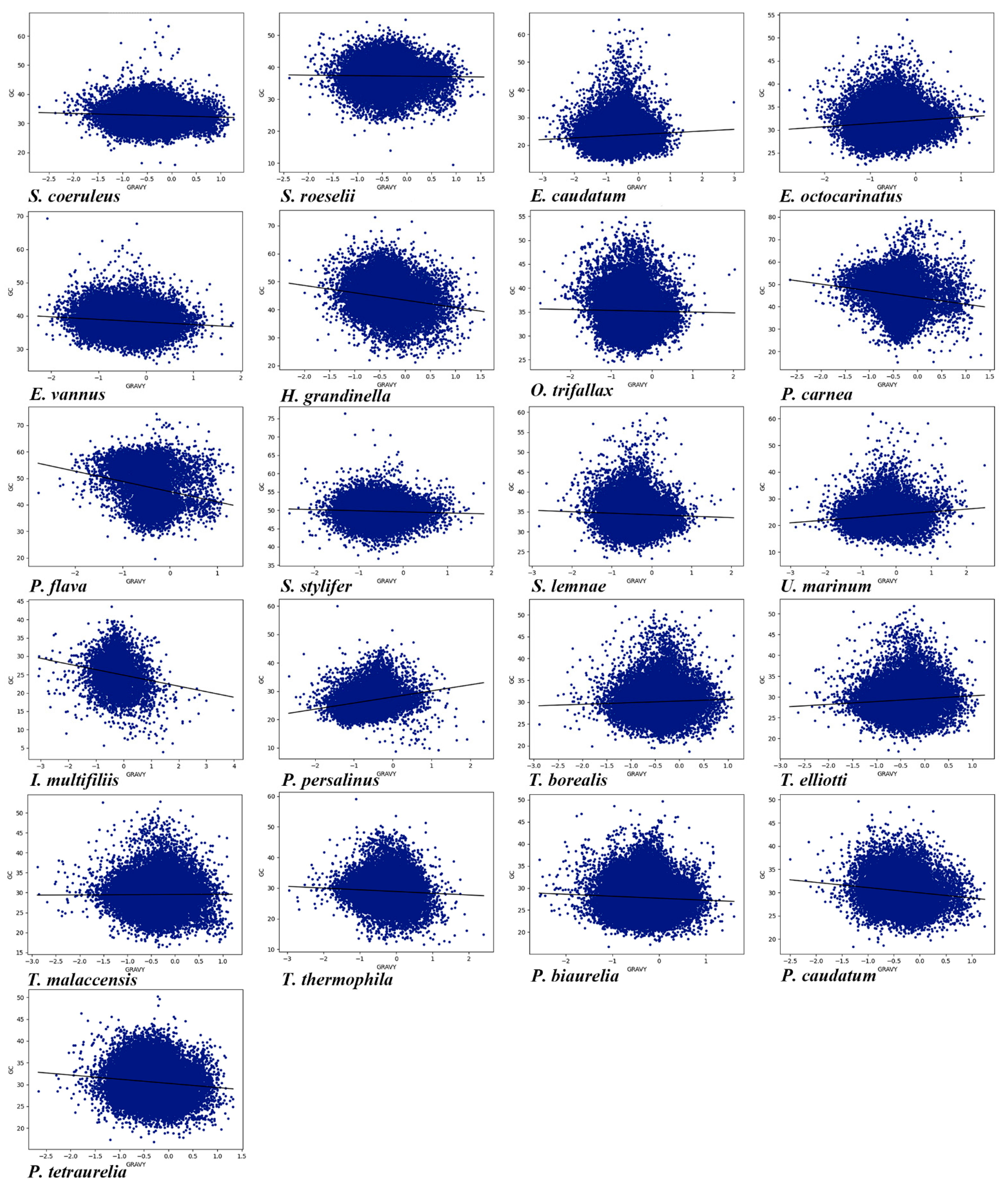
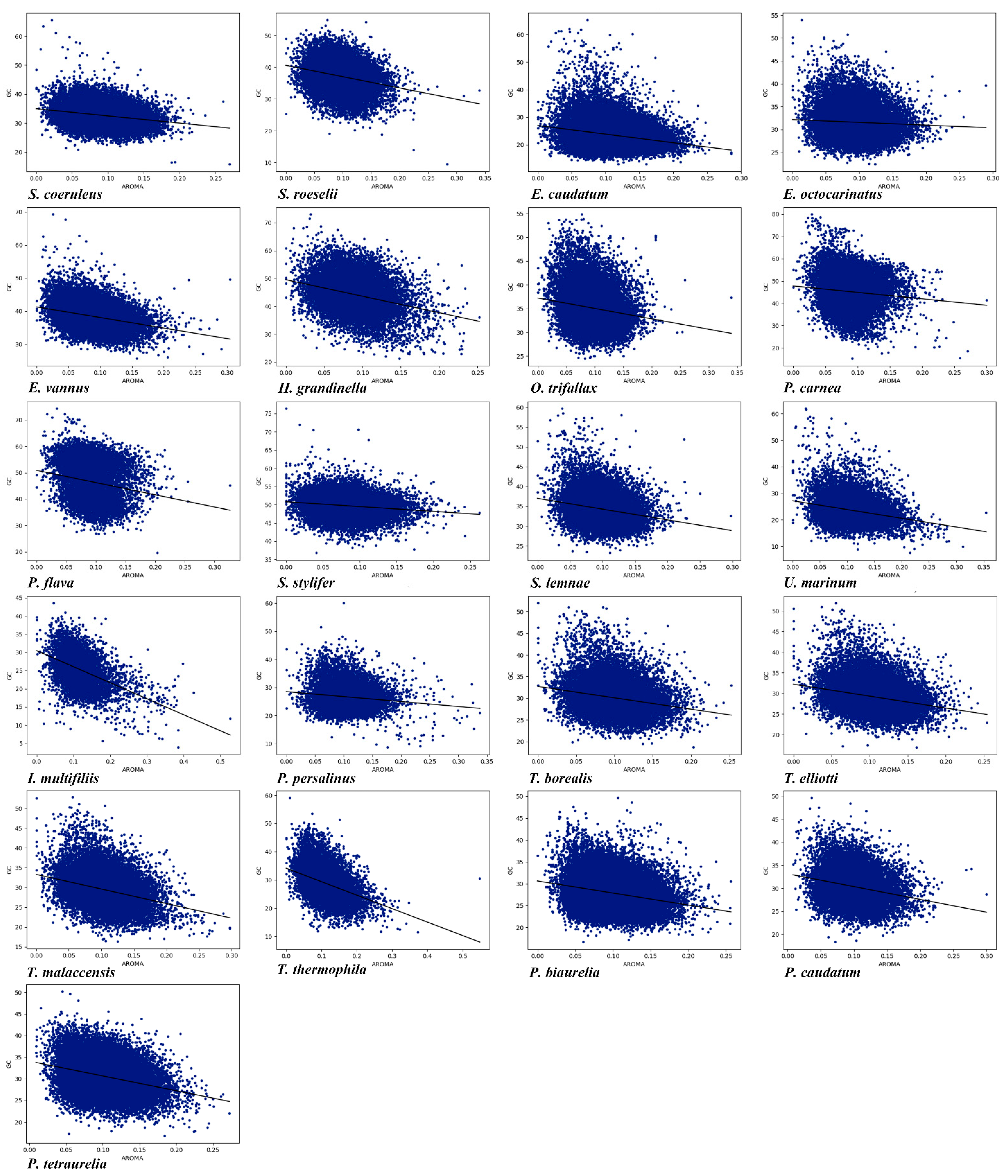
2.6. Grand Average Hydropathicity (Gravy) and Aromaticity (Aroma) Indices
2.7. Correspondence Analysis
2.8. Parity Rule 2 (PR2) Plot Analysis
2.9. Translational Selection Index
2.10. Neutrality Plot Analysis
2.11. Statistical Analysis
2.12. Phylogenetic Analysis
3. Results
3.1. Nucleotide Compositions
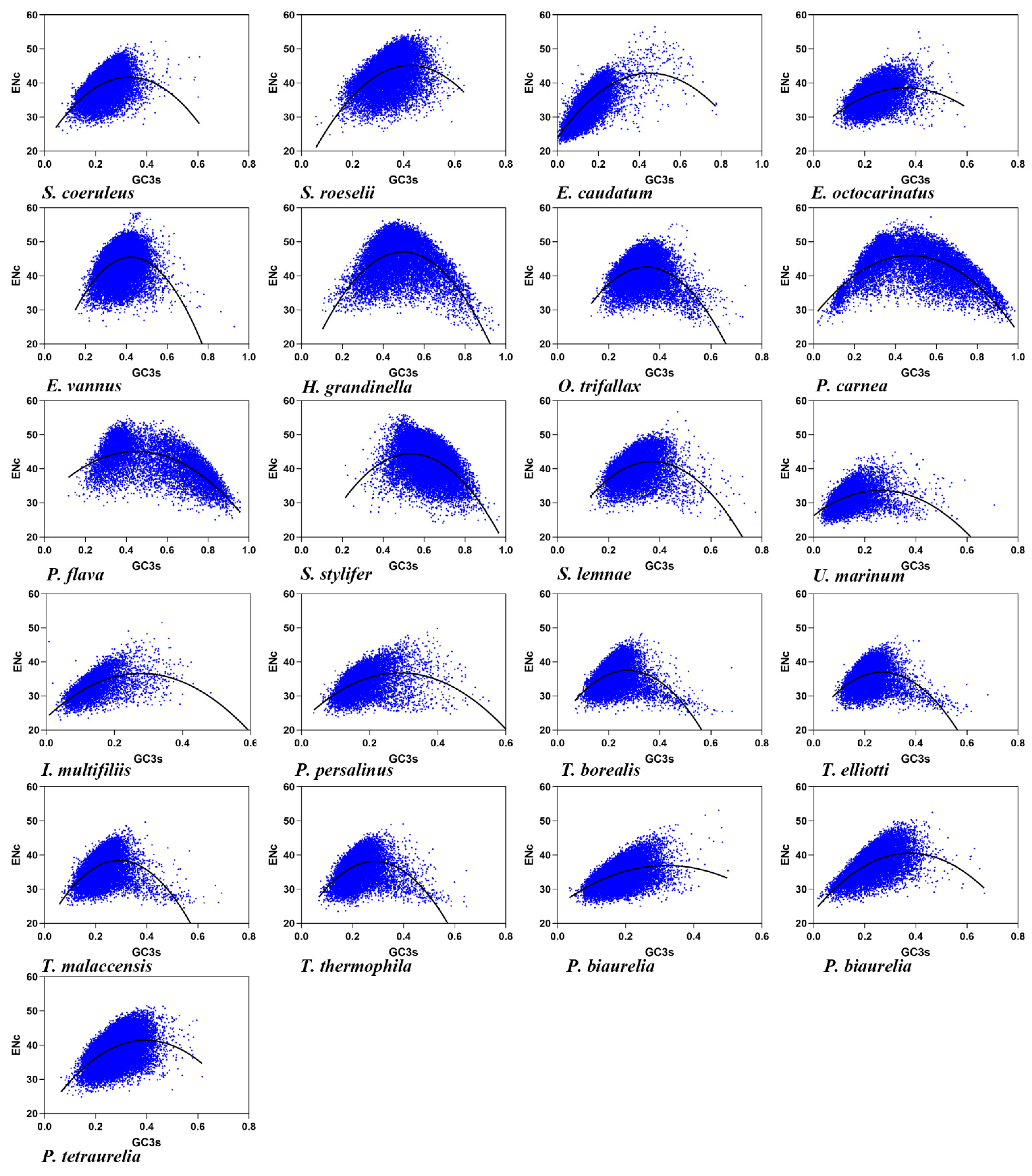
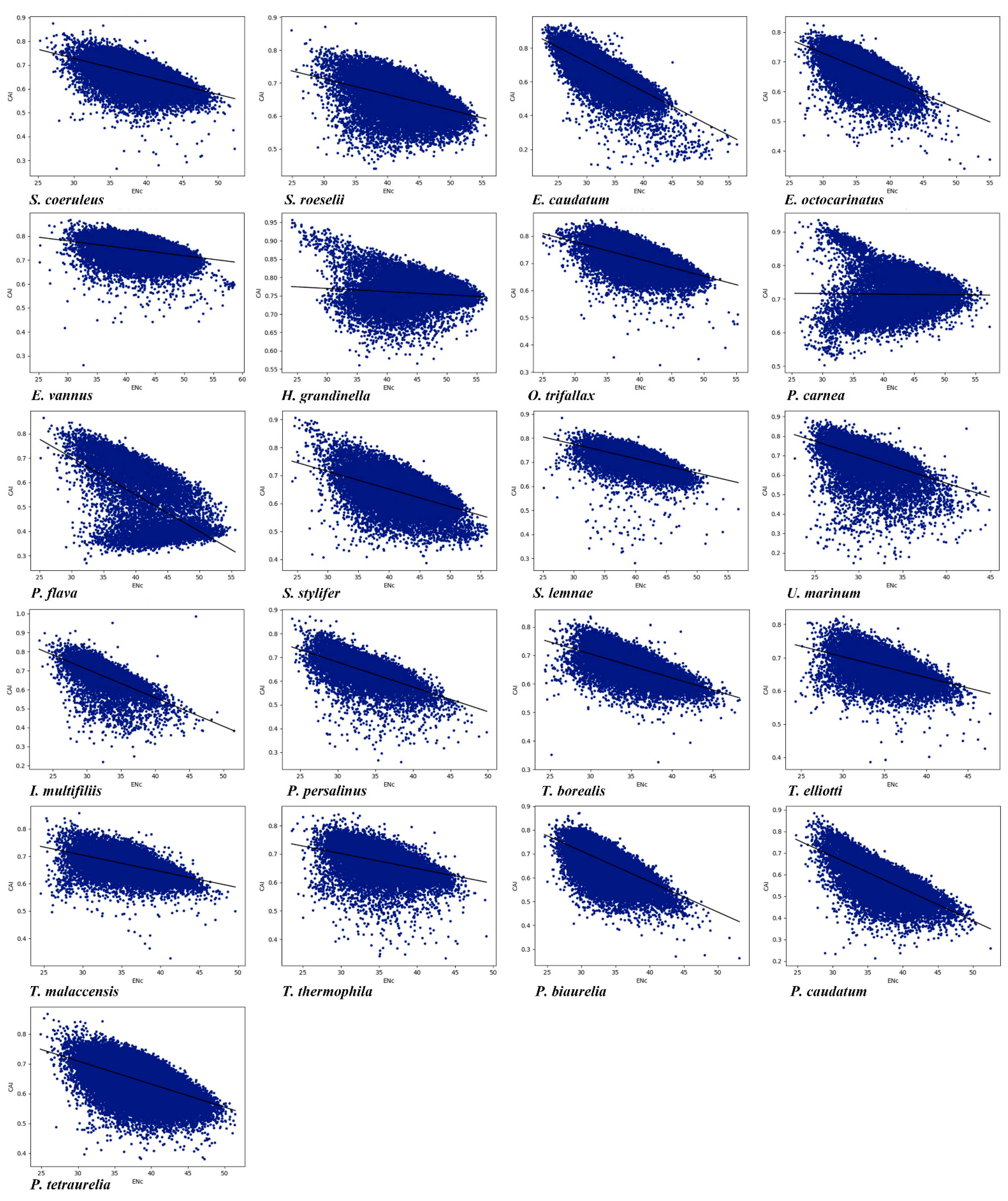
3.2. Effective Number of Codons and its Association with GC3
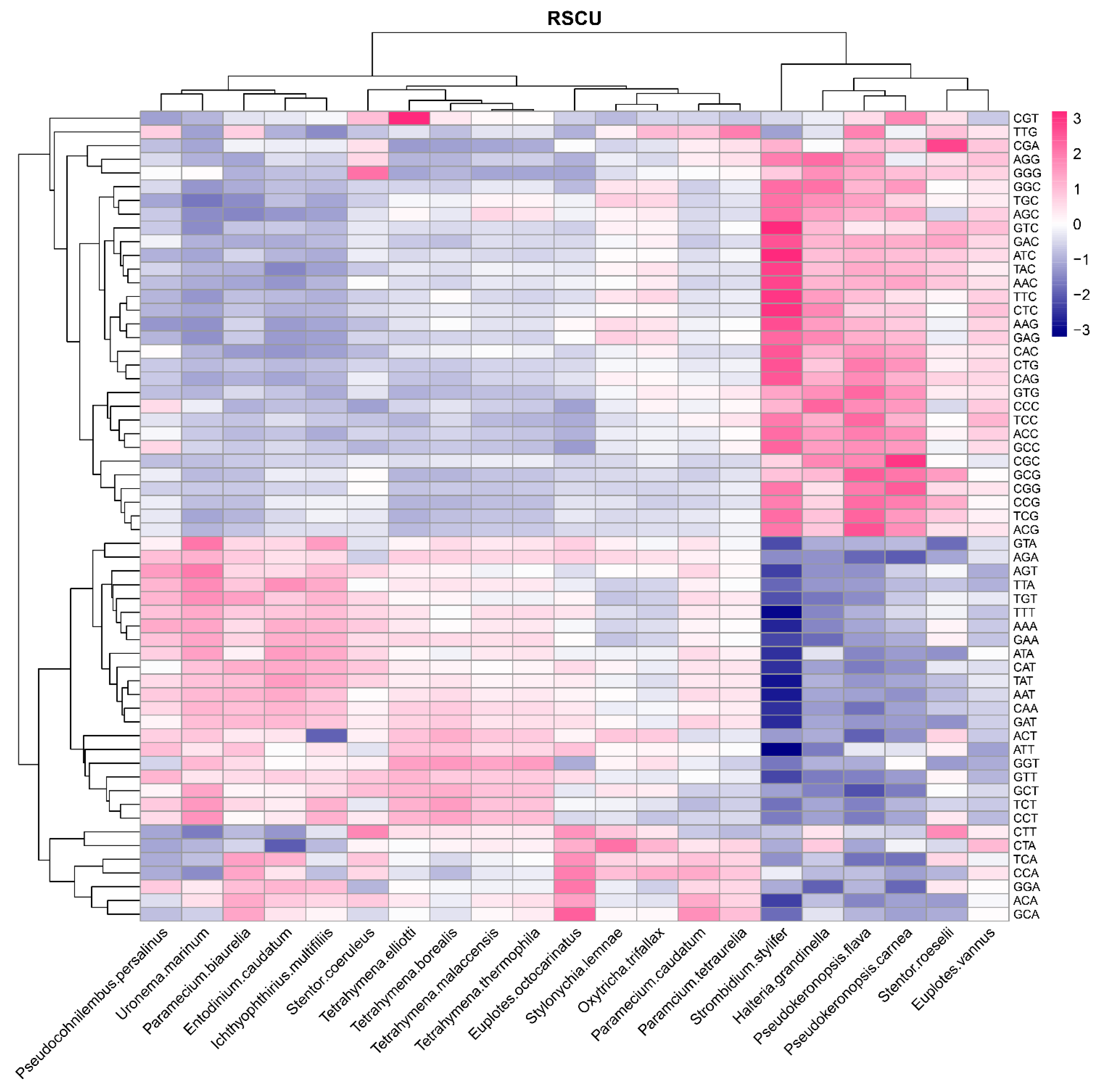
3.3. Relative Synonymous Codon Usage (RSCU) and Putative Optimal Codons
3.4. PR2-Plot Analysis
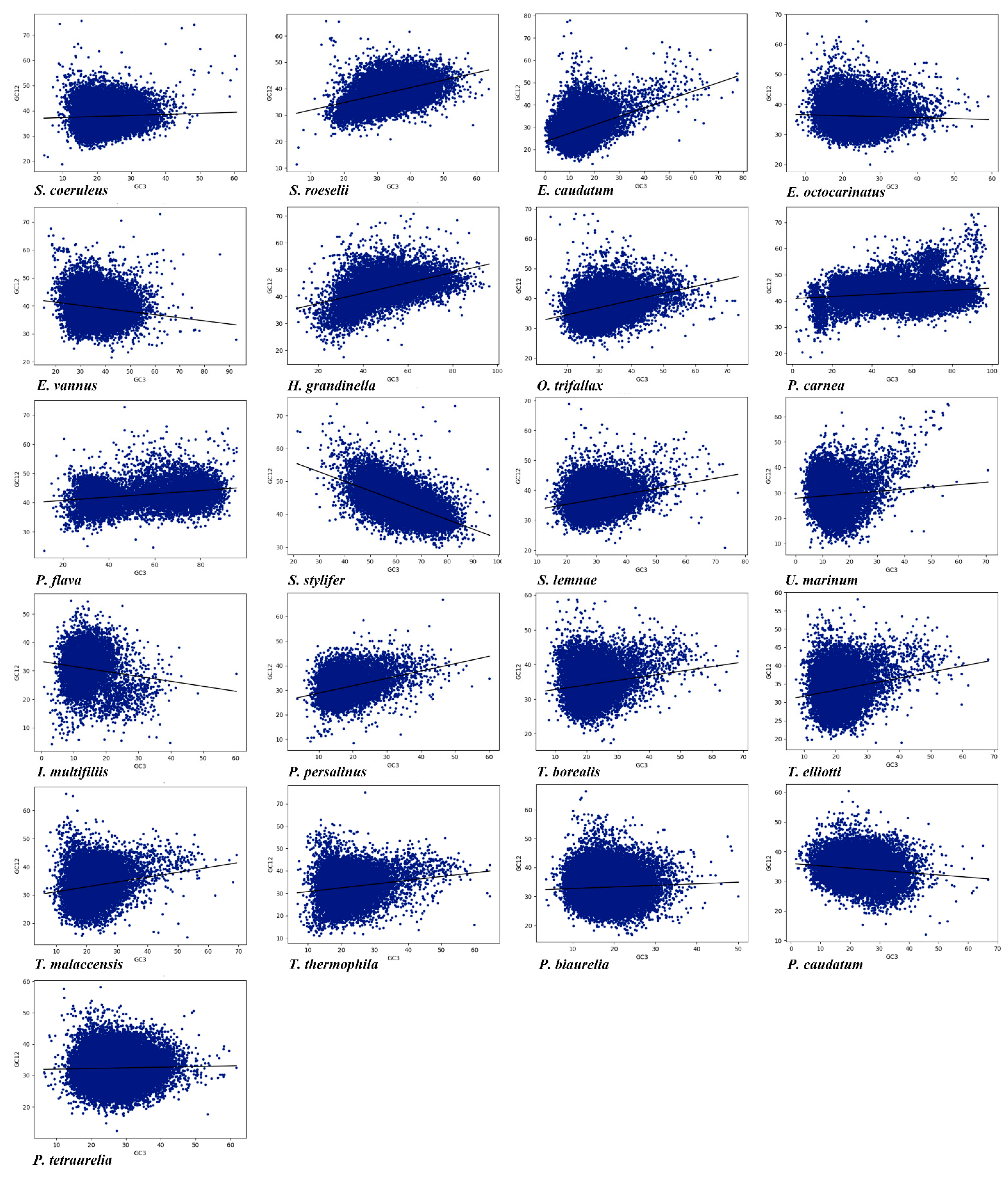
3.5. Neutrality Plot Analysis
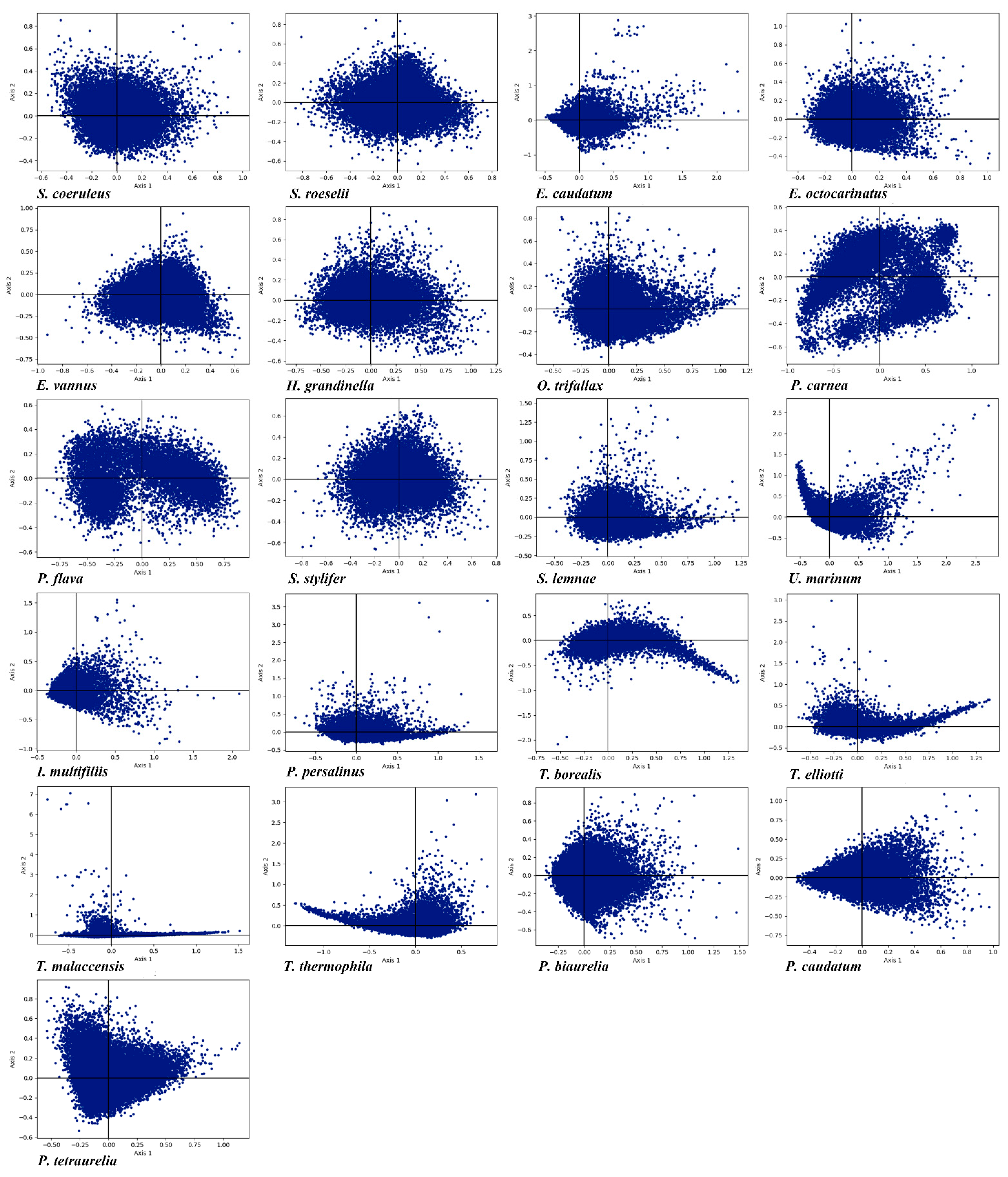
3.6. Correspondence Analysis
3.7. Prediction of Gene Expression in 21 Ciliates Species
3.8. Compositions and Gene Lengths of Amino Acids
3.9. Translation Selection (P2) and Choice between Pyrimidines in the 3rd Position of Codon
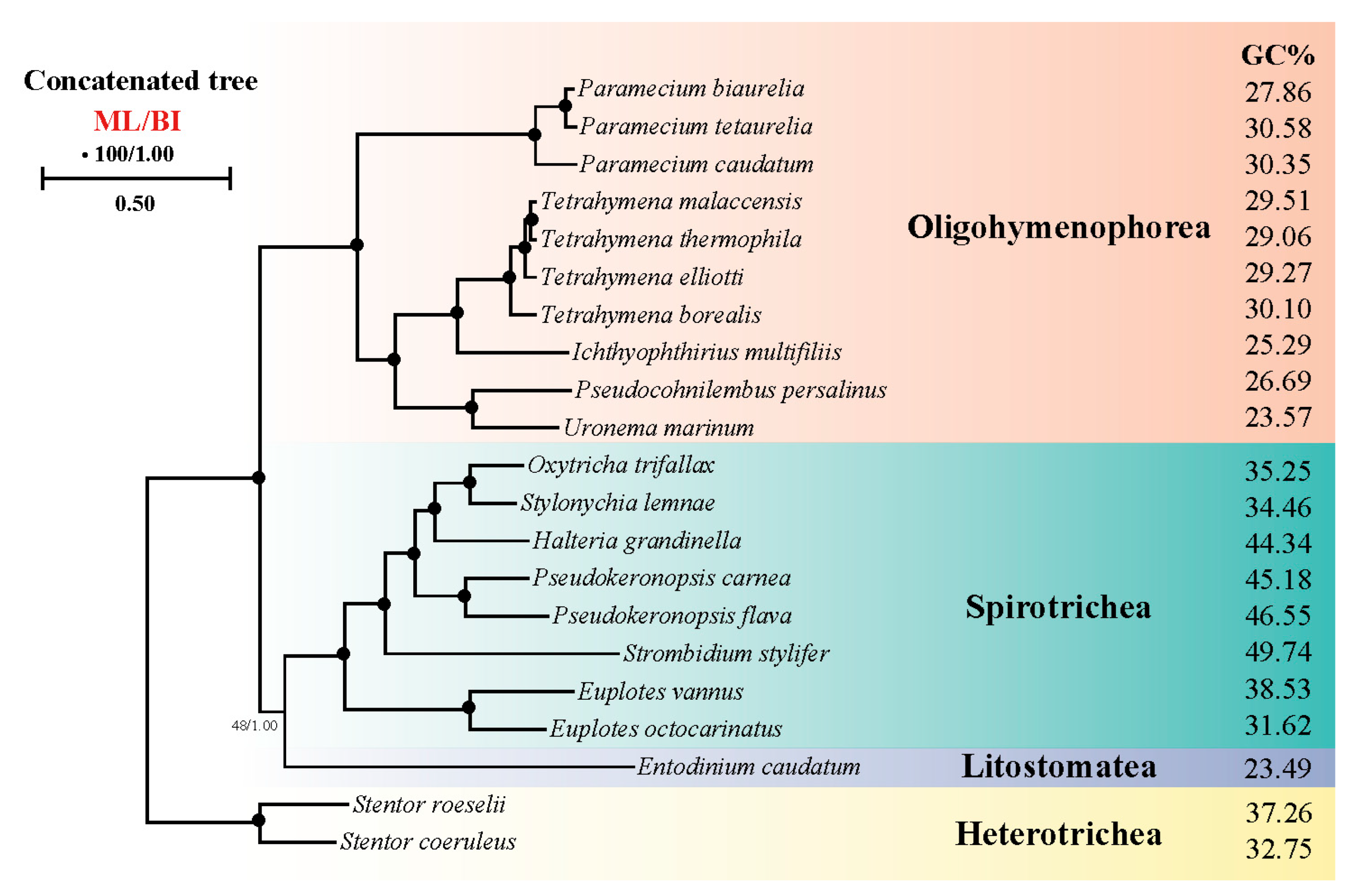
3.10. Phylogenomic Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ikemura, T. Codon usage and tRNA content in unicellular and multicellular organisms. Mol. Biol. Evol. 1985, 2, 13–34. [Google Scholar] [PubMed]
- Biro, J.C. Studies on the origin and evolution of codon bias. arXiv 2008, arXiv:0807.3901. [Google Scholar]
- Bulmer, M. The selection-mutation-drift theory of synonymous codon usage. Genetics 1991, 129, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Akashi, H.; Eyre-Walker, A. Translational selection and molecular evolution. Curr. Opin. Genet. Dev. 1998, 8, 688–693. [Google Scholar] [CrossRef]
- Akashi, H. Gene expression and molecular evolution. Curr. Opin. Genet. Dev. 2001, 11, 660–666. [Google Scholar] [CrossRef]
- Hershberg, R.; Petrov, D.A. Selection on codon bias. Annu. Rev. Genet. 2008, 42, 287–299. [Google Scholar]
- Ikemura, T. Correlation between the abundance of Escherichia coli transfer RNAs and the occurrence of the respective codons in its protein genes: A proposal for a synonymous codon choice that is optimal for the E. coli translational system. J. Mol. Biol. 1981, 151, 389–409. [Google Scholar] [CrossRef]
- Sharp, P.M.; Averof, M.; Lloyd, A.T.; Matassi, G.; Peden, J.F. DNA sequence evolution: The sounds of silence. Phil. Trans. R Soc. Lond. B Biol. Sci. 1995, 349, 241–247. [Google Scholar]
- Duncan, G.A.; Dunigan, D.D.; Van Etten, J.L.V. Diversity of tRNA clusters in the Chloroviruses. Viruses 2020, 12, 1173. [Google Scholar] [CrossRef]
- Moriyama, E.N.; Powell, J.R. Gene length and codon usage bias in Drosophila melanogaster, Saccharomyces cerevisiae and Escherichia coli. Nucleic Acids Res. 1998, 26, 3188–3193. [Google Scholar] [CrossRef]
- Eyre-Walker, A. Synonymous codon bias is related to gene length in Escherichia coli: Selection for translational accuracy? Mol. Biol. Evol. 1996, 13, 864–872. [Google Scholar] [CrossRef]
- Gouy, M.; Gautier, C. Codon usage in bacteria: Correlation with gene expressivity. Nucleic Acids Res. 1982, 10, 7055–7074. [Google Scholar] [CrossRef]
- Iannacone, R.; Grieco, P.D.; Cellini, F. Specific sequence modifications of a cry3B endotoxin gene result in high levels of expression and insect resistance. Plant Mol. Biol. 1997, 34, 485–496. [Google Scholar] [CrossRef]
- Mazumdar, P.; Binti Othman, R.; Mebus, K.; Ramakrishnan, N.; Ann Harikrishna, J. Codon usage and codon pair patterns in non-grass monocot genomes. Ann. Bot. 2017, 120, 893–909. [Google Scholar] [CrossRef] [PubMed]
- Galtier, N.; Roux, C.; Rousselle, M.; Romiguier, J.; Figuet, E.; Glémin, S.; Bierne, N.; Duret, L. Codon usage bias in animals: Disentangling the effects of natural selection, effective population size, and GC-biased gene conversion. Mol. Biol. Evol. 2018, 35, 1092–1103. [Google Scholar] [CrossRef]
- Parvathy, S.T.; Udayasuriyan, V.; Bhadana, V. Codon usage bias. Mol. Biol. Rep. 2022, 49, 539–565. [Google Scholar] [CrossRef]
- Kliman, R.M.; Hey, J. Reduced natural selection associated with low recombination in Drosophila melanogaster. Mol. Biol. Evol. 1993, 10, 1239–1258. [Google Scholar] [PubMed]
- Munté, A.; Aguadé, M.; Segarra, C. Divergence of the yellow gene between Drosophila melanogaster and D. subobscura: Recombination rate, codon bias and synonymous substitutions. Genetics 1997, 147, 165–175. [Google Scholar] [CrossRef]
- Zhou, T.; Lu, Z.H.; Sun, X. The correlation between recombination rate and codon bias in yeast mainly results from mutational bias associated with recombination rather than Hill-Robertson Interference. Conf. Proc. IEEE Eng. Med. Biol. Soc. 2005, 2005, 4787–4790. [Google Scholar] [PubMed]
- Zhou, J.; Wan, J.; Shu, X.E.; Mao, Y.; Liu, X.M.; Yuan, X.; Zhang, X.; Hess, M.E.; Brüning, J.C.; Qian, S.B. N6-methyladenosine guides mRNA alternative translation during integrated stress response. Mol. Cell 2018, 69, 636–647.e7. [Google Scholar] [CrossRef] [PubMed]
- Qing, G.; Xia, B.; Inouye, M. Enhancement of translation initiation by A/T-rich sequences downstream of the initiation codon in Escherichia coli. J. Mol. Microbiol. Biotechnol. 2003, 6, 133–144. [Google Scholar] [CrossRef]
- Foster, P.G.; Jermiin, L.S.; Hickey, D.A. Nucleotide composition bias affects amino acid content in proteins coded by animal mitochondria. J. Mol. Evol. 1997, 44, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.X.; Cao, X.; Ma, P.; Chang, Q.Y.; Li, L.J.; Zhou, X.K.; Zhang, D.R.; Li, M.S.; Ma, Z.R. Comparative genomic analysis for nucleotide, codon, and amino acid usage patterns of mycoplasmas. J. Basic Microbiol. 2018, 58, 425–439. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.Q.; Ning, L.W.; Zhang, H.X.; Guo, F.B. Analysis of the relationship between genomic GC Content and patterns of base usage, codon usage and amino acid usage in prokaryotes: Similar GC content adopts similar compositional frequencies regardless of the phylogenetic lineages. PLoS ONE 2014, 9, e107319. [Google Scholar] [CrossRef]
- Matsuo, Y. The adenine/thymine deleterious selection model for GC content evolution at the third codon position of the histone genes in Drosophila. Genes 2021, 12, 721. [Google Scholar] [CrossRef]
- Cavalcanti, A.R.; Stover, N.A.; Orecchia, L.; Doak, T.G.; Landweber, L.F. Coding properties of Oxytricha trifallax (Sterkiella histriomuscorum) macronuclear chromosomes: Analysis of a pilot genome project. Chromosoma 2004, 113, 69–76. [Google Scholar] [CrossRef]
- Seffens, W.; Digby, D. mRNAs have greater negative folding free energies than shuffled or codon choice randomized sequences. Nucleic Acids Res. 1999, 27, 1578–1584. [Google Scholar] [CrossRef] [PubMed]
- Kahali, B.; Basak, S.; Ghosh, T.C. Reinvestigating the codon and amino acid usage of S. cerevisiae genome: A new insight from protein secondary structure analysis. Biochem. Biophys. Res. Commun. 2007, 354, 693–699. [Google Scholar] [CrossRef]
- Qin, H.; Wu, W.B.; Comeron, J.M.; Kreitman, M.; Li, W.H. Intragenic spatial patterns of codon usage bias in prokaryotic and eukaryotic genomes. Genetics 2004, 168, 2245–2260. [Google Scholar] [CrossRef]
- Zhao, F.; Zhou, Z.; Dang, Y.; Na, H.; Adam, C.; Lipzen, A.; Ng, V.; Grigoriev, I.V.; Liu, Y. Genome-wide role of codon usage on transcription and identification of potential regulators. Proc. Natl. Acad. Sci. USA 2021, 118, e2022590118. [Google Scholar] [CrossRef]
- Salim, H.M.; Cavalcanti, A.R. Factors influencing codon usage bias in genomes. J. Braz. Chem. Soc. 2008, 19, 257–262. [Google Scholar] [CrossRef]
- Tuller, T.; Waldman, Y.Y.; Kupiec, M.; Ruppin, E. Translation efficiency is determined by both codon bias and folding energy. Proc. Natl. Acad. Sci. USA 2010, 107, 3645–3650. [Google Scholar] [CrossRef] [PubMed]
- Kurland, C.G. Translational accuracy and the fitness of bacteria. Annu. Rev. Genet. 1992, 26, 29–50. [Google Scholar] [CrossRef] [PubMed]
- Lyu, X.; Liu, Y. Nonoptimal codon usage is critical for protein structure and function of the master general amino acid control regulator CPC-1. mBio 2020, 11, e02605-20. [Google Scholar] [CrossRef] [PubMed]
- Goetz, R.M.; Fuglsang, A. Correlation of codon bias measures with mRNA levels: Analysis of transcriptome data from Escherichia coli. Biochem. Biophys. Res. Commun. 2005, 327, 4–7. [Google Scholar] [CrossRef]
- Frumkin, I.; Lajoie, M.J.; Gregg, C.J.; Hornung, G.; Church, G.M.; Pilpel, Y. Codon usage of highly expressed genes affects proteome-wide translation efficiency. Proc. Natl. Acad. Sci. USA 2018, 115, E4940–E4949. [Google Scholar] [CrossRef]
- Konczal, J.; Bower, J.; Gray, C.H. Re-introducing non-optimal synonymous codons into codon-optimized constructs enhances soluble recovery of recombinant proteins from Escherichia coli. PLoS ONE 2019, 14, e0215892. [Google Scholar] [CrossRef]
- Weiner, I.; Atar, S.; Schweitzer, S.; Eilenberg, H.; Feldman, Y.; Avitan, M.; Blau, M.; Danon, A.; Tuller, T.; Yacoby, I. Enhancing heterologous expression in Chlamydomonas reinhardtii by transcript sequence optimization. Plant J. 2018, 94, 22–31. [Google Scholar] [CrossRef]
- Han, Z.; Lo, W.S.; Lightfoot, J.W.; Witte, H.; Sun, S.; Sommer, R.J. Improving transgenesis efficiency and CRISPR-associated tools through codon optimization and native intron addition in pristionchus nematodes. Genetics 2020, 216, 947–956. [Google Scholar] [CrossRef]
- Kucho, K.; Kakoi, K.; Yamaura, M.; Iwashita, M.; Abe, M.; Uchiumi, T. Codon-optimized antibiotic resistance gene improves efficiency of transient transformation in Frankia. J. Biosci. 2013, 38, 713–717. [Google Scholar] [CrossRef]
- Ingvarsson, P.K. Molecular evolution of synonymous codon usage in Populus. BMC Evol. Biol. 2008, 8, 307. [Google Scholar] [CrossRef]
- Wang, B.; Yuan, J.; Liu, J.; Jin, L.; Chen, J.Q. Codon usage bias and determining forces in green plant mitochondrial genomes. J. Integr. Plant Biol. 2011, 53, 324–334. [Google Scholar] [CrossRef]
- Zhang, P.; Xu, W.; Lu, X.; Wang, L. Analysis of codon usage bias of chloroplast genomes in Gynostemma species. Physiol. Mol. Biol. Plants. 2021, 27, 2727–2737. [Google Scholar] [CrossRef]
- Lee, S.; Basu, S.; Tyler, C.; Wei, I.W. Ciliate populations as bio-indicators at Deer Island Treatment Plant. Adv. Environ. Res. 2004, 8, 371–378. [Google Scholar] [CrossRef]
- Chen, Q.H.; Xu, R.L.; Tam, N.F.; Cheung, S.G.; Shin, P.K. Use of ciliates (Protozoa: Ciliophora) as bioindicator to assess sediment quality of two constructed mangrove sewage treatment belts in southern China. Mar. Pollut. Bull. 2008, 57, 689–694. [Google Scholar] [CrossRef]
- Chariton, A.A.; Stephenson, S.; Morgan, M.J.; Steven, A.D.L.; Colloff, M.J.; Court, L.N.; Hardy, C.M. Metabarcoding of benthic eukaryote communities predicts the ecological condition of estuaries. Environ. Pollut. 2015, 203, 165–174. [Google Scholar] [CrossRef]
- Dohra, H.; Fujishima, M.; Suzuki, H. Analysis of amino acid and codon usage in Paramecium bursaria. FEBS Lett. 2015, 589, 3113–3118. [Google Scholar] [CrossRef]
- Salim, H.M.; Ring, K.L.; Cavalcanti, A.R. Patterns of codon usage in two ciliates that reassign the genetic code: Tetrahymena thermophila and Paramecium tetraurelia. Protist 2008, 159, 283–298. [Google Scholar] [CrossRef]
- Wuitschick, J.D.; Karrer, K.M. Analysis of genomic G + C content, codon usage, initiator codon context and translation termination sites in Tetrahymena thermophila. J. Eukaryot. Microbiol. 1999, 46, 239–247. [Google Scholar] [CrossRef]
- Barth, D.; Berendonk, T.U. The mitochondrial genome sequence of the ciliate Paramecium caudatum reveals a shift in nucleotide composition and codon usage within the genus Paramecium. BMC Genomics. 2011, 12, 272. [Google Scholar] [CrossRef]
- Eschenlauer, S.C.; McEwan, N.R.; Calza, R.E.; Wallace, R.J.; Onodera, R.; Newbold, C.J. Phylogenetic position and codon usage of two centrin genes from the rumen ciliate protozoan, Entodinium caudatum. FEMS Microbiol. Lett. 1998, 166, 147–154. [Google Scholar] [CrossRef]
- Larsen, L.K.; Andreasen, P.H.; Dreisig, H.; Palm, L.; Nielsen, H.; Engberg, J.; Kristiansen, K. Cloning and characterization of the gene encoding the highly expressed ribosomal protein l3 of the ciliated protozoan Tetrahymena thermophila. Evidence for differential codon usage in highly expressed genes. Cell Biol. Int. 1999, 23, 551–560. [Google Scholar] [CrossRef]
- Wang, Y.; Yao, L.; Fan, J.; Zhao, X.; Zhang, Q.; Chen, Y.; Guo, C. The codon usage bias analysis of free-living ciliates’ macronuclear genomes and clustered regularly interspaced short palindromic repeats/Cas9 vector construction of Stylonychia lemnae. Front. Microbiol. 2022, 13, 785889. [Google Scholar] [CrossRef]
- Tourancheau, A.B.; Tsao, N.; Klobutcher, L.A.; Pearlman, R.E.; Adoutte, A. Genetic code deviations in the ciliates: Evidence for multiple and independent events. EMBO J. 1995, 14, 3262–3267. [Google Scholar] [CrossRef]
- Anwar, A.M. BCAWT: Automated tool for codon usage bias analysis for molecular evolution. J. Open Source Softw. 2019, 4, 1500. [Google Scholar] [CrossRef]
- Wright, F. The ‘effective number of codons’ used in a gene. Gene 1990, 87, 23–29. [Google Scholar] [CrossRef]
- Comeron, J.M.; Aguadé, M. An evaluation of measures of synonymous codon usage bias. J. Mol. Evol. 1998, 47, 268–274. [Google Scholar] [CrossRef]
- Sun, X.; Yang, Q.; Xia, X. An improved implementation of effective number of codons (Nc). Mol. Biol. Evol. 2013, 30, 191–196. [Google Scholar] [CrossRef]
- Kawabe, A.; Miyashita, N.T. Patterns of codon usage bias in three dicot and four monocot plant species. Genes Genet. Syst. 2003, 78, 343–352. [Google Scholar] [CrossRef]
- Zhang, W.J.; Zhou, J.; Li, Z.F.; Wang, L.; Gu, X.; Zhong, Y. Comparative analysis of codon usage patterns among mitochondrion, chloroplast and nuclear genes in Triticum aestivum L. J. Integr. Plant Biol. 2007, 49, 246–254. [Google Scholar] [CrossRef]
- Sharp, P.M.; Li, W.H. The codon adaptation index--a measure of directional synonymous codon usage bias, and its potential applications. Nucleic Acids Res. 1987, 15, 1281–1295. [Google Scholar] [CrossRef]
- Sharp, P.M.; Tuohy, T.M.; Mosurski, K.R. Codon usage in yeast: Cluster analysis clearly differentiates highly and lowly expressed genes. Nucleic Acids Res. 1986, 14, 5125–5143. [Google Scholar] [CrossRef]
- Wong, E.H.; Smith, D.K.; Rabadan, R.; Peiris, M.; Poon, L.L. Codon usage bias and the evolution of influenza A viruses. codon usage biases of influenza virus. BMC Evol. Biol. 2010, 10, 253. [Google Scholar] [CrossRef]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef]
- Lobry, J.R.; Gautier, C. Hydrophobicity, expressivity and aromaticity are the major trends of amino-acid usage in 999 Escherichia coli chromosome-encoded genes. Nucleic Acids Res. 1994, 22, 3174–3180. [Google Scholar] [CrossRef]
- Zhou, M.; Long, W.; Li, X. Patterns of synonymous codon usage bias in chloroplast genomes of seed plants. For. Stud. China 2008, 10, 235–242. [Google Scholar] [CrossRef]
- Sueoka, N. Translation-coupled violation of Parity Rule 2 in human genes is not the cause of heterogeneity of the DNA G+C content of third codon position. Gene 1999, 238, 53–58. [Google Scholar] [CrossRef]
- Chakraborty, S.; Nag, D.; Mazumder, T.H.; Uddin, A. Codon usage pattern and prediction of gene expression level in Bungarus species. Gene 2017, 604, 48–60. [Google Scholar] [CrossRef]
- Sueoka, N. Directional mutation pressure and neutral molecular evolution. Proc. Natl. Acad. Sci. USA 1988, 85, 2653–2657. [Google Scholar] [CrossRef]
- Sueoka, N. Directional mutation pressure, mutator mutations, and dynamics of molecular evolution. J. Mol. Evol. 1993, 37, 137–153. [Google Scholar] [CrossRef]
- Emms, D.M.; Kelly, S. OrthoFinder: Phylogenetic orthology inference for comparative genomics. Genome Biol. 2019, 20, 238. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Lartillot, N.; Rodrigue, N.; Stubbs, D.; Richer, J. PhyloBayes MPI: Phylogenetic reconstruction with infinite mixtures of profiles in a parallel environment. Syst. Biol. 2013, 62, 611–615. [Google Scholar] [CrossRef]
- Plotkin, J.B.; Kudla, G. Synonymous but not the same: The causes and consequences of codon bias. Nat. Rev. Genet. 2011, 12, 32–42. [Google Scholar] [CrossRef]
- Jenkins, G.M.; Holmes, E.C. The extent of codon usage bias in human RNA viruses and its evolutionary origin. Virus Res. 2003, 92, 1–7. [Google Scholar] [CrossRef]
- Sueoka, N. Intrastrand parity rules of DNA base composition and usage biases of synonymous codons. J. Mol. Evol. 1995, 40, 318–325. [Google Scholar] [CrossRef]
- Chen, H.; Sun, S.; Norenburg, J.L.; Sundberg, P. Mutation and selection cause codon usage and bias in mitochondrial genomes of ribbon worms (Nemertea). PLoS ONE 2014, 9, e85631. [Google Scholar] [CrossRef]
- Liu, H.; Huang, Y.; Du, X.; Chen, Z.; Zeng, X.; Chen, Y.; Zhang, H. Patterns of synonymous codon usage bias in the model grass Brachypodium distachyon. Genet. Mol. Res. 2012, 11, 4695–4706. [Google Scholar] [CrossRef]
- Fassbinder, J. Correspondence Analysis Handbook: Computational Statistics & Data Analysis; Benzecri, J.-P., Ed.; Elsevier: Amsterdam, The Netherlands, 1996; Volume 21, pp. 374–375. [Google Scholar]
- Zhou, T.; Gu, W.; Ma, J.; Sun, X.; Lu, Z. Analysis of synonymous codon usage in H5N1 virus and other influenza A viruses. Biosystems 2005, 81, 77–86. [Google Scholar] [CrossRef]
- Bennetzen, J.L.; Hall, B.D. Codon selection in yeast. J. Biol. Chem. 1982, 257, 3026–3031. [Google Scholar] [CrossRef]
- Wang, L.; Xing, H.; Yuan, Y.; Wang, X.; Saeed, M.; Tao, J.; Feng, W.; Zhang, G.; Song, X.; Sun, X. Genome-wide analysis of codon usage bias in four sequenced cotton species. PLoS ONE 2018, 13, e0194372. [Google Scholar] [CrossRef]
- Sharp, P.M.; Matassi, G. Codon usage and genome evolution. Curr. Opin. Genet. Dev. 1994, 4, 851–860. [Google Scholar] [CrossRef]
- Crick, F.H. The origin of the genetic code. J. Mol. Biol. 1968, 38, 367–379. [Google Scholar] [CrossRef]
- Kanaya, S.; Yamada, Y.; Kinouchi, M.; Kudo, Y.; Ikemura, T. Codon usage and tRNA genes in eukaryotes: Correlation of codon usage diversity with translation efficiency and with CG-dinucleotide usage as assessed by multivariate analysis. J. Mol. Evol. 2001, 53, 290–298. [Google Scholar] [CrossRef]
- Coghlan, A.; Wolfe, K.H. Relationship of codon bias to mRNA concentration and protein length in Saccharomyces cerevisiae. Yeast 2000, 16, 1131–1145. [Google Scholar] [CrossRef]
- Wang, L.; Roossinck, M.J. Comparative analysis of expressed sequences reveals a conserved pattern of optimal codon usage in plants. Plant Mol. Biol. 2006, 61, 699–710. [Google Scholar] [CrossRef]
- Eisen, J.A.; Coyne, R.S.; Wu, M.; Wu, D.; Thiagarajan, M.; Wortman, J.R.; Badger, J.H.; Ren, Q.; Amedeo, P.; Jones, K.M.; et al. Macronuclear genome sequence of the ciliate Tetrahymena thermophila, a model eukaryote. PLoS Biol. 2006, 4, e286. [Google Scholar] [CrossRef]
- Sharp, P.M.; Stenico, M.; Peden, J.F.; Lloyd, A.T. Codon usage: Mutational bias.; translational selection.; or both? Biochem. Soc. Trans. 1993, 21, 835–841. [Google Scholar] [CrossRef]
- Marais, G. Biased gene conversion: Implications for genome and sex evolution. Trends Genet. 2003, 19, 330–338. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | A3% | T3% | C3% | G3% | GC% | GC1% | GC2% | GC12% | GC3(s)% |
|---|---|---|---|---|---|---|---|---|---|
| Stentor roeselii | 31.91 | 33.87 | 19.34 | 14.88 | 37.26 | 44.26 | 33.29 | 38.77 | 34.22 |
| Stentor coeruleus | 38.05 | 39.29 | 12.4 | 10.25 | 32.75 | 42.79 | 32.80 | 37.79 | 22.66 |
| Euplotes vannus | 33.13 | 30.16 | 20.20 | 16.51 | 38.53 | 41.45 | 37.48 | 39.45 | 36.71 |
| Euplotes octocarinatus | 39.51 | 37.90 | 12.14 | 10.45 | 31.62 | 41.56 | 30.70 | 36.13 | 22.59 |
| Strombidium stylifer | 19.40 | 18.61 | 35.78 | 26.20 | 49.74 | 52.32 | 34.91 | 43.61 | 61.98 |
| Halteria grandinella | 25.13 | 27.26 | 27.21 | 20.40 | 44.34 | 48.67 | 36.72 | 42.70 | 47.61 |
| Stylonychia lemnae | 33.23 | 37.19 | 16.69 | 12.89 | 34.46 | 42.10 | 31.70 | 36.90 | 29.58 |
| Oxytricha trifallax | 33.15 | 35.51 | 18.31 | 12.95 | 35.25 | 42.32 | 32.15 | 37.21 | 31.30 |
| Pseudokeronopsis flava | 22.32 | 23.32 | 28.54 | 25.82 | 46.55 | 50.97 | 34.32 | 42.65 | 54.36 |
| Pseudokeronopsis carnea | 23.81 | 26.40 | 27.60 | 22.20 | 45.18 | 51.67 | 34.07 | 42.87 | 49.79 |
| Entodinium caudatum | 42.95 | 43.72 | 7.61 | 5.78 | 23.49 | 32.93 | 24.20 | 28.56 | 13.33 |
| Tetrahymena borealis | 34.45 | 43.87 | 13.60 | 8.08 | 30.10 | 37.88 | 30.74 | 34.31 | 21.68 |
| Tetrahymena elliotti | 35.75 | 43.37 | 13.24 | 7.65 | 29.27 | 37.30 | 29.63 | 33.46 | 20.88 |
| Tetrahymena malaccensis | 35.54 | 42.38 | 14.05 | 8.03 | 29.51 | 36.89 | 29.56 | 33.22 | 22.09 |
| Tetrahymena thermophila | 36.89 | 36.06 | 14.57 | 12.49 | 29.06 | 36.10 | 29.32 | 32.71 | 21.76 |
| Ichthyophthirius multifiliis | 41.19 | 44.54 | 8.22 | 6.05 | 25.29 | 34.70 | 26.84 | 30.77 | 14.27 |
| Pseudocohnilembus persalinus | 39.94 | 42.21 | 11.04 | 6.81 | 26.69 | 35.65 | 26.56 | 31.10 | 17.85 |
| Uronema marinum | 41.76 | 45.43 | 7.87 | 4.95 | 23.57 | 32.83 | 25.09 | 28.96 | 12.80 |
| Paramecium biaurelia | 40.84 | 41.88 | 8.96 | 8.31 | 27.86 | 37.04 | 29.27 | 33.15 | 17.00 |
| Paramecium caudatum | 39.35 | 38.08 | 11.91 | 10.67 | 30.35 | 38.38 | 30.09 | 34.24 | 22.57 |
| Paramcium tetraurelia | 36.89 | 36.06 | 14.57 | 12.49 | 30.58 | 35.14 | 29.56 | 32.35 | 27.06 |
| Species | Range | Mean | SD |
|---|---|---|---|
| Stentor roeselii | 24.8233~55.6122 | 43.2310 | 4.7995 |
| Stentor coeruleus | 25.2281~52.2843 | 39.4773 | 3.3917 |
| Euplotes vannus | 25.1212~58.6122 | 44.2310 | 3.9264 |
| Euplotes octocarinatus | 25.8303~55.0067 | 36.4383 | 2.8887 |
| Strombidium stylifer | 24.1399~56.0129 | 42.6091 | 4.7208 |
| Halteria grandinella | 24.0406~56.6128 | 44.8589 | 5.4764 |
| Stylonychia lemnae | 25.0396~56.7110 | 40.5681 | 3.4984 |
| Oxytricha trifallax | 25.0583~55.2462 | 41.5530 | 3.5766 |
| Pseudokeronopsis flava | 25.1127~55.5793 | 41.8680 | 5.2695 |
| Pseudokeronopsis carnea | 25.4563~57.3212 | 42.6069 | 5.2678 |
| Entodinium caudatum | 22.1304~56.4895 | 33.2936 | 3.5350 |
| Tetrahymena borealis | 54.3635~48.3126 | 36.2745 | 3.2871 |
| Tetrahymena elliotti | 24.2852~47.6539 | 35.8999 | 3.0246 |
| Tetrahymena malaccensis | 24.5190~49.6401 | 36.6933 | 3.4687 |
| Tetrahymena thermophila | 23.4909~49.0903 | 36.5193 | 3.3741 |
| Ichthyophthirius multifiliis | 22.9875~51.5511 | 33.0615 | 3.3160 |
| Pseudocohnilembus persalinus | 23.8975~49.8253 | 34.2206 | 3.4134 |
| Uronema marinum | 24.8432~51.4703 | 38.8706 | 3.8384 |
| Paramecium biaurelia | 24.5789~53.1388 | 34.2286 | 2.7692 |
| Paramecium caudatum | 24.7160~52.5174 | 34.2473 | 3.8172 |
| Paramcium tetraurelia | 22.6886~44.8700 | 31.4826 | 2.5502 |
| Species | Axis1 | Axis1-4 |
|---|---|---|
| Stentor roeselii | 11.40% | 31.06% |
| Stentor coeruleus | 7.30% | 29.02% |
| Euplotes vannus | 8.46% | 25.84% |
| Euplotes octocarinatus | 5.82% | 31.80% |
| Strombidium stylifer | 8.71% | 34.37% |
| Halteria grandinella | 18.48% | 36.86% |
| Stylonychia lemnae | 9.72% | 24.75% |
| Oxytricha trifallax | 13.49% | 23.71% |
| Pseudokeronopsis flava | 32.40% | 49.76% |
| Pseudokeronopsis carnea | 37.59% | 48.13% |
| Entodinium caudatum | 9.07% | 36.80% |
| Tetrahymena borealis | 16.41% | 30.54% |
| Tetrahymena elliotti | 13.46% | 31.80% |
| Tetrahymena malaccensis | 11.74% | 34.70% |
| Tetrahymena thermophila | 10.54% | 36.96% |
| Ichthyophthirius multifiliis | 9.23% | 48.71% |
| Pseudocohnilembus persalinus | 16.36% | 55.11% |
| Uronema marinum | 10.86% | 51.81% |
| Paramecium biaurelia | 7.01% | 34.35% |
| Paramecium caudatum | 11.00% | 34.63% |
| Paramcium tetraurelia | 8.08% | 36.31% |
| Species | Range | Mean | SD |
|---|---|---|---|
| Stentor roeselii | 0.4404~0.8811 | 0.6505 | 0.0516 |
| Stentor coeruleus | 0.2648~0.8749 | 0.6570 | 0.0502 |
| Euplotes vannus | 0.2612~0.8672 | 0.736 | 0.0344 |
| Euplotes octocarinatus | 0.334~0.8298 | 0.6700 | 0.0469 |
| Strombidium stylifer | 0.3869~0.9063 | 0.6354 | 0.0583 |
| Halteria grandinella | 0.5605~0.9565 | 0.7572 | 0.0391 |
| Stylonychia lemnae | 0.2803~0.8844 | 0.7117 | 0.0401 |
| Oxytricha trifallax | 0.3252~0.8593 | 0.7054 | 0.0402 |
| Pseudokeronopsis flava | 0.2695~0.8644 | 0.5226 | 0.1326 |
| Pseudokeronopsis carnea | 0.5025~0.9352 | 0.7142 | 0.0610 |
| Entodinium caudatum | 0.084~0.9423 | 0.6605 | 0.0832 |
| Tetrahymena borealis | 0.3253~0.8352 | 0.6524 | 0.0447 |
| Tetrahymena elliotti | 0.3861~0.8234 | 0.6663 | 0.0375 |
| Tetrahymena malaccensis | 0.3268~0.8574 | 0.6643 | 0.0381 |
| Tetrahymena thermophila | 0.3325~0.8438 | 0.6673 | 0.0433 |
| Ichthyophthirius multifiliis | 0.2185~0.9845 | 0.6617 | 0.0833 |
| Pseudocohnilembus persalinus | 0.2584~0.8712 | 0.6359 | 0.0593 |
| Uronema marinum | 0.1479~0.8936 | 0.6806 | 0.0812 |
| Paramecium biaurelia | 0.2625~0.8712 | 0.6574 | 0.0595 |
| Paramecium caudatum | 0.2127~0.8852 | 0.5774 | 0.0848 |
| Paramcium tetraurelia | 0.3803~0.8669 | 0.6405 | 0.0563 |
| Species | WWC | WWT | WWY | SST | SSC | SSY | P2 |
|---|---|---|---|---|---|---|---|
| Stentor roeselii | 37.69 | 55.54 | 93.23 | 20.44 | 7.57 | 28.01 | 0.4906 |
| Stentor coeruleus | 22.70 | 63.02 | 85.72 | 24.25 | 4.68 | 28.93 | 0.4190 |
| Euplotes vannus | 34.21 | 50.08 | 84.29 | 17.56 | 10.57 | 28.13 | 0.4559 |
| Euplotes octocarinatus | 23.14 | 65.13 | 88.27 | 15.17 | 3.01 | 18.18 | 0.3680 |
| Strombidium stylifer | 45.48 | 15.65 | 61.13 | 13.36 | 16.06 | 29.42 | 0.6492 |
| Halteria grandinella | 36.46 | 36.62 | 73.08 | 17.05 | 19.90 | 36.95 | 0.4794 |
| Stylonychia lemnae | 41.92 | 84.27 | 126.19 | 25.99 | 9.94 | 35.93 | 0.4222 |
| Oxytricha trifallax | 47.00 | 81.83 | 128.83 | 27.04 | 11.46 | 38.50 | 0.4454 |
| Pseudokeronopsis flava | 34.85 | 37.49 | 72.34 | 14.95 | 17.14 | 32.09 | 0.5006 |
| Pseudokeronopsis carnea | 46.97 | 54.43 | 101.40 | 28.16 | 26.77 | 54.93 | 0.5101 |
| Entodinium caudatum | 20.02 | 134.57 | 154.59 | 17.74 | 4.13 | 21.87 | 0.2231 |
| Tetrahymena borealis | 40.34 | 118.10 | 158.44 | 36.07 | 7.58 | 43.65 | 0.3850 |
| Tetrahymena elliotti | 36.80 | 113.12 | 149.92 | 31.12 | 6.27 | 37.39 | 0.3688 |
| Tetrahymena malaccensis | 36.84 | 105.11 | 141.95 | 28.84 | 6.53 | 35.37 | 0.3712 |
| Tetrahymena thermophila | 37.91 | 115.27 | 153.18 | 29.81 | 6.46 | 36.27 | 0.3623 |
| Ichthyophthirius multifiliis | 15.66 | 88.17 | 103.83 | 19.21 | 3.73 | 22.94 | 0.2805 |
| Pseudocohnilembus persalinus | 23.97 | 106.22 | 130.19 | 18.74 | 7.57 | 26.31 | 0.2761 |
| Uronema marinum | 22.8 | 137.77 | 160.57 | 25.51 | 4.25 | 29.76 | 0.2679 |
| Paramecium biaurelia | 18.54 | 86.69 | 105.23 | 19.38 | 3.6 | 22.98 | 0.3057 |
| Paramecium caudatum | 22.94 | 77.54 | 100.48 | 18.01 | 5.26 | 23.27 | 0.3348 |
| Paramcium tetraurelia | 28.64 | 77.53 | 106.17 | 13.69 | 6.02 | 19.71 | 0.3346 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, Y.; Liang, F.; Li, C.; Warren, A.; Shin, M.K.; Li, L. Codon Usage Bias Analysis in Macronuclear Genomes of Ciliated Protozoa. Microorganisms 2023, 11, 1833. https://doi.org/10.3390/microorganisms11071833
Fu Y, Liang F, Li C, Warren A, Shin MK, Li L. Codon Usage Bias Analysis in Macronuclear Genomes of Ciliated Protozoa. Microorganisms. 2023; 11(7):1833. https://doi.org/10.3390/microorganisms11071833
Chicago/Turabian StyleFu, Yu, Fasheng Liang, Congjun Li, Alan Warren, Mann Kyoon Shin, and Lifang Li. 2023. "Codon Usage Bias Analysis in Macronuclear Genomes of Ciliated Protozoa" Microorganisms 11, no. 7: 1833. https://doi.org/10.3390/microorganisms11071833
APA StyleFu, Y., Liang, F., Li, C., Warren, A., Shin, M. K., & Li, L. (2023). Codon Usage Bias Analysis in Macronuclear Genomes of Ciliated Protozoa. Microorganisms, 11(7), 1833. https://doi.org/10.3390/microorganisms11071833