
Immunostimulatory Activity of Lactococcus lactis subsp. lactis CAB701 Isolated from Jeju Cabbage


Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation of Lactic Acid Bacteria Strains and Growth Conditions
2.2. Culture of Animal Cells
2.2.1. Cell Lines and Culture Conditions
2.2.2. Preparation of the Lactic Acid Bacterial Sample
2.3. Identification of Lactic Acid Bacteria
2.3.1. Nucleotide Sequencing of 16S rRNA Gene
2.3.2. Scanning Electron Microscopy
2.3.3. Carbohydrate Utilization Pattern of Lactic Acid Bacteria
2.4. Evaluation of the Probiotic Characteristics of Lactic Acid Bacteria
2.4.1. Acid Tolerance
2.4.2. Bile Tolerance
2.4.3. Pancreatin Tolerance
2.4.4. Gut Adhesion Ability
2.4.5. Antioxidant Activity
2.5. Evaluation of Immunostimulatory Activities
2.5.1. Cell Viability Assay
2.5.2. Nitric Oxide Assay
2.5.3. Total RNA Extraction
2.5.4. Synthesis of cDNA and Real-Time Quantitative PCR (RT-qPCR)
2.5.5. Protein Extraction
2.5.6. Western Blotting
2.6. Statistical Analyses
3. Results
3.1. Selection of Lactic Acid Bacteria with Immunostimulatory Function and Identification of the Selected Strain
3.1.1. Selection of Lactic Acid Bacteria with Immunostimulatory Function
3.1.2. Identification of Selected Lactic Acid Bacteria Strain
3.2. Probiotic Characteristics
3.2.1. Acid, Bile, and Pancreatin Tolerances and the Ability to Adhere to Caco-2 Cells
3.2.2. Antioxidant Activities of Lactic Acid Bacteria
3.3. Immunostimulatory Activities of L. lactis subsp. lactis CAB701
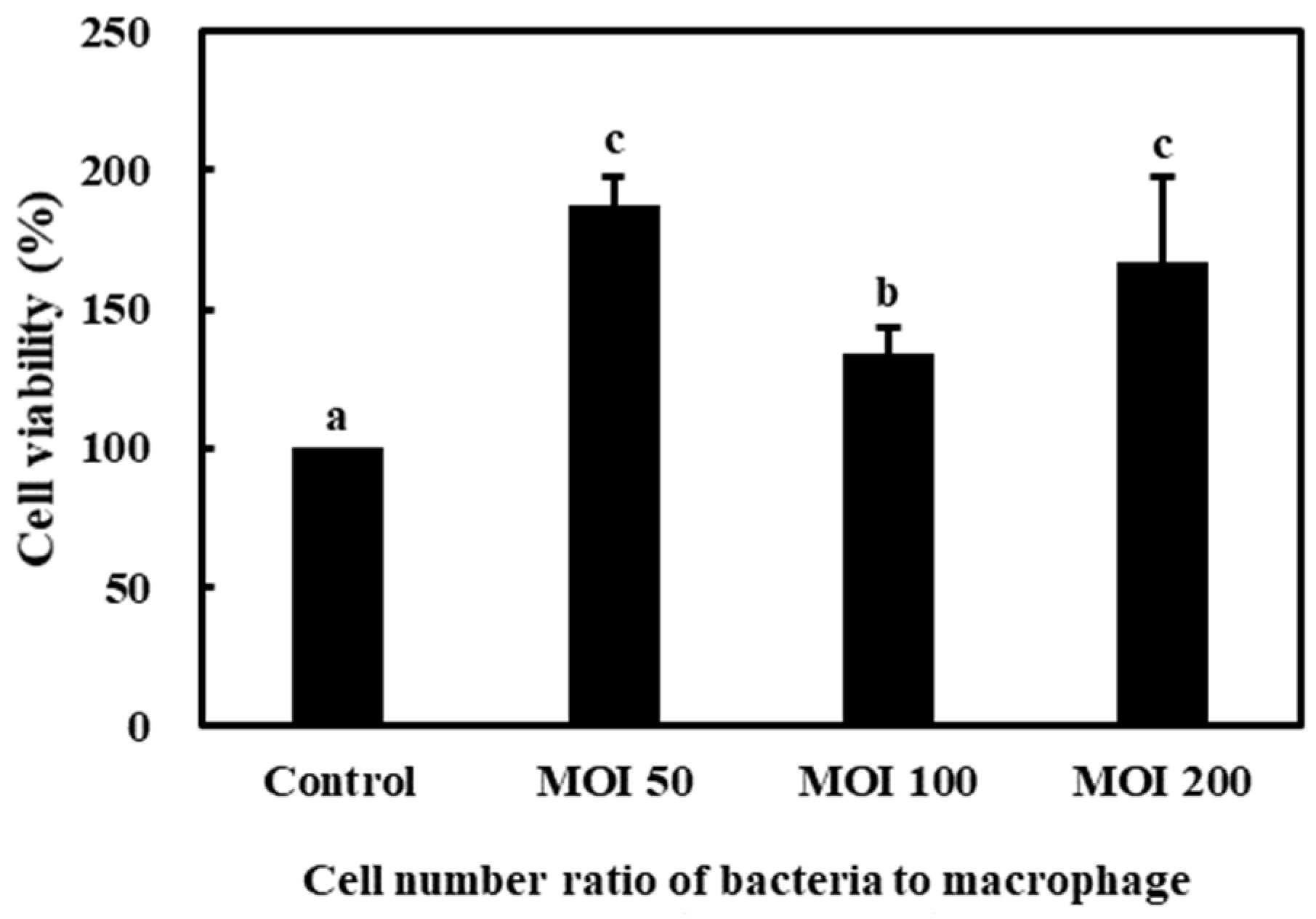
3.3.1. Cell Viability
3.3.2. Production of NO
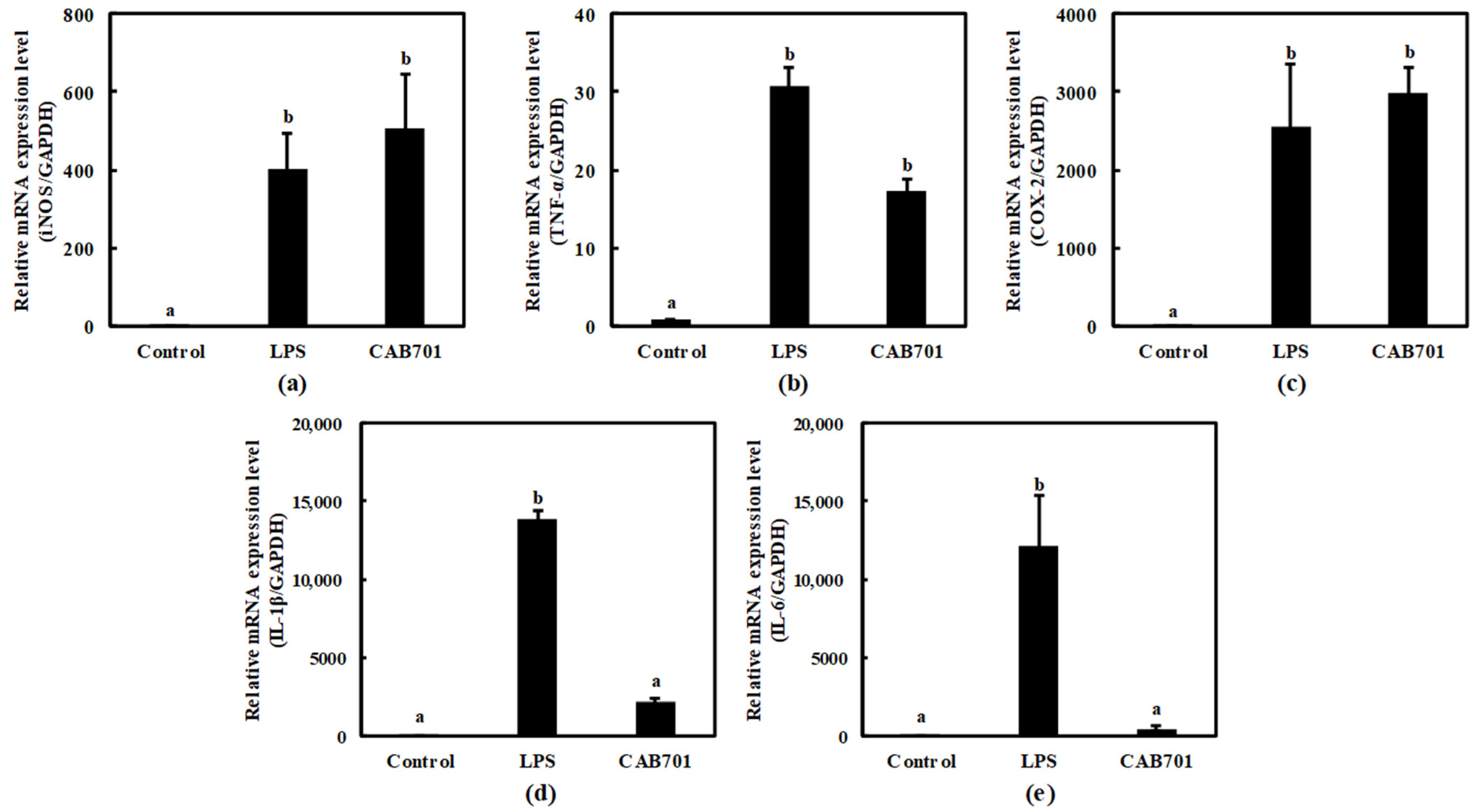
3.3.3. Quantitative Analysis of Cytokine Expression in Macrophages Treated with Lactic Acid Bacteria
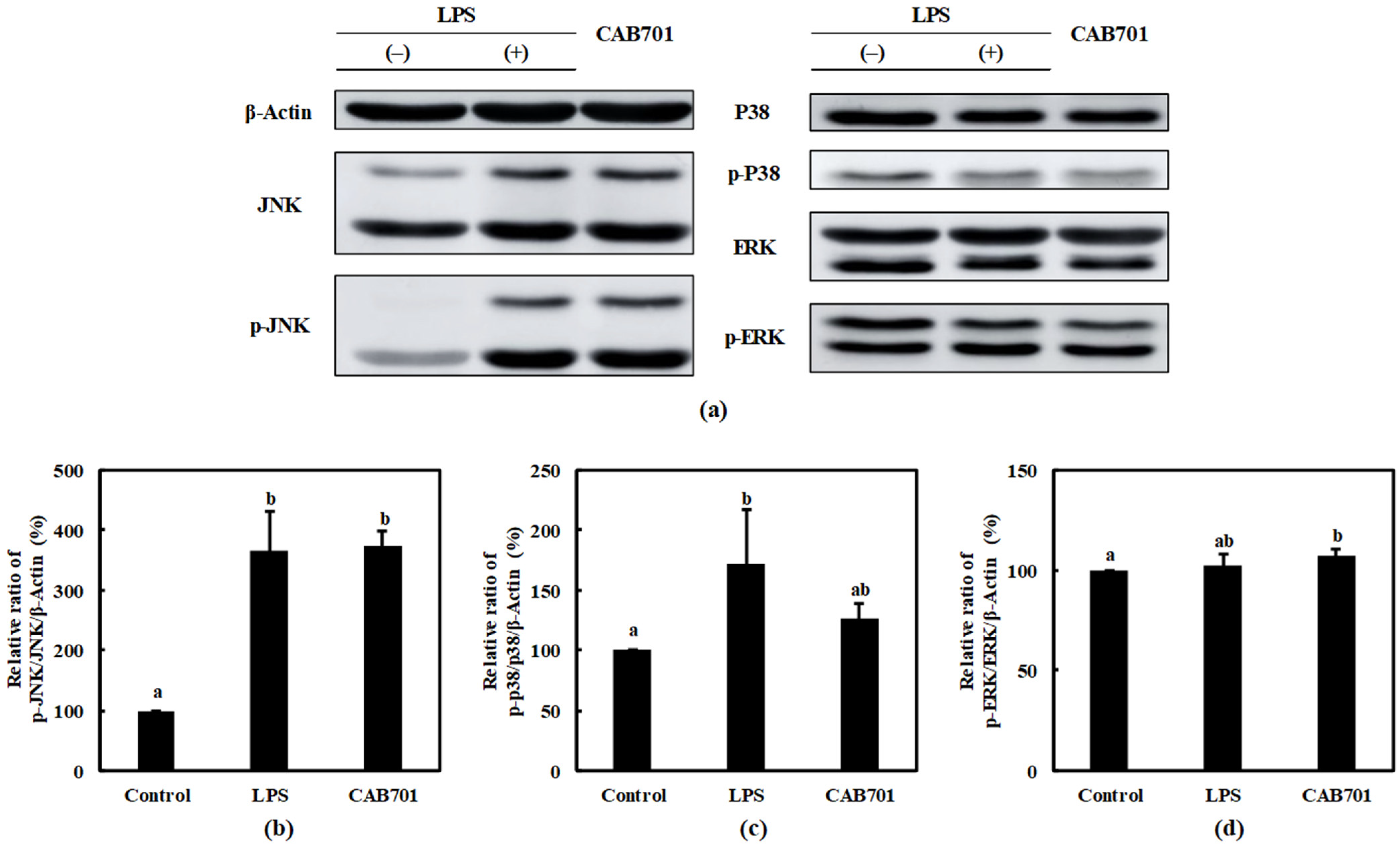
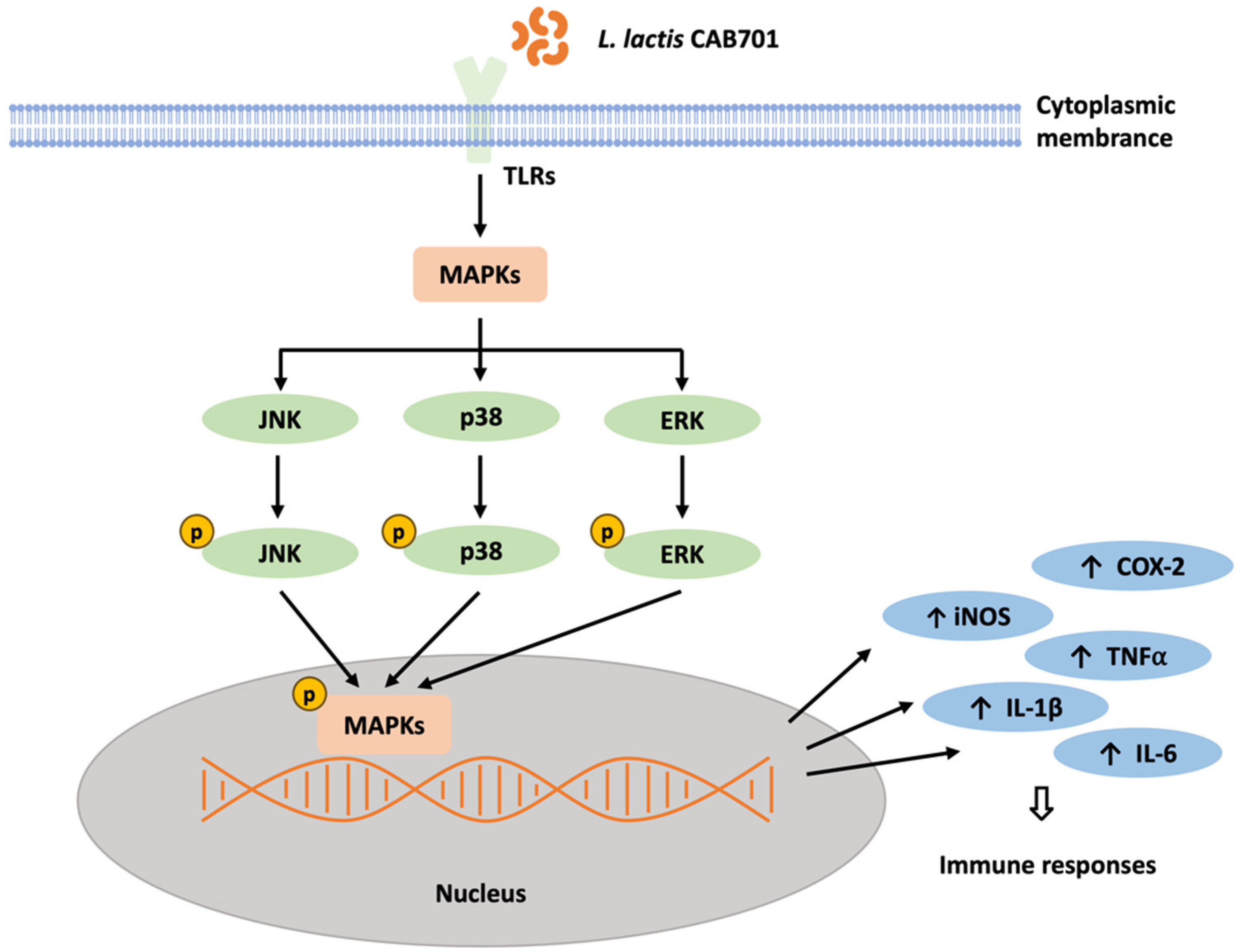
3.3.4. Quantitative Analysis of Mitogen-Activated Protein Kinase (MAPK) Pathway Protein Expression in Macrophages Treated with L. lactis subsp. lactis CAB701
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food Agriculture Organization/World Health Organization Working Group. Guidelines for the Evaluation of Probiotics in Food; Report of a Joint FAO/WHO; FAO/WHO: London, ON, Canada, 2002. [Google Scholar]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Sanders, M.E.; Guarner, F.; Guerrant, R.; Holt, P.R.; Quigley, E.M.; Sartor, R.B.; Sherman, P.M.; Mayer, E.A. An update on the use and investigation of probiotics in health and disease. Gut 2013, 62, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Buffie, C.G.; Pamer, E.G. Microbiota-mediated colonization resistance against intestinal pathogens. Nat. Rev. Immunol. 2013, 13, 790–801. [Google Scholar] [CrossRef] [PubMed]
- Zmora, N.; Zilberman-Schapira, G.; Suez, J.; Mor, U.; Dori-Bachash, M.; Bashiardes, S.; Kotler, E.; Zur, M.; Regev-Lehavi, D.; Brik, R.B.; et al. Personalized gut mucosal colonization resistance to empiric probiotics is associated with unique host and microbiome features. Cell 2018, 174, 1388–1405.e1321. [Google Scholar] [CrossRef]
- Dinan, T.G.; Cryan, J.F. The microbiome-gut-brain axis in health and disease. Gastroenterol. Clin. North Am. 2017, 46, 77–89. [Google Scholar] [CrossRef]
- Sanchez, M.; Darimont, C.; Drapeau, V.; Emady-Azar, S.; Lepage, M.; Rezzonico, E.; Ngom-Bru, C.; Berger, B.; Philippe, L.; Ammon-Zuffrey, C.; et al. Effect of Lactobacillus rhamnosus CGMCC1.3724 supplementation on weight loss and maintenance in obese men and women. Br. J. Nutr. 2014, 111, 1507–1519. [Google Scholar] [CrossRef]
- Kabat, A.M.; Srinivasan, N.; Maloy, K.J. Modulation of immune development and function by intestinal microbiota. Trends Immunol. 2014, 35, 507–517. [Google Scholar] [CrossRef]
- Song, A.A.; In, L.L.A.; Lim, S.H.E.; Rahim, R.A. A review on Lactococcus lactis: From food to factory. Microb. Cell Fact. 2017, 16, 55. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Han, H.T.; Kim, T.R.; Sohn, M.; Park, Y.S. Immunostimulatory activity of Lactococcus lactis LM1185 isolated from Hydrangea macrophylla. Food Sci. Biotechnol. 2023, 32, 497–506. [Google Scholar] [CrossRef]
- Jin, S.W.; Lee, G.H.; Jang, M.J.; Hong, G.E.; Kim, J.Y.; Park, G.D.; Jin, H.; Kim, H.S.; Choi, J.H.; Choi, C.Y.; et al. Immunomodulatory activity of Lactococcus lactis GCWB1176 in cyclophosphamide-induced immunosuppression model. Microorganisms 2020, 8, 1175. [Google Scholar] [CrossRef]
- Beck, B.R.; Kim, D.; Jeon, J.; Lee, S.M.; Kim, H.K.; Kim, O.J.; Lee, J.I.; Suh, B.S.; Do, H.K.; Lee, K.H.; et al. The effects of combined dietary probiotics Lactococcus lactis BFE920 and Lactobacillus plantarum FGL0001 on innate immunity and disease resistance in olive flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2015, 42, 177–183. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Kechagia, M.; Basoulis, D.; Konstantopoulou, S.; Dimitriadi, D.; Gyftopoulou, K.; Skarmoutsou, N.; Fakiri, E.M. Health benefits of probiotics: A review. ISRN Nutr. 2013, 2013, 481651. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Kang, D.K.; Paik, H.D.; Park, Y.S. Beyond probiotics: A narrative review on an era of revolution. Food Sci. Biotechnol. 2023, 32, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, B.M.; Stanton, C.; Fitzgerald, G.F.; Ross, R.P. Survival of probiotic lactobacilli in acidic environments is enhanced in the presence of metabolizable sugars. Appl. Environ. Microbiol. 2005, 71, 3060–3067. [Google Scholar] [CrossRef]
- Liong, M.T.; Shah, N.P. Bile salt deconjugation ability, bile salt hydrolase activity and cholesterol co-precipitation ability of lactobacilli strains. Int. Dairy J. 2005, 15, 391–398. [Google Scholar] [CrossRef]
- Charteris, W.P.; Kelly, P.M.; Morelli, L.; Collins, J.K. Development and application of an in vitro methodology to determine the transit tolerance of potentially probiotic Lactobacillus and Bifidobacterium species in the upper human gastrointestinal tract. J. Appl. Microbiol. 1998, 84, 759–768. [Google Scholar] [CrossRef]
- Duary, R.K.; Rajput, Y.S.; Batish, V.K.; Grover, S. Assessing the adhesion of putative indigenous probiotic lactobacilli to human colonic epithelial cells. Indian J. Med. Res. 2011, 134, 664–671. [Google Scholar]
- Servin, A.L. Antagonistic activities of lactobacilli and bifidobacteria against microbial pathogens. FEMS Microbiol. Rev. 2004, 28, 405–440. [Google Scholar] [CrossRef]
- Gareau, M.G.; Sherman, P.M.; Walker, W.A. Probiotics and the gut microbiota in intestinal health and disease. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Bron, P.A.; van Baarlen, P.; Kleerebezem, M. Emerging molecular insights into the interaction between probiotics and the host intestinal mucosa. Nat. Rev. Microbiol. 2011, 10, 66–78. [Google Scholar] [CrossRef] [PubMed]
- Seth, A.; Yan, F.; Polk, D.B.; Rao, R.K. Probiotics ameliorate the hydrogen peroxide-induced epithelial barrier disruption by a PKC- and MAP kinase-dependent mechanism. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G1060–G1069. [Google Scholar] [CrossRef]
- Lebeer, S.; Vanderleyden, J.; De Keersmaecker, S.C. Host interactions of probiotic bacterial surface molecules: Comparison with commensals and pathogens. Nat. Rev. Microbiol. 2010, 8, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Mortezaei, F.; Royan, M.; Allaf Noveirian, H.; Babakhani, A.; Alaie Kordghashlaghi, H.; Balcazar, J.L. In vitro assessment of potential probiotic characteristics of indigenous Lactococcus lactis and Weissella oryzae isolates from rainbow trout (Oncorhynchus mykiss Walbaum). J. Appl. Microbiol. 2020, 129, 1004–1019. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Oxidative stress: Oxidants and antioxidants. Exp. Physiol. 1997, 82, 291–295. [Google Scholar] [CrossRef]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free radicals, antioxidants in disease and health. Int. J. Biomed. Sci. 2008, 4, 89–96. [Google Scholar]
- Hoffmann, A.; Kleniewska, P.; Pawliczak, R. Antioxidative activity of probiotics. Arch. Med. Sci. 2021, 17, 792–804. [Google Scholar] [CrossRef]
- Krishnamoorthi, R.; Srinivash, M.; Mahalingam, P.U.; Malaikozhundan, B.; Suganya, P.; Gurushankar, K. Antimicrobial, anti-biofilm, antioxidant and cytotoxic effects of bacteriocin by Lactococcus lactis strain CH3 isolated from fermented dairy products—An in vitro and in silico approach. Int. J. Biol. Macromol. 2022, 220, 291–306. [Google Scholar] [CrossRef] [PubMed]
- Sylvere, N.; Mustopa, A.Z.; Budiarti, S.; Meilina, L.; Hertati, A.; Handayani, I. Whole-genome sequence analysis and probiotic characteristics of Lactococcus lactis subsp. lactis strain Lac3 isolated from traditional fermented buffalo milk (Dadih). J. Genet. Eng. Biotechnol. 2023, 21, 49. [Google Scholar] [CrossRef] [PubMed]
- Douillard, F.P.; de Vos, W.M. Functional genomics of lactic acid bacteria: From food to health. Microb. Cell Fact. 2014, 13 (Suppl. S1), S8. [Google Scholar] [CrossRef] [PubMed]
- Farinha, L.; Sabo, S.S.; Porto, M.C.; Souza, E.C.; Oliveira, M.N.; Oliveira, R.P. Influence of prebiotic ingredients on the growth kinetics and bacteriocin production of Lactococcus lactis. Chem. Eng. Trans. 2015, 43, 33–318. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences | Size (bp) | |
|---|---|---|---|
| GAPDH | F | 5′- CAT GGC CTT CCG TGT TCC TAC -3′ | 122 |
| R | 5′- TCA GTG GGC CCT CAG ATG C -3′ | ||
| COX-2 | F | 5′- CTC AGC CAT ACA GCA AAT CCT T -3′ | 101 |
| R | 5′- GTC CGG GTA CAA TCG CAC TTA T -3′ | ||
| iNOS | F | 5′- CCA GCC TGC CCC TTC AAT -3′ | 104 |
| R | 5′- ATC CTT CGG CCC ACT TCC T -3′ | ||
| IL-1β | F | 5′- TGA CGG ACC CCA AAA GAT -3′ | 122 |
| R | 5′- GTG ATA CTG CCT GCC TGA AG -3′ | ||
| IL-6 | F | 5′- CCG GAG AGG AGA CTT CAC AGA G -3′ | 107 |
| R | 5′- TCA TTT CCA CGA TTT CCC AGA G -3′ | ||
| TNF-α | F | 5′- AGG CAC TCC CCC AAA AGA TG -3′ | 122 |
| R | 5′- CAC CCC GAA GTT CAG TAG ACA GA -3′ | ||
| Ingredients | L. lactis subsp. lactis NCTC 6681 * | L. lactis subsp. lactis CAB701 | Ingredients | L. lactis subsp. lactis NCTC 6681 | L. lactis subsp. lactis CAB701 |
|---|---|---|---|---|---|
| Glycerol | − ** | − | Esculin ferric citrate | + | + |
| Erythritol | − | − | Salicin | + | + |
| d-Arabinose | − | − | d-Cellobiose | + | + |
| l-Arabinose | − | − | d-Maltose | + | + |
| d-Ribose | + | + | d-Lactose (bovine origin) | + | + |
| d-Xylose | + | + | d-Melibiose | − | − |
| l-Xylose | + | + | d-Saccharose (sucrose) | + | + |
| d-Adonitol | − | − | d-Trehalose | + | + |
| Methyl-β-d- xylopyranoside | − | − | Inulin | − | + |
| d-Galactose | − | − | d-Melezitose | − | − |
| d-Glucose | + | + | d-Raffinose | − | − |
| d-Fructose | + | + | Amidon (starch) | + | + |
| d-Mannose | + | + | Glycogen | − | − |
| l-Sorbose | + | + | Xylitol | − | − |
| l-Rhanmose | − | − | Gentiobiose | + | + |
| Dulcitol | − | − | d-Turanose | − | − |
| Inositol | − | − | d-Lyxose | − | − |
| d-Mannitol | − | − | d-Tagatose | − | − |
| d-Sorbitol | + | + | d-Fucose | − | − |
| Methyl-α-d- mannopyranoside | − | − | l-Fucose | − | − |
| Methyl-α-d- glucopyranoside | − | − | d-Arabitol | − | − |
| N-Acetyl glucosamine | − | − | l-Arabitol | − | − |
| Amygdalin | + | + | Potassium Gluconate | + | + |
| Arbutin | + | + | Potassium 2- ketogluconate | − | − |
| Esculin ferric citrate | + | + | Potassium 5- ketogluconate | − | − |
| Strain | Acid-Tolerance (%) | Bile-Tolerance (%) | Pancreatin- Tolerance (%) | Adhesion Ability (%) |
|---|---|---|---|---|
| Lactococcus lactis subsp. lactis CAB701 | * 47.6 ± 2.0 | * 63.6 ± 9.3 | * 101.9 ± 0.3 | 75.2 ± 5.4 |
| Lacticaseibacillus rhamnosus GG | 83.3 ± 2.3 | 88.2 ± 0.8 | 104.1 ± 0.3 | 77.2 ± 1.4 |
| Strain | Antioxidant Activity (%) |
|---|---|
| Lactococcus lactis subsp. lactis CAB701 | 95.6 ± 1.3 |
| Lacticaseibacillus rhamnosus GG | 97.2 ± 0.6 |
| Strain | NO Production (%) |
|---|---|
| Lactococcus lactis subsp. lactis CAB701 | * 86.3 ± 6.8 |
| Lacticaseibacillus rhamnosus GG | 18.5 ± 3.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, H.; Kim, S.; Hwang, U.-S.; Choi, H.; Park, Y.-S. Immunostimulatory Activity of Lactococcus lactis subsp. lactis CAB701 Isolated from Jeju Cabbage. Microorganisms 2023, 11, 1718. https://doi.org/10.3390/microorganisms11071718
Jeong H, Kim S, Hwang U-S, Choi H, Park Y-S. Immunostimulatory Activity of Lactococcus lactis subsp. lactis CAB701 Isolated from Jeju Cabbage. Microorganisms. 2023; 11(7):1718. https://doi.org/10.3390/microorganisms11071718
Chicago/Turabian StyleJeong, Huijin, Suin Kim, Un-Sik Hwang, Hyukjoon Choi, and Young-Seo Park. 2023. "Immunostimulatory Activity of Lactococcus lactis subsp. lactis CAB701 Isolated from Jeju Cabbage" Microorganisms 11, no. 7: 1718. https://doi.org/10.3390/microorganisms11071718
APA StyleJeong, H., Kim, S., Hwang, U.-S., Choi, H., & Park, Y.-S. (2023). Immunostimulatory Activity of Lactococcus lactis subsp. lactis CAB701 Isolated from Jeju Cabbage. Microorganisms, 11(7), 1718. https://doi.org/10.3390/microorganisms11071718