
Biocontrol of Pathogen Microorganisms in Ripened Foods of Animal Origin




Abstract
1. Introduction
2. Pathogenic Bacteria in Ripened Foods and Biocontrol Strategies
2.1. Pathogenic Bacteria
2.1.1. Listeria monocytogenes
2.1.2. Salmonella enterica
2.1.3. Staphylococcus aureus
2.1.4. Clostridium botulinum
2.1.5. Escherichia coli
2.2. Microbial Biocontrol Strategies
2.3. Plant-Derivative Biocontrol Agents
3. Pathogenic Yeasts in Ripened Foods and Biocontrol Strategies
3.1. Pathogenic Yeasts

{kind=link}
| Type of Cheese | Yeast | Pathogenic Capacity | Reference |
|---|---|---|---|
| Cottage and cream cheese | Geotrichum species | No | [119] |
| Domiati and Kariesh cheese | Candida albicans Candida lusitaniae (teleomorph of Clavispora lusitaniae) Candida catenulata | Yes | [110] |
| Feta cheese | Candida tropicalis | No | [128] |
| Smeared cheese | Issatchenkia orientalis (teleomorph of Candida krusei) C. lusitaniae (teleomorph of C. lusitaniae) Candida rugosa Candida famata (teleomorph of Debaryomyces hansenii) | Yes | [134] |
| Several types of cheeses | Candida parapsilosis, C. catenulata, Yarrowia lipolytica, Rhodotorula glutinis and Trichosporon species | Yes | [129] |
| Artisanal cheese | C. albicans and 97.6% as Candida non-albicans, distributed in 79.3% of C. krusei, 12.3% of C. glabrata and 6.0% of C. tropicalis | No | [126] |
| Swiss-type blue cheese, Goat’s milk Cheddar | C. parapsilosis | No | [115] |
| Manteca (Italian cheese) | C. parapsilosis | No | [115] |
| Washed rind cheese | Y. lipolytica | No | [115] |
| Camembert Blue-veined cheeses | Candida intermedia | No | [135] |
3.2. Yeast Biocontrol Strategies Using Microorganisms
3.3. Plant-Derivative Biocontrol Strategies
4. Pathogenic and Toxigenic Molds in Ripened Foods and Biocontrol Strategies
4.1. Biocontrol Strategies against Ochratoxin A-Producing Molds
4.2. Biocontrol Strategies against Aflatoxin-Producing Molds
4.3. Biocontrol Strategies against Cyclopiazonic Acid-Producing Molds
4.4. Biocontrol Strategies against Sterigmatocystin-Producing Molds
4.5. Biocontrol Strategies against Citrinin-Producing Molds
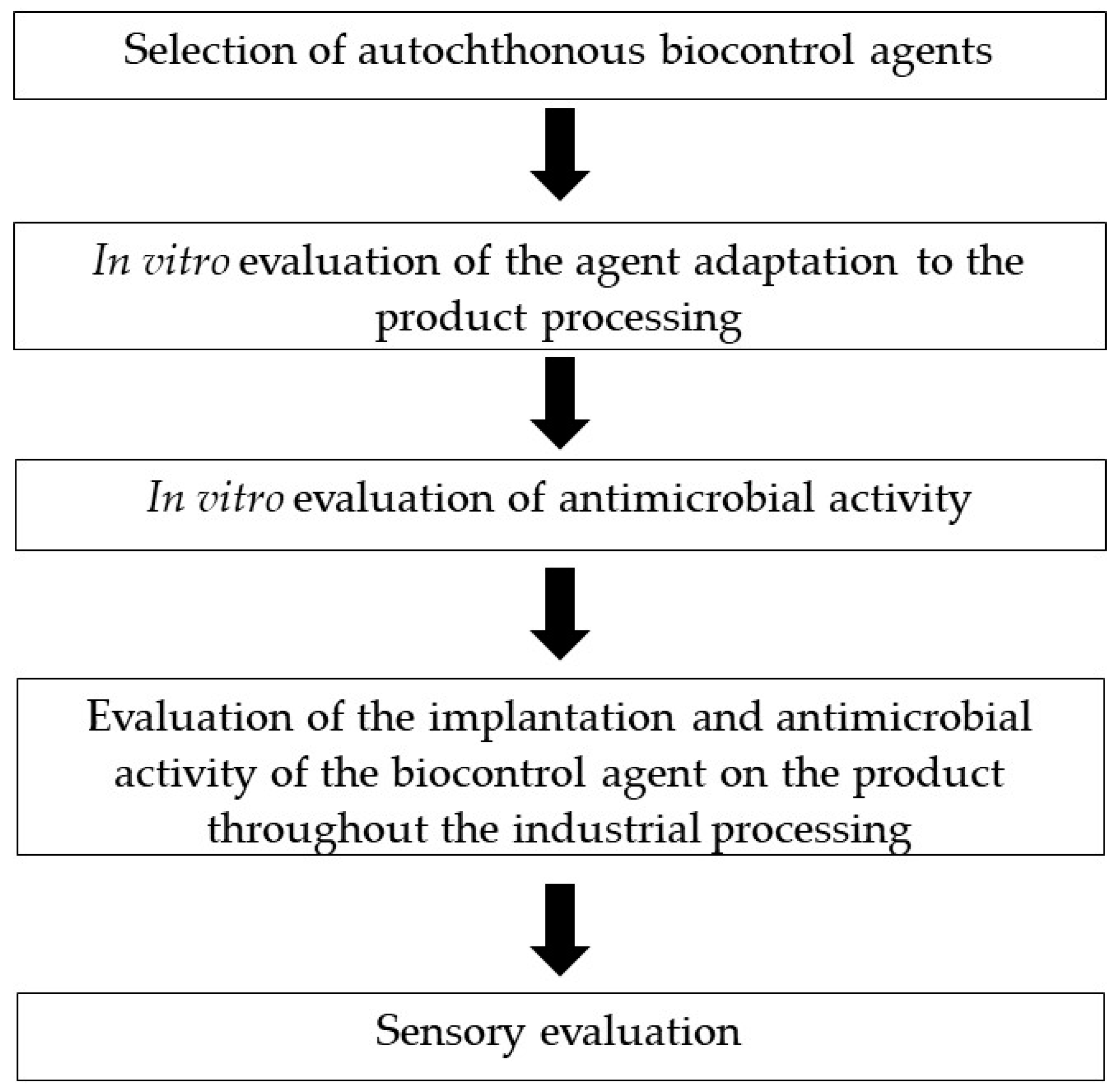
5. Challenges in the Application of Microorganisms as Biocontrol Agents in Animal Origin Ripened Foods
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Andrade, M.J.; Córdoba, J.J.; Casado, E.M.; Córdoba, M.G.; Rodríguez, M. Effect of selected strains of Debaryomyces hansenii on the volatile compound production of dry fermented sausage “salchichón”. Meat Sci. 2010, 85, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Camargo, A.C.; de Araújo, J.P.A.; Fusieger, A.; de Carvalho, A.F.; Nero, L.A. Microbiological quality and safety of Brazilian artisanal cheeses. Braz. J. Microbiol. 2021, 52, 393–409. [Google Scholar] [CrossRef] [PubMed]
- Cocolin, L.; Urso, R.; Rantsiou, K.; Cantoni, C.; Comi, G. Dynamics and characterization of yeasts during natural fermentation of Italian sausages. FEMS Yeast Res. 2006, 6, 692–701. [Google Scholar] [CrossRef] [PubMed]
- Martín, A.; Córdoba, J.J.; Aranda, E.; Córdoba, M.G.; Asensio, M.A. Contribution of a selected fungal population to the volatile compounds on dry-cured ham. Int. J. Food Microbiol. 2006, 110, 8–18. [Google Scholar] [CrossRef]
- Martín, B.; Garriga, M.; Hugas, M.; Bover-Cid, S.; Veciana-Nogués, M.T.; Aymerich, T. Molecular, technological and safety characterization of Gram-positive catalase-positive cocci from slightly fermented sausages. Int. J. Food Microbiol. 2006, 107, 148–158. [Google Scholar] [CrossRef]
- Prpich, N.Z.P.; Camprubí, G.E.; Cayré, M.E.; Castro, M.P. Indigenous microbiota to leverage traditional dry sausage production. Int. J. Food Sci. 2021, 2015, 6696856. [Google Scholar]
- Benito, M.J.; Martín, A.; Aranda, E.; Pérez-Nevado, F.; Ruiz-Moyano, S.; Córdoba, M.G. Characterization and selection of autochthonous lactic acid bacteria isolated from traditional Iberian dry-fermented salchichón and chorizo sausages. J. Food Sci. 2007, 72, 193–201. [Google Scholar] [CrossRef]
- Casquete, R.; Benito, M.J.; Martín, A.; Ruiz-Moyano, S.; Aranda, E.; Córdoba, M.G. Microbiological quality of salchichón and chorizo, traditional Iberian dry-fermented sausages from two different industries, inoculated with autochthonous starter cultures. Food Control 2012, 24, 191–198. [Google Scholar] [CrossRef]
- Yoon, Y.; Lee, S.; Choi, K.H. Microbial benefits and risks of raw milk cheese. Food Control 2016, 63, 201–215. [Google Scholar] [CrossRef]
- Hospital, X.F.; Hierro, E.; Stringer, S.; Fernández, M. A study on the toxigenesis by Clostridium botulinum in nitrate and nitrite-reduced dry fermented sausages. Int. J. Food Microbiol. 2016, 218, 66–70. [Google Scholar] [CrossRef]
- Chaidoutis, E.; Keramydas, D.; Papalexis, P.; Migdanis, A.; Migdanis, I.; Lazaris, A.C.; Kavantzas, N. Foodborne botulism: A brief review of cases transmitted by cheese products. Biomed. Rep. 2022, 16, 41. [Google Scholar] [CrossRef] [PubMed]
- Falardeau, J.; Trmčić, A.; Wang, S. The occurrence, growth, and biocontrol of Listeria monocytogenes in fresh and surface-ripened soft and semisoft cheeses. Compr. Rev. Food Sci. Food Saf. 2021, 20, 4019–4048. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, V.; Barbosa, J.; Silva, J.; Felício, M.T.; Mena, C.; Hogg, T.; Gibbs, P.; Teixeira, P. Characterisation of alheiras, traditional sausages produced in the North of Portugal, with respect to their microbiological safety. Food Control 2007, 18, 436–440. [Google Scholar] [CrossRef]
- Gonçalves, M.T.P.; Benito, M.J.; Córdoba, M.d.G.; Egas, C.; Merchán, A.V.; Galván, A.I.; Ruiz-Moyano, S. Bacterial communities in Serpa cheese by culture dependent techniques, 16S rRNA gene sequencing and high-throughput sequencing analysis. J. Food Sci. 2018, 83, 1333–1341. [Google Scholar] [CrossRef] [PubMed]
- Koutsoumanis, K.; Allende, A.; Álvarez-Ordóñez, A.; Bover-Cid, S.; Chemaly, M.; De Cesare, A.; Herman, L.; Hilbert, F.; Lindqvist, R.; Nauta, M.; et al. Microbiological safety of aged meat. EFSA J. 2023, 21, 7745. [Google Scholar]
- Lobacz, A.; Zulewska, J. Fate of Salmonella spp. in the fresh soft raw milk cheese during storage at different temperatures. Microorganisms 2021, 9, 938. [Google Scholar] [CrossRef]
- Meloni, D. Presence of Listeria monocytogenes in Mediterranean-style dry fermented sausages. Foods 2015, 4, 34–50. [Google Scholar] [CrossRef]
- Possas, A.; Bonilla-Luque, O.M.; Valero, A. From cheese-making to consumption: Exploring the microbial safety of cheeses through predictive microbiology models. Foods 2021, 10, 355. [Google Scholar] [CrossRef]
- Ertas Onmaz, N.; Gungor, C.; Al, S.; Dishan, A.; Hizlisoy, H.; Yildirim, Y.; Kasap Tekinsen, F.; Disli, H.B.; Barel, M.; Karadal, F. Mycotoxigenic and phylogenetic perspective to the yeasts and filamentous moulds in mould-matured Turkish cheese. Int. J. Food Microbiol. 2021, 357, 109385. [Google Scholar] [CrossRef]
- Battilani, P.; Pietri, A.; Giorni, P.; Formenti, S.; Bertuzzi, T.; Toscani, T.; Virgili, R.; Kozakiewicz, Z. Penicillium populations in dry-cured ham manufacturing plants. J. Food. Prot. 2007, 70, 975–980. [Google Scholar] [CrossRef]
- López-Díaz, T.M.; Santos, J.A.; García-López, M.L.; Otero, A. Surface mycoflora of a Spanish fermented meat sausage and toxigenicity of Penicillium isolates. Int. J. Food Microbiol. 2001, 68, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Martı́n, A.; Córdoba, J.J.; Núñez, F.; Benito, M.J.; Asensio, M.A. Contribution of a selected fungal population to proteolysis on dry-cured ham. Int. J. Food Microbiol. 2004, 94, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Dobson, A.D.W. Mycotoxins in Cheese. In Cheese. Chemistry, Physics & Microbiology; McSweeney, P.L.H., Fox, P.F., Cotter, P.D., Everett, D.W., Eds.; Academic Press: Cambridge, MA, USA; Elsevier Ltd.: Amsterdam, The Netherlands, 2017; Volume 1, ISBN 9780122636530. [Google Scholar]
- Lešić, T.; Vulić, A.; Vahčić, N.; Šarkanj, B.; Hengl, B.; Kos, I.; Polak, T.; Kudumija, N.; Pleadin, J. The Occurrence of five unregulated mycotoxins most important for traditional dry-cured meat products. Toxins 2022, 14, 476. [Google Scholar] [CrossRef] [PubMed]
- Markov, K.; Pleadin, J.; Bevardi, M.; Vahčić, N.; Sokolić-Mihalak, D.; Frece, J. Natural occurrence of aflatoxin B1, ochratoxin A and citrinin in Croatian fermented meat products. Food Control 2013, 34, 312–317. [Google Scholar] [CrossRef]
- European Commission Rapid Alert System for Food and Feed (RASFF). Available online: https://webgate.ec.europa.eu/rasff-window/screen/search (accessed on 26 March 2023).
- Asensio, M.A.; Núñez, F.; Delgado, J.; Bermúdez, E. Control of toxigenic molds in food processing. In Microbial Food Safety and Preservation Techniques; Rai, V.R., Bai, A.J., Eds.; CRC Press: Boca Raton, FL, USA, 2014; pp. 329–357. [Google Scholar]
- Alcano, M.J.; Jahn, R.C.; Scherer, C.D.; Wigmann, É.F.; Moraes, V.M.; Garcia, M.V.; Mallmann, C.A.; Copetti, M.V. Susceptibility of Aspergillus spp. to acetic and sorbic acids based on pH and effect of sub-inhibitory doses of sorbic acid on ochratoxin A production. Food Res. Int. 2016, 81, 25–30. [Google Scholar] [CrossRef]
- Román, S.; Sánchez-Siles, L.; Siegrist, M. The importance of food naturalness for consumers: Results of a systematic review. Trends Food Sci. Technol. 2017, 67, 44–57. [Google Scholar] [CrossRef]
- Álvarez, M.; Andrade, M.J.; García, C.; Rondán, J.J.; Núñez, F. Effects of preservative agents on quality attributes of dry-cured fermented sausages. Foods 2020, 9, 1505. [Google Scholar] [CrossRef] [PubMed]
- Cebrián, E.; Núñez, F.; Álvarez, M.; Roncero, E.; Rodríguez, M. Biocontrol of ochratoxigenic Penicillium nordicum in dry-cured fermented sausages by Debaryomyces hansenii and Staphylococcus xylosus. Int. J. Food Microbiol. 2022, 375, 109744. [Google Scholar] [CrossRef] [PubMed]
- Cebrián, E.; Rodríguez, M.; Peromingo, B.; Bermúdez, E.; Núñez, F. Efficacy of the combined protective cultures of Penicillium chrysogenum and Debaryomyces hansenii for the control of ochratoxin A hazard in dry-cured ham. Toxins 2019, 11, 710. [Google Scholar] [CrossRef]
- Alía, A.; Córdoba, J.J.; Rodríguez, A.; García, C.; Andrade, M.J. Evaluation of the efficacy of Debaryomyces hansenii as protective culture for controlling Listeria monocytogenes in sliced dry-cured ham. LWT—Food Sci. Technol. 2020, 119, 108886. [Google Scholar] [CrossRef]
- Delgado, J.; Peromingo, B.; Rodríguez, A.; Rodríguez, M. Biocontrol of Penicillium griseofulvum to reduce cyclopiazonic acid contamination in dry-fermented sausages. Int. J. Food Microbiol. 2019, 293, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ho, V.T.T.; Lo, R.; Bansal, N.; Turner, M.S. Characterisation of Lactococcus lactis isolates from herbs, fruits and vegetables for use as biopreservatives against Listeria monocytogenes in cheese. Food Control 2018, 85, 472–483. [Google Scholar] [CrossRef]
- Campagnollo, F.B.; Pedrosa, G.T.S.; Kamimura, B.A.; Furtado, M.M.; Baptista, R.C.; Nascimento, H.M.; Alvarenga, V.O.; Magnani, M.; Sant’Ana, A.S. Growth potential of three strains of Listeria monocytogenes and Salmonella enterica in Frescal and semi-hard artisanal Minas microcheeses: Impact of the addition of lactic acid bacteria with antimicrobial activity. LWT—Food Sci. Technol. 2022, 158, 113169. [Google Scholar] [CrossRef]
- Iacumin, L.; Manzano, M.; Andyanto, D.; Comi, G. Biocontrol of ochratoxigenic moulds (Aspergillus ochraceus and Penicillium nordicum) by Debaryomyces hansenii and Saccharomycopsis fibuligera during speck production. Food Microbiol. 2017, 62, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Martín, I.; Alía, A.; Rodríguez, A.; Gómez, F.; Córdoba, J.J. Growth and expression of virulence genes of Listeria monocytogenes during the processing of dry-cured fermented “salchichón” manufactured with a selected Lactilactobacillus sakei. Biology 2021, 10, 1258. [Google Scholar] [CrossRef]
- Peromingo, B.; Andrade, M.J.; Delgado, J.; Sánchez-Montero, L.; Núñez, F. Biocontrol of aflatoxigenic Aspergillus parasiticus by native Debaryomyces hansenii in dry-cured meat products. Food Microbiol. 2019, 82, 269–276. [Google Scholar] [CrossRef]
- Álvarez, M.; Andrade, M.J.; Delgado, J.; Núñez, F.; Román, Á.C.; Rodrigues, P. Rosmarinus officinalis reduces the ochratoxin A production by Aspergillus westerdijkiae in a dry-cured fermented sausage-based medium. Food Control 2023, 145, 109436. [Google Scholar] [CrossRef]
- Álvarez, M.; Rodríguez, A.; Núñez, F.; Silva, A.; Andrade, M.J. In vitro antifungal effects of spices on ochratoxin A production and related gene expression in Penicillium nordicum on a dry-cured fermented sausage medium. Food Control 2020, 114, 107222. [Google Scholar] [CrossRef]
- García-Díez, J.; Alheiro, J.; Pinto, A.L.; Soares, L.; Falco, V.; Fraqueza, M.J.; Patarata, L. Behaviour of food-borne pathogens on dry cured sausage manufactured with herbs and spices essential oils and their sensorial acceptability. Food Control 2016, 59, 262–270. [Google Scholar] [CrossRef]
- Hlebová, M.; Foltinová, D.; Vešelényiová, D.; Medo, J.; Šramková, Z.; Tančinová, D.; Mrkvová, M.; Hleba, L. The vapor phase of selected essential oils and their antifungal activity in vitro and in situ against Penicillium commune, a common contaminant of cheese. Foods 2022, 11, 3517. [Google Scholar] [CrossRef]
- Sánchez-Montero, L.; Córdoba, J.J.; Alía, A.; Peromingo, B.; Núñez, F. Effect of Spanish smoked paprika “Pimentón de La Vera” on control of ochratoxin A and aflatoxins production on a dry-cured meat model system. Int. J. Food Microbiol. 2019, 308, 108303. [Google Scholar] [CrossRef] [PubMed]
- Tatlisu, N.B.; Yilmaz, M.T.; Arici, M. Fabrication and characterization of thymol-loaded nanofiber mats as a novel antimould surface material for coating cheese surface. Food Packag. Shelf Life 2019, 21, 100347. [Google Scholar] [CrossRef]
- Abebe, E.; Gugsa, G.; Ahmed, M. Review on major food-borne zoonotic bacterial pathogens. J. Trop Med. 2020, 2020, 4674235. [Google Scholar] [CrossRef] [PubMed]
- Schlech, W.F. Epidemiology and clinical manifestations of Listeria monocytogenes infection. Microbiol. Spectr. 2019, 7, 793–802. [Google Scholar] [CrossRef]
- Mateus, T.; Silva, J.; Maia, R.L.; Teixeira, P. Listeriosis during pregnancy: A Public Health Concern. ISRN Obstet. Gynecol. 2013, 2013, 851712. [Google Scholar] [CrossRef]
- Chen, J.; Luo, X.; Jiang, L.; Jin, P.; Wei, W.; Liu, D.; Fang, W. Molecular characteristics and virulence potential of Listeria monocytogenes isolates from Chinese food systems. Food Microbiol. 2009, 26, 103–111. [Google Scholar] [CrossRef]
- Aljasir, S.F.; D’Amico, D.J. The effect of protective cultures on Staphylococcus aureus growth and enterotoxin production. Food Microbiol. 2020, 91, 103541. [Google Scholar] [CrossRef]
- Le, H.H.T.; Dalsgaard, A.; Andersen, P.S.; Nguyen, H.M.; Ta, Y.T.; Nguyen, T.T. Large-scale Staphylococcus aureus foodborne disease poisoning outbreak among primary school children. Microbiol. Res. 2021, 12, 43–52. [Google Scholar] [CrossRef]
- Pal, M. Staphylococcus aureus: A Major pathogen of food poisoning. Nutr. Food Process 2022, 5, 1–3. [Google Scholar] [CrossRef]
- Hamzah, A.M.C.; Yeo, C.C.; Puah, S.M.; Chua, K.H.; Chew, C.H. Staphylococcus aureus infections in Malaysia: A review of antimicrobial resistance and characteristics of the clinical isolates, 1990–2017. Antibiotics 2019, 8, 1990–2017. [Google Scholar] [CrossRef]
- Irlinger, F. Safety assessment of dairy microorganisms: Coagulase-negative staphylococci. Int. J. Food Microbiol. 2008, 126, 302–310. [Google Scholar] [CrossRef]
- Soares, J.C.; Marques, M.R.; Tavaria, F.K.; Pereira, J.O.; Malcata, F.X.; Pintado, M.M. Biodiversity and characterization of Staphylococcus species isolated from a small manufacturing dairy plant in Portugal. Int. J. Food Microbiol. 2011, 146, 123–129. [Google Scholar] [CrossRef]
- Cheung, G.Y.C.; Bae, J.S.; Otto, M. Pathogenicity and virulence of Staphylococcus aureus. Virulence 2021, 12, 547–569. [Google Scholar] [CrossRef]
- Doyle, M. Foodborne Bacterial Pathogens (Food Science and Technology), 1st ed.; CRC Press: Boca Raton, FL, USA, 1989; ISBN 978-0824778668. [Google Scholar]
- Alizadeh, A.M.; Hashempour-Baltork, F.; Alizadeh-Sani, M.; Maleki, M.; Azizi-Lalabadi, M.; Khosravi-Darani, K. Inhibition of Clostridium botulinum and its toxins by probiotic bacteria and their metabolites: An update review. Qual. Assur. Saf. Crop. Foods 2020, 12, 59–68. [Google Scholar] [CrossRef]
- Hamad, G.; Ombarak, R.A.; Eskander, M.; Mehany, T.; Anees, F.R.; Elfayoumy, R.A.; Omar, S.A.; Lorenzo, J.M.; Abou-Alella, S.A.E. Detection and inhibition of Clostridium botulinum in some Egyptian fish products by probiotics cell-free supernatants as bio-preservation agents. LWT—Food Sci. Technol. 2022, 163, 113603. [Google Scholar] [CrossRef]
- Shah, M.K. A review on pathogenic Escherichia coli in Malaysia. Adv. Anim. Vet. Sci. 2018, 6, 95–107. [Google Scholar] [CrossRef]
- Yang, S.C.; Lin, C.H.; Aljuffali, I.A.; Fang, J.Y. Current pathogenic Escherichia coli foodborne outbreak cases and therapy development. Arch. Microbiol. 2017, 199, 811–825. [Google Scholar] [CrossRef] [PubMed]
- Mersha, G.; Asrat, D.; Zewde, B.M.; Kyule, M. Occurrence of Escherichia coli O157:H7 in faeces, skin and carcasses from sheep and goats in Ethiopia. Lett. Appl. Microbiol. 2010, 50, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Castellano, P.; Ibarreche, M.P.; Massani, M.B.; Fontana, C.; Vignolo, G.M. Strategies for pathogen biocontrol using lactic acid bacteria and their metabolites: A focus on meat ecosystems and industrial environments. Microorganisms 2017, 5, 38. [Google Scholar] [CrossRef]
- Abdelhamid, A.G.; El-Dougdoug, N.K. Controlling foodborne pathogens with natural antimicrobials by biological control and antivirulence strategies. Heliyon 2020, 6, e05020. [Google Scholar] [CrossRef]
- Martín, I.; Rodríguez, A.; Sánchez-Montero, L.; Padilla, P.; Córdoba, J.J. Effect of the dry-cured fermented sausage “salchichón” processing with a selected Lactobacillus sakei in Listeria monocytogenes and microbial population. Foods 2021, 10, 856. [Google Scholar] [CrossRef] [PubMed]
- Martín, I.; Rodríguez, A.; Córdoba, J.J. Application of selected lactic-acid bacteria to control Listeria monocytogenes in soft-ripened “Torta del Casar” cheese. LWT—Food Sci. Technol. 2022, 168, 113873. [Google Scholar] [CrossRef]
- Margalho, L.P.; Jorge, G.P.; Noleto, D.A.P.; Silva, C.E.; Abreu, J.S.; Piran, M.V.F.; Brocchi, M.; Sant’Ana, A.S. Biopreservation and probiotic potential of a large set of lactic acid bacteria isolated from Brazilian artisanal cheeses: From screening to in product approach. Microbiol. Res. 2021, 242, 126622. [Google Scholar] [CrossRef] [PubMed]
- Pragalaki, T.; Bloukas, J.G.; Kotzekidou, P. Inhibition of Listeria monocytogenes and Escherichia coli O157: H7 in liquid broth medium and during processing of fermented sausage using autochthonous starter cultures. Meat Sci. 2013, 95, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Kačániová, M.; Borotová, P.; Terentjeva, M.; Kunová, S.; Felšöciová, S.; Haščík, P.; Lopašovský, Ľ.; Štefániková, J. Bryndza cheese of Slovak origin as potential resources of probiotic bacteria. Potravin Slovak J. Food Sci. 2020, 14, 641–646. [Google Scholar] [CrossRef]
- Kaban, G.; Kaya, M. Effect of starter culture on growth of Staphylococcus aureus in sucuk. Food Control 2006, 17, 797–801. [Google Scholar] [CrossRef]
- Di Gioia, D.; Mazzola, G.; Nikodinoska, I.; Aloisio, I.; Langerholc, T.; Rossi, M.; Raimondi, S.; Melero, B.; Rovira, J. Lactic acid bacteria as protective cultures in fermented pork meat to prevent Clostridium spp. growth. Int. J. Food Microbiol. 2016, 235, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Tomović, V.; Šojić, B.; Savanović, J.; Kocić-Tanackov, S.; Pavlić, B.; Jokanović, M.; Dordević, V.; Parunović, N.; Martinović, A.; Vujadinović, D. New formulation towards healthier meat products: Juniperus communis L. essential oil as alternative for sodium nitrite in dry fermented sausages. Foods 2020, 9, 1066. [Google Scholar] [CrossRef]
- Dos Santos, L.R.; Alía, A.; Martin, I.; Gottardo, F.M.; Rodrigues, L.B.; Borges, K.A.; Furian, T.Q.; Córdoba, J.J. Antimicrobial activity of essential oils and natural plant extracts against Listeria monocytogenes in a dry-cured ham-based model. J. Sci. Food Agric. 2022, 102, 1729–1735. [Google Scholar] [CrossRef]
- Rodríguez-Sánchez, S.; Ramos, I.M.; Rodríguez-Pérez, M.; Poveda, J.M.; Seseña, S.; Palop, M.L. Lactic acid bacteria as biocontrol agents to reduce Staphylococcus aureus growth, enterotoxin production and virulence gene expression. LWT—Food Sci. Technol. 2022, 170, 114025. [Google Scholar] [CrossRef]
- EFSA BIOHAZ Panel. Updated list of QPS recommended biological agents for safety risk assessments carried out by EFSA. EFSA J. 2023, 21, e07747. [Google Scholar]
- Carvalho, M.I.P.; Albano, H.C.P.; Teixeira, P.C.M. Influence of oregano essential oil on the inhibition of selected pathogens in “Alheira” during storage. Acta Sci. Pol. Technol. Aliment. 2019, 18, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Albano, H.; Oliveira, M.; Aroso, R.; Cubero, N.; Hogg, T.; Teixeira, P. Antilisterial activity of lactic acid bacteria isolated from “Alheiras” (traditional Portuguese fermented sausages): In situ assays. Meat Sci. 2007, 76, 796–800. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Lozano, J.C.; Reguera-Useros, J.I.; Peláez-Martínez, M.d.C.; Sacristán-Pérez-Minayo, G.; Gutiérrez-Fernández, Á.J.; Hardisson de la Torre, A. The effect of the pediocin PA-1 produced by Pediococcus acidilactici against Listeria monocytogenes and Clostridium perfringens in Spanish dry-fermented sausages and frankfurters. Food Control 2010, 21, 679–685. [Google Scholar] [CrossRef]
- Kasra-Kermanshahi, R.; Mobarak-Qamsari, E. Inhibition effect of lactic acid bacteria against food born pathogen, Listeria monocytogenes. Appl. Food Biotechnol. 2015, 2, 11–19. [Google Scholar]
- Martín, I.; Rodríguez, A.; Delgado, J.; Córdoba, J.J. Strategies for biocontrol of Listeria monocytogenes using lactic acid bacteria and their metabolites in ready-to-eat meat-and dairy-ripened products. Foods 2022, 11, 542. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Commission Regulation (EU) No 1129/2011 of 11 November 2011 amending Annex II to Regulation (EC) No 1333/2008 of the European Parliament and of the Council by establishing a Union list of food additives Text with EEA relevance. Off. J. Eir. Union 2013, 045, 131–307. [Google Scholar]
- Darbandi, A.; Asadi, A.; Ari, M.M.; Ohadi, E.; Talebi, M.; Zadeh, M.H.; Emamie, A.D.; Ghanavati, R.; Kakanj, M. Bacteriocins: Properties and potencial use as antimicrobials. J. Clin. Lab. Anal. 2022, 36, e24093. [Google Scholar] [CrossRef]
- Koohestani, M.; Moradi, M.; Tajik, H.; Badali, A. Effects of cell-free supernatant of Lactobacillus acidophilus LA5 and Lactobacillus casei 431 against planktonic form and biofilm of Staphylococcus aureus. Vet. Res. Forum. 2018, 9, 301–306. [Google Scholar]
- Araújo-Rodrigues, H.; dos Santos, M.T.P.G.; Ruiz-Moyano, S.; Tavaria, F.K.; Martins, A.P.L.; Alvarenga, N.; Pintado, M.E. Technological and protective performance of LAB isolated from Serpa PDO cheese: Towards selection and development of an autochthonous starter culture. LWT—Food Sci. Techno. 2021, 150, 112079. [Google Scholar] [CrossRef]
- Martín, I.; Cordoba, J.J.; Alía, A.; Martínez, R.; Rodríguez, A. Selection and characterization of lactic acid bacteria from traditional ripened foods with activity against Listeria monocytogenes. LWT—Food Sci. Techno. 2022, 163, 113579. [Google Scholar] [CrossRef]
- Martín, I.; García, C.; Rodríguez, A.; Córdoba, J.J. Effect of a selected protective culture of Lactilactobacillus sakei on the evolution of volatile compounds and on the final sensorial characteristics of traditional dry-cured fermented “Salchichón”. Biology 2023, 12, 88. [Google Scholar] [CrossRef] [PubMed]
- Martín, I.; Rodríguez, A.; García, C.; Córdoba, J.J. Evolution of volatile vompounds during ripening and final sensory changes of traditional raw Ewe’s milk cheese “Torta del Casar” maturated with selected protective lactic acid bacteria. Foods 2022, 11, 2658. [Google Scholar] [CrossRef] [PubMed]
- Baka, A.M.; Papavergou, E.J.; Pragalaki, T.; Bloukas, J.G.; Kotzekidou, P. Effect of selected autochthonous starter cultures on processing and quality characteristics of Greek fermented sausages. LWT—Food Sci. Technol. 2011, 44, 54–61. [Google Scholar] [CrossRef]
- Zdenkova, K.; Alibayov, B.; Karamonova, L.; Purkrtova, S.; Karpiskova, R.; Demnerova, K. Transcriptomic and metabolic responses of Staphylococcus aureus in mixed culture with Lactobacillus plantarum, Streptococcus thermophilus and Enterococcus durans in milk. J. Ind Microbiol. Biotechnol. 2016, 43, 1237–1247. [Google Scholar] [CrossRef]
- Wareing, P. Controlling Clostridium botulinum—Using challenge testing to create safe chilled foods. Leatherhead Food Res. 2017. Available online: https://www.leatherheadfood.com/wp-content/uploads/2017/04/White-paper-45-Controlling-Clostridium-botulinum.pdf (accessed on 24 March 2023).
- Govari, M.; Pexara, A. Nitrates and nitrites in meat products. J. Hell Vet. Med. Soc. 2015, 66, 127–140. [Google Scholar] [CrossRef]
- Cantwell, M.; Elliot, C. Nitrates, Nitrites and Nitrosamines from processed meat intake and colorectal cancer risk. J. Clin. Nutr. Diet. 2017, 3, 27. [Google Scholar] [CrossRef]
- Hatoum, R.; Labrie, S.; Fliss, I. Identification and partial characterization of antilisterial compounds produced by dairy yeasts. Probiotics Antimicrob. Proteins 2013, 5, 8–17. [Google Scholar] [CrossRef]
- Muccilli, S.; Restuccia, C. Bioprotective role of yeasts. Microorganisms 2015, 3, 588–611. [Google Scholar] [CrossRef]
- Goerges, S.; Koslowsky, M.; Velagic, S.; Borst, N.; Bockelmann, W.; Heller, K.J.; Scherer, S. Anti-listerial potential of food-borne yeasts in red smear cheese. Int. Dairy J. 2011, 21, 83–89. [Google Scholar] [CrossRef]
- Sadeghi, A.; Ebrahimi, M.; Shahryari, S.; Kharazmi, M.S.; Jafari, S.M. Food applications of probiotic yeasts; focusing on their techno-functional, postbiotic and protective capabilities. Trends Food Sci. Technol. 2022, 128, 278–295. [Google Scholar] [CrossRef]
- de Lima, M.D.S.F.; de Souza, K.M.S.; Albuquerque, W.W.C.; Teixeira, J.A.C.; Cavalcanti, M.T.H.; Porto, A.L.F. Saccharomyces cerevisiae from Brazilian kefir-fermented milk: An in vitro evaluation of probiotic properties. Microb. Pathog. 2017, 110, 670–677. [Google Scholar] [CrossRef] [PubMed]
- Merchán, A.V.; Benito, M.J.; Galván, A.I.; Ruiz-Moyano Seco de Herrera, S. Identification and selection of yeast with functional properties for future application in soft paste cheese. LWT—Food Sci. Technol. 2020, 124, 109173. [Google Scholar] [CrossRef]
- Yousefi, M.; Khorshidian, N.; Hosseini, H. Potential application of essential oils for mitigation of Listeria monocytogenes in meat and poultry products. Front. Nutr. 2020, 7, 577287. [Google Scholar] [CrossRef]
- Oliveira, M.; Ferreira, V.; Magalhães, R.; Teixeira, P. Biocontrol strategies for Mediterranean-style fermented sausages. Food Res. Int. 2018, 103, 438–449. [Google Scholar] [CrossRef]
- Rangel-Ortega, S.d.C.; Campos-Múzquiz, L.G.; Charles-Rodriguez, A.V.; Chávez-Gonzaléz, M.L.; Palomo-Ligas, L.; Contreras-Esquivel, J.C.; Solanilla-Duque, J.F.; Flores-Gallegos, A.C.; Rodríguez-Herrera, R. Biological control of pathogens in artisanal cheeses. Int. Dairy J. 2023, 140, 105612. [Google Scholar] [CrossRef]
- Food and Drug Administration. Microorganisms and Derived Ingredients Used in Food (Partial List). Available online: https://www.fda.gov/food/generally-recognized-safe-gras/microorganisms-microbial-derived-ingredients-used-food-partial-list (accessed on 20 March 2023).
- Pavli, F.; Argyri, A.A.; Skandamis, P.; Nychas, G.J.; Tassou, C.; Chorianopoulos, N. Antimicrobial activity of oregano essential oil incorporated in sodium alginate edible films: Control of Listeria monocytogenes and spoilage in ham slices treated with high pressure processing. Materials 2019, 12, 3726. [Google Scholar] [CrossRef]
- Kalemba, D.; Kunicka, A. Antibacterial and antifungal properties of essential oils. Curr. Med. Chem. 2005, 10, 813–829. [Google Scholar] [CrossRef]
- De Billerbeck, V.G.; Roques, C.G.; Bessière, J.-M.; Fonvieille, J.-L.; Dargent, R. Effects of Cymbopogon nardus (L.) W. Watson essential oil on the growth and morphogenesis of Aspergillus niger. Can. J. Microbiol. 2001, 47, 9–17. [Google Scholar] [CrossRef]
- Castaño, H.; Ciro, G.; Zapata, J.E.; Jiménez, S. Actividad bactericida del extracto etanólico y del aceite esencial de hojas de Rosmarinus officinalis L. sobre algunas bacterias de interés alimentario. VITAE 2010, 17, 149–154. [Google Scholar]
- Jardak, M.; Elloumi-Mseddi, J.; Aifa, S.; Mnif, S. Chemical composition, anti-biofilm activity and potential cytotoxic effect on cancer cells of Rosmarinus officinalis L. essential oil from Tunisia. Lipids Health Dis. 2017, 16, 190. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Baltar, A.; Pérez-Boto, D.; Medina, M.; Montiel, R. Genomic diversity and characterization of Listeria monocytogenes from dry-cured ham processing plants. Food Microbiol. 2021, 99, 103779. [Google Scholar] [CrossRef]
- Montiel, R.; Peirotén, Á.; Ortiz, S.; Bravo, D.; Gaya, P.; Martínez-Suárez, J.V.; Tapiador, J.; Nuñez, M.; Medina, M. Inactivation of Listeria monocytogenes during dry-cured ham processing. Int. J. Food Microbiol. 2020, 318, 108469. [Google Scholar] [CrossRef] [PubMed]
- El-Sharoud, W.M.; Belloch, C.; Peris, D.; Querol, A. Molecular identification of yeasts associated with traditional Egyptian dairy products. J. Food Sci. 2009, 74, M341–M346. [Google Scholar] [CrossRef]
- Venturini Copetti, M. Yeasts and molds in fermented food production: An ancient bioprocess. Curr. Opin. Food Sci. 2019, 25, 57–61. [Google Scholar] [CrossRef]
- Corsetti, A.; Rossi, J.; Gobbetti, M. Interactions between yeasts and bacteria in the smear surface-ripened cheeses. Int. J. Food Microbiol. 2001, 69, 1–10. [Google Scholar] [CrossRef]
- Ozturk, I.; Sagdic, O.; Yetim, H. Effects of autochthonous yeast cultures on some quality characteristics of traditional Turkish fermented sausage“Sucuk”. Food Sci. Anim. Resour. 2021, 41, 196. [Google Scholar] [CrossRef]
- Ramos-Moreno, L.; Ruiz-Pérez, F.; Rodríguez-Castro, E.; Ramos, J. Debaryomyces hansenii is a real tool to improve a diversity of characteristics in sausages and dry-meat products. Microorganisms 2021, 9, 1512. [Google Scholar] [CrossRef]
- Banjara, N.; Suhr, M.J.; Hallen-Adams, H.E. Diversity of yeast and mold species from a variety of cheese types. Curr Microbiol. 2015, 70, 792–800. [Google Scholar] [CrossRef]
- Riesute, R.; Salomskiene, J.; Moreno, D.S.; Gustiene, S. Effect of yeasts on food quality and safety and possibilities of their inhibition. Trends Food Sci. Technol. 2021, 108, 1–10. [Google Scholar] [CrossRef]
- Banjara, N.; Nickerson, K.W.; Suhr, M.J.; Hallen-Adams, H.E. Killer toxin from several food-derived Debaryomyces hansenii strains effective against pathogenic Candida yeasts. Int. J. Food Microbiol. 2016, 222, 23–29. [Google Scholar] [CrossRef]
- Jacques, N.; Casaregola, S. Safety assessment of dairy microorganisms: The hemiascomycetous yeasts. Int. J. Food Microbiol. 2008, 126, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Godič Torkar, K.; Golk Teger, S. The presence of some pathogen microorganisms, yeasts and moulds in cheese samples produced at small dairy-processing plants. Acta Agric. Slov. 2006, 88, 37–51. [Google Scholar]
- Encinas, J.P.; López-Díaz, T.M.; García-López, M.L.; Otero, A.; Moreno, B. Yeast populations on Spanish fermented sausages. Meat Sci. 2000, 54, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Núñez, F.; Rodríguez, M.M.; Bermúdez, M.E.; Córdoba, J.J.; Asensio, M.A. Composition and toxigenic potential of the mould population on dry-cured Iberian ham. Int. J. Food Microbiol. 1996, 32, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Simoncini, N.; Rotelli, D.; Virgili, R.; Quintavalla, S. Dynamics and characterization of yeasts during ripening of typical Italian dry-cured ham. Food Microbiol. 2007, 24, 577–584. [Google Scholar] [CrossRef]
- Sharaf, O.; Ibrahim, G.; Abd El-Khalik Mahmoud Effat, D.; Ibrahim, A.; Tawfek, N.F.; Effat, B.; El Shafei, K.; F El-Din, H.M.; Salem, M.M. Prevalence of some pathogenic microorganisms in factories Domiati, Feta cheeses and UHT milk in relation to public health sold under market conditions in Cairo. Artic. Int. J. Chem. Tech. Res. 2014, 6, 2807–2814. [Google Scholar]
- Singh, G.; Raksha, A.D.U. Candidal infection: Epidemiology, pathogenesis and recent advances for diagnosis. Bull Pharm. Med. Sci. 2013, 1, 1–8. [Google Scholar]
- Pfaller, M.A.; Diekema, D.J. Role of sentinel surveillance of candidemia: Trends in species distribution and antifungal susceptibility. J. Clin. Microbiol. 2002, 40, 3551–3557. [Google Scholar] [CrossRef]
- Wanderley, L.; Bianchin, A.; Arruda Teo, C.R.P.; Fuentefria, A.M. Occurrence and pathogenicity of Candida spp. in unpasteurized cheese. Braz. J. Biosci. 2013, 11, 145–148. [Google Scholar]
- Seiler, H.; Busse, M. The yeasts of cheese brines. Int. J. Food Microbiol. 1990, 11, 289–303. [Google Scholar] [CrossRef] [PubMed]
- Tzanetakis, N.; Hatzikamari, M.; Litopoulou-Tzanetaki, E. Yeasts of the surface microflora of Feta cheese. In Proceedings of the Yeasts in the Dairy Industry: Positive and Negative Aspects, Copenhagen, Denmark, 2–3 September 1996; pp. 34–43. [Google Scholar]
- Spanamberg, A.; Pais Ramos, J.; Leoncini, O.; Hartz Alves, S.; Valente, P. High frequency of potentially pathogenic yeast species in goat’s raw milk and creamed cheese in Southern Brazil. Acta Sci. Vet. 2009, 37, 133–141. [Google Scholar] [CrossRef]
- Colombo, A.L.; Nucci, M.; Park, B.J.; Nouér, S.A.; Arthington-Skaggs, B.; da Matta, D.A.; Warnock, D.; Morgan, J. Epidemiology of candidemia in Brazil: A nationwide sentinel surveillance of candidemia in eleven medical centers. J. Clin. Microbiol. 2006, 44, 2816. [Google Scholar] [CrossRef] [PubMed]
- Falagas, M.E.; Roussos, N.; Vardakas, K.Z. Relative frequency of albicans and the various non-albicans Candida spp among candidemia isolates from inpatients in various parts of the world: A systematic review. Int. J. Infect. Dis. 2010, 14, e954–e966. [Google Scholar] [CrossRef]
- Miceli, M.H.; Díaz, J.A.; Lee, S.A. Emerging opportunistic yeast infections. Lancet Infect. Dis. 2011, 11, 142–151. [Google Scholar] [CrossRef]
- Groenewald, M.; Boekhout, T.; Neuvéglise, C.; Gaillardin, C.; Van Dijck, P.W.M.; Wyss, M. Yarrowia lipolytica: Safety assessment of an oleaginous yeast with a great industrial potential. Crit. Rev. Microbiol. 2014, 40, 187–206. [Google Scholar] [CrossRef]
- Fröhlich-Wyder, M.T.; Arias-Roth, E.; Jakob, E. Cheese yeasts. Yeast 2019, 36, 129–141. [Google Scholar] [CrossRef]
- Zhang, L.; Huang, C.; Malskær, A.H.; Jespersen, L.; Arneborg, N.; Johansen, P.G. The effects of NaCl and temperature on growth and survival of yeast strains isolated from Danish cheese brines. Curr. Microbiol. 2020, 77, 3377–3384. [Google Scholar] [CrossRef]
- Muccilli, S.; Wemhoff, S.; Restuccia, C.; Meinhardt, F. Exoglucanase-encoding genes from three Wickerhamomyces anomalus killer strains isolated from olive brine. Yeast 2013, 30, 33–43. [Google Scholar] [CrossRef]
- Buzzini, P.; Martini, A. Large-scale screening of selected Candida maltosa, Debaryomyces hansenii and Pichia anomala killer toxin activity against pathogenic yeasts. Med. Mycol. 2001, 39, 479–482. [Google Scholar] [CrossRef]
- Vadkertiová, R.; Sláviková, E. Killer activity of yeasts isolated from natural environments against some medically important Candida species. Pol. J. Microbiol. 2007, 56, 39–43. [Google Scholar] [PubMed]
- Ceugniez, A.; Drider, D.; Jacques, P.; Coucheney, F. Yeast diversity in a traditional French cheese “Tomme d’orchies” reveals infrequent and frequent species with associated benefits. Food Microbiol. 2015, 52, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Makki, G.M.; Kozak, S.M.; Jencarelli, K.G.; Alcaine, S.D. Evaluation of the efficacy of commercial protective cultures to inhibit mold and yeast in cottage cheese. Dairy Sci. 2021, 104, 2709–2718. [Google Scholar] [CrossRef]
- Ravichandran, S.; Muthuraman, S. Examining the anti-candidal activity of 10 selected Indian herbs and investigating the effect of Lawsonia inermis extract on germ tube formation, protease, phospholipase, and aspartate dehydrogenase enzyme activity in Candida albicans. Indian J. Pharmacol. 2016, 48, 47. [Google Scholar]
- Dorsaz, S.; Snäkä, T.; Favre-Godal, Q.; Maudens, P.; Boulens, N.; Furrer, P.; Ebrahimi, S.N.; Hamburger, M.; Allémann, E.; Gindro, K.; et al. Identification and mode of action of a plant natural product targeting human fungal pathogens. Antimicrob. Agents Chemother. 2017, 61, e00829-17. [Google Scholar] [CrossRef] [PubMed]
- Gawlik-Dziki, U.; Durak, A.; Pecio, Ł.; Kowalska, I. Nutraceutical potential of tinctures from fruits, green husks, and leaves of Juglans regia L. Sci. World J. 2014, 2014, 501392. [Google Scholar] [CrossRef]
- Nzeako, B.C.; N Al-Kharousi, Z.S.; Al-Mahrooqui, Z. Antimicrobial activities of clove and thyme extracts. Sultan Qaboos Univ. Med. J. 2006, 6, 33. [Google Scholar]
- Campaniello, D.; Corbo, M.R.; Sinigaglia, M. Antifungal activity of Eugenol against Penicillium, Aspergillus, and Fusarium species. J. Food Prot. 2010, 73, 1124–1128. [Google Scholar] [CrossRef]
- Zhang, T.; Chen, W. The Candida albicans inhibitory activity of the extract from Papaya (Carica papaya L.) Seed relates to mitochondria dysfunction. Int. J. Mol. Sci. 2017, 18, 1858. [Google Scholar] [CrossRef]
- Li, W.R.; Shi, Q.S.; Dai, H.Q.; Liang, Q.; Xie, X.B.; Huang, X.M.; Zhao, G.Z.; Zhang, L.X. Antifungal activity, kinetics and molecular mechanism of action of garlic oil against Candida albicans. Sci. Rep. 2016, 6, 22805. [Google Scholar] [CrossRef]
- Khorshidian, N.; Yousefi, M.; Khanniri, E.; Mortazavian, A.M. Potential application of essential oils as antimicrobial preservatives in cheese. Innov. Food Sci. Emerg. Technol. 2018, 45, 62–72. [Google Scholar] [CrossRef]
- Shabeer, S.; Asad, S.; Jamal, A.; Ali, A. Aflatoxin contamination, its impact and management strategies: An updated review. Toxins 2022, 14, 307. [Google Scholar] [CrossRef] [PubMed]
- Pattono, D.; Grosso, A.; Stocco, P.P.; Pazzi, M.; Zeppa, G. Survey of the presence of patulin and ochratoxin A in traditional semi-hard cheeses. Food Control 2013, 33, 54–57. [Google Scholar] [CrossRef]
- Malir, F.; Ostry, V.; Pfohl-Leszkowicz, A.; Malir, J.; Toman, J. Ochratoxin A: 50 years of research. Toxins 2016, 8, 12–15. [Google Scholar] [CrossRef] [PubMed]
- IARC Aflatoxins. A Review of Human Carcinogens. Chemical Agents and Related Occupations; IARC: Lyon, France, 2012; pp. 225–248. ISBN 978 92 832 1323 9. [Google Scholar]
- IARC; Ochratoxin, A. IARC Monogr Eval Carcinog Risk Chem to Humans; IARC: Lyon, France, 1993; pp. 489–521. [Google Scholar]
- Schrenk, D.; Bodin, L.; Chipman, J.K.; del Mazo, J.; Grasl-Kraupp, B.; Hogstrand, C.; Hoogenboom, L.; Leblanc, J.; Nebbia, C.S.; Nielsen, E.; et al. Risk assessment of ochratoxin A in food. EFSA J. 2020, 18, 6113. [Google Scholar]
- Álvarez, M.; Núñez, F.; Delgado, J.; Andrade, M.J.; Rodrigues, P. Proteomic evaluation of the effect of antifungal agents on Aspergillus westerdijkiae ochratoxin A production in a dry-cured fermented sausage-based medium. Int. J. Food Microbiol. 2022, 379, 109858. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, M.; Delgado, J.; Núñez, F.; Cebrián, E.; Andrade, M.J. Proteomic analyses reveal mechanisms of action of biocontrol agents on ochratoxin A repression in Penicillium nordicum. Food Control 2021, 129, 108232. [Google Scholar] [CrossRef]
- Álvarez, M.; Rodríguez, A.; Peromingo, B.; Núñez, F.; Rodríguez, M. Enterococcus faecium: A promising protective culture to control growth of ochratoxigenic moulds and mycotoxin production in dry-fermented sausages. Mycotoxin Res. 2019, 36, 137–145. [Google Scholar] [CrossRef]
- Delgado, J.; Núñez, F.; Asensio, M.A.; Owens, R.A. Quantitative proteomic profiling of ochratoxin A repression in Penicillium nordicum by protective cultures. Int. J. Food Microbiol. 2019, 305, 108243. [Google Scholar] [CrossRef]
- Meftah, S.; Abid, S.; Dias, T.; Rodrigues, P. Effect of dry-sausage starter culture and endogenous yeasts on Aspergillus westerdijkiae and Penicillium nordicum growth and OTA production. LWT—Food Sci. Technol. 2018, 87, 250–258. [Google Scholar] [CrossRef]
- Cebrián, E.; Núñez, F.; Gálvez, F.J.; Delgado, J.; Bermúdez, E.; Rodríguez, M. Selection and evaluation of Staphylococcus xylosus as a biocontrol agent against toxigenic moulds in a dry-cured ham model system. Microorganisms 2020, 8, 793. [Google Scholar] [CrossRef] [PubMed]
- Simoncini, N.; Virgili, R.; Spadola, G.; Battilani, P. Autochthonous yeasts as potential biocontrol agents in dry-cured meat products. Food Control 2014, 46, 160–167. [Google Scholar] [CrossRef]
- Nazareth, T.D.M.; Calpe, J.; Luz, C.; Mañes, J. Manufacture of a potential antifungal ingredient using lactic acid bacteria from dry-cured sausages. Foods 2023, 12, 1427. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, M.; Delgado, J.; Núñez, F.; Roncero, E.; Andrade, M.J. Proteomic approach to unveil the ochratoxin A repression by Debaryomyces hansenii and rosemary on Penicillium nordicum during dry-cured fermented sausages ripening. Food Control 2022, 137, 108695. [Google Scholar] [CrossRef]
- Peromingo, B.; Núñez, F.; Rodríguez, A.; Alía, A.; Andrade, M.J. Potential of yeasts isolated from dry-cured ham to control ochratoxin A production in meat models. Int. J. Food Microbiol. 2018, 268, 73–80. [Google Scholar] [CrossRef]
- Álvarez, M.; Núñez, F.; Delgado, J.; Andrade, M.J.; Rodríguez, M.; Rodríguez, A. Competitiveness of three biocontrol candidates against ochratoxigenic Penicillium nordicum under dry-cured meat environmental and nutritional conditions. Fungal. Biol. 2021, 129, 108232. [Google Scholar] [CrossRef]
- Rodríguez, A.; Bernáldez, V.; Rodríguez, M.; Andrade, M.J.; Núñez, F.; Córdoba, J.J. Effect of selected protective cultures on ochratoxin A accumulation in dry-cured Iberian ham during its ripening process. LWT—Food Sci. Technol. 2015, 60, 923–928. [Google Scholar] [CrossRef]
- Bernáldez, V.; Córdoba, J.J.; Rodríguez, M.; Cordero, M.; Polo, L.; Rodríguez, A. Effect of Penicillium nalgiovense as protective culture in processing of dry-fermented sausage “ salchichón”. Food Control 2013, 32, 69–76. [Google Scholar] [CrossRef]
- Guimarães, A.; Ramos, Ó.; Cerqueira, M.; Venâncio, A.; Abrunhosa, L. Active whey protein edible films and coatings incorporating Lactobacillus buchneri for Penicillium nordicum control in cheese. Food Bioprocess Technol. 2020, 13, 1074–1086. [Google Scholar] [CrossRef]
- Delgado, J.; Rodríguez, A.; García, A.; Núñez, F.; Asensio, M. Inhibitory effect of PgAFP and protective cultures on Aspergillus parasiticus growth and aflatoxins production on dry-fermented sausage and cheese. Microorganisms 2018, 6, 69. [Google Scholar] [CrossRef]
- Leonelli Pires de Campos, A.C.; Saldanha Nandi, R.D.; Scandorieiro, S.; Gonçalves, M.C.; Reis, G.F.; Dibo, M.; Medeiros, L.P.; Panagio, L.A.; Fagan, E.P.; Takayama Kobayashi, R.K.; et al. Antimicrobial effect of Origanum vulgare (L.) essential oil as an alternative for conventional additives in the Minas cheese manufacture. LWT—Food Sci. Technol. 2022, 157, 113063. [Google Scholar] [CrossRef]
- Vitalini, S.; Nalbone, L.; Bernardi, C.; Iriti, M.; Costa, R.; Cicero, N.; Giarratana, F.; Vallone, L. Ginger and parsley essential oils: Chemical composition, antimicrobial activity, and evaluation of their application in cheese preservation. Nat. Prod. Res. 2022, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, B.I.; Fente, C.; Franco, C.M.; Vazquez, M.J.; Cepeda, A. Inhibitory effects of eugenol and thymol on Penicillium citrinum strains in culture media and cheese. Int. J. Food Microbiol. 2001, 67, 157–163. [Google Scholar] [CrossRef]
- Akrami, F.; Misaghi, A.; Gheisari, H.; Akhondzadeh, A.; Amiri, A.; Razi, S.; Derakhshan, Z.; Dehghani, R. The effect of Zataria multi flora Boiss essential oil on the growth and citrinin production of Penicillium citrinum in culture media and cheese. Food Chem. Toxicol. 2018, 118, 691–694. [Google Scholar]
- Noori, N.; Yahyaraeyat, R.; Khosravi, A.; Atefi, P.; Basti, A.A.; Akrami, F.; Bahonar, A.; Misaghi, A.L.I. Effect of Zataria multiflora Boiss essential oil on growth and citrinin production by Penicillium citrinum in culture media and mozzarella cheese. J. Food Sci. 2012, 32, 445–451. [Google Scholar] [CrossRef]
- Schlösser, I.; Prange, A. Effects of selected natural preservatives on the mycelial growth and ochratoxin A production of the food-related moulds Aspergillus westerdijkiae and Penicillium verrucosum. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess 2019, 36, 1411–1418. [Google Scholar] [CrossRef] [PubMed]
- Císarová, M.; Hleba, L.; Medo, J.; Tančinová, D.; Mašková, Z.; Čuboň, J.; Kováčik, A.; Foltinová, D.; Božik, M.; Klouček, P. The in vitro and in situ effect of selected essential oils in vapour phase against bread spoilage toxicogenic aspergilli. Food Control 2020, 110, 107007. [Google Scholar] [CrossRef]
- Gützkow, K.L.; Al Ayoubi, C.; Soler Vasco, L.; Rohn, S.; Maul, R. Analysis of ochratoxin A, aflatoxin B1 and its biosynthetic precursors in cheese—Method development and market sample screening. Food Control 2022, 143, 109241. [Google Scholar] [CrossRef]
- Rodríguez-Cañás, I.; González-Jartín, J.M.; Alvariño, R.; Alfonso, A.; Vieytes, M.R.; Botana, L.M. Detection of mycotoxins in cheese using an optimized analytical method based on a QuEChERS extraction and UHPLC-MS/MS quantification. Food Chem. 2023, 408, 135182. [Google Scholar] [CrossRef]
- Mareze, J.; Ramos-Pereira, J.; Santos, J.A.; Beloti, V.; López-Díaz, T.M. Identification and characterisation of lactobacilli isolated from an artisanal cheese with antifungal and antibacterial activity against cheese spoilage and mycotoxigenic Penicillium spp. Int. Dairy J. 2022, 130, 105367. [Google Scholar] [CrossRef]
- Kumar, P.; Mahato, D.K.; Kamle, M.; Mohanta, T.K.; Kang, S.G. Aflatoxins: A global concern for food safety, human health and their management. Front. Microbiol. 2017, 7, 2170. [Google Scholar] [CrossRef] [PubMed]
- Pleadin, J.; Zadravec, M.; Brnić, D.; Perković, I.; Škrivanko, M.; Kovačević, D. Moulds and mycotoxins detected in the regional speciality fermented sausage “slavonski kulen” during a 1-year production period. Food Addit. Contam. Part A 2017, 34, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Perrone, G.; Rodriguez, A.; Magistá, D.; Magan, N. Insights into existing and future fungal and mycotoxin contamination of cured meats. Curr. Opin. Food Sci. 2019, 29, 20–27. [Google Scholar] [CrossRef]
- Zadravec, M.; Vahčić, N.; Brnić, D.; Markov, K.; Frece, J.; Beck, R.; Lešić, T.; Pleadin, J. A study of surface moulds and mycotoxins in Croatian traditional dry-cured meat products. Int. J. Food Microbiol. 2020, 317, 108459. [Google Scholar] [CrossRef]
- Carvajal-Moreno, M.; Vargas-Ortiz, M.; Hernández-Camarillo, E.; Ruiz-Velasco, S.; Rojo-Callejas, F. Presence of unreported carcinogens, Aflatoxins and their hydroxylated metabolites, in industrialized Oaxaca cheese from Mexico City. Food Chem. Toxicol. 2019, 124, 128–138. [Google Scholar] [CrossRef]
- Onmaz, N.E.; Çinicioglu, S.; Gungor, C. Moulds and aflatoxins in traditional moldy civil cheese: Presence and public health concerns. J. Hell. Vet. Med. Soc. 2020, 85, 3–11. [Google Scholar]
- Sakin, F.; Tekeli, İ.O.; Yipel, M.; Kürekci, C. Occurrence and health risk assessment of aflatoxins and ochratoxin a in Sürk, a Turkish dairy food, as studied by HPLC. Food Control 2018, 90, 317–323. [Google Scholar] [CrossRef]
- Maragos, C.M.; Probyn, C.; Proctor, R.H.; Sieve, K.K. Cyclopiazonic acid in soft-ripened and blue cheeses marketed in the USA. Food Addit. Contam. Part B 2023, 16, 14–23. [Google Scholar] [CrossRef]
- Ostry, V.; Toman, J.; Grosse, Y.; Malir, F. Cyclopiazonic acid: 50th anniversary of its discovery. World Mycotoxin J. 2018, 11, 135–148. [Google Scholar] [CrossRef]
- Díaz Nieto, C.H.; Granero, A.M.; Zon, M.A.; Fernández, H. Sterigmatocystin: A mycotoxin to be seriously considered. Food Chem. Toxicol. 2018, 118, 460–470. [Google Scholar] [CrossRef]
- IARC. Agents Classified by the IARC Monographs, Volumes 1–123; International Agency for Research on Cancer: Lyon, France, 2018; pp. 1–37.
- Pietri, A.; Leni, G.; Mulazzi, A.; Bertuzzi, T. Ochratoxin A and sterigmatocystin in long-ripened Grana cheese: Occurrence, wheel rind contamination and effectiveness of cleaning techniques on grated products. Toxins 2022, 14, 306. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Tebar, N.; González-Navarro, E.J.; López-Díaz, T.M.; Santos, J.A.; Ortiz de Elguea-Culebras, G.; García-Martínez, M.M.; Molina, A.; Carmona, M.; Berruga, M.I. Biological activity of extracts from aromatic plants as control agents against spoilage molds isolated from sheep cheese. Foods 2021, 10, 1576. [Google Scholar] [CrossRef]
- Coton, M.; Au, A.; Poirier, E.; Debaets, S.; Coton, E.; Dantigny, P. Production and migration of ochratoxin A and citrinin in Comté cheese by an isolate of Penicillium verrucosum selected among Penicillium spp. mycotoxin producers in YES medium. Food Microbiol. 2019, 82, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Gayathri, L.; Dhivya, R.; Dhanasekaran, D.; Periasamy, V.S.; Alshatwi, A.A.; Akbarsha, M.A. Hepatotoxic effect of ochratoxin A and citrinin, alone and in combination, and protective effect of vitamin E: In vitro study in HepG2 cell. Food Chem. Toxicol. 2015, 83, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Kamle, M.; Mahato, D.K.; Gupta, A.; Pandhi, S.; Sharma, N.; Sharma, B.; Mishra, S.; Arora, S.; Selvakumar, R.; Saurabh, V.; et al. Citrinin mycotoxin contamination in food and feed: Impact on agriculture, human health, and detection and management strategies. Toxins 2022, 14, 85. [Google Scholar] [CrossRef]
- Schmidt-Heydt, M.; Stoll, D.; Schütz, P.; Geisen, R. Oxidative stress induces the biosynthesis of citrinin by Penicillium verrucosum at the expense of ochratoxin. Int. J. Food Microbiol. 2015, 192, 1–6. [Google Scholar] [CrossRef]
- Schmidt-Heydt, M.; Graf, E.; Stoll, D.; Geisen, R. The biosynthesis of ochratoxin A by Penicillium as one mechanism for adaptation to NaCl rich foods. Food Microbiol. 2012, 29, 233–241. [Google Scholar] [CrossRef]
- Tsuda, H.; Matsumoto, T.; Ishimi, Y. Selection of lactic acid bacteria as starter cultures for fermented meat products. Food Sci. Technol. Res. 2012, 18, 713–721. [Google Scholar] [CrossRef]
- Paik, H.D.; Lee, J.Y. Investigation of reduction and tolerance capability of lactic acid bacteria isolated from kimchi against nitrate and nitrite in fermented sausage condition. Meat Sci. 2014, 97, 609–614. [Google Scholar] [CrossRef]
- Zagorec, M.; Champomier-Vergès, M.C. Lactobacillus sakei: A starter for sausage fermentation, a protective culture for meat products. Microorganisms 2017, 5, 56. [Google Scholar] [CrossRef]
- Crowther, T.W.; Boddy, L.; Maynard, D.S. The use of artificial media in fungal ecology. Fungal. Ecol. 2018, 32, 87–91. [Google Scholar] [CrossRef]
- Álvarez, M.; Andrade, M.J.; Núñez, F.; Rodríguez, M.; Delgado, J. Proteomics as a new-generation tool for studying moulds related to food safety and quality. Int. J. Mol. Sci. 2023, 24, 4709. [Google Scholar] [CrossRef] [PubMed]
- Picozzi, C.; Bonacina, G.; Vigentini, I.; Foschino, R. Genetic diversity in Italian Lactobacillus sanfranciscensis strains assessed by multilocus sequence typing and pulsed-field gel electrophoresis analyses. Microbiology 2010, 156, 2035–2045. [Google Scholar] [CrossRef] [PubMed]
- Andrade, M.J.; Rodríguez, M.; Sánchez, B.; Aranda, E.; Córdoba, J.J. DNA typing methods for differentiation of yeasts related to dry-cured meat products. Int. J. Food Microbiol. 2006, 107, 48–58. [Google Scholar] [CrossRef]
- Andrade, M.J.; Rodríguez, M.; Casado, E.; Córdoba, J.J. Efficiency of mitochondrial DNA restriction analysis and RAPD-PCR to characterize yeasts growing on dry-cured Iberian ham at the different geographic areas of ripening. Meat Sci. 2010, 84, 377–383. [Google Scholar] [CrossRef]
- Sánchez, B.; Rodríguez, M.; Casado, E.M.; Martín, A.; Córdoba, J.J. Development of an efficient fungal DNA extraction method to be used in random amplified polymorphic DNA-PCR analysis to differentiate cyclopiazonic acid mold producers. J. Food Prot. 2008, 71, 2497–2503. [Google Scholar] [CrossRef]

| Matrix | Pathogenic Bacteria | Biocontrol Agent | Pathogenic Bacteria Viability Reduction | Reference |
|---|---|---|---|---|
| Spanish dry-cured fermented sausages | Listeria monocytogenes | Latilactobacillus sakei 197 | 1.77 log CFU g−1 | [65] |
| Short-ripening Spanish traditional cheeses | L. monocytogenes | Lacticaseibacillus casei 116 | 2.2 log CFU g−1 | [66] |
| Brazilian artisanal cheeses | L. monocytogenes and Staphylococcus aureus | Lactiplantibacillus plantarum (1QB77) | 3 and 2.3 log CFU g−1, respectively | [67] |
| Frescal and semi-hard artisanal Minas microcheeses | L. monocytogenes | Levilactobacillus brevis L. plantarum Enterococcus faecalis | Inactivation | [36] |
| Dry-fermented Greek sausage | L. monocytogenes and Escherichia coli | L. sakei 8416 L. sakei 4413 | 2.2 log CFU g−1 | [68] |
| Bryndza cheese | Salmonella enterica and S. aureus | Lactococcus lactis Lactiplantibacillus paraplantarum | Significative reduction | [69] |
| Traditional dry-fermented sausage suçuk | S. aureus | L. sakei Staphylococcus carnosus | More than 2 log CFU g−1 | [70] |
| Pediococcus acidilactici, Latilactobacillus curvatus, Staphylococcus xylosus | More than 2 log CFU g−1 | |||
| Fermented salami | Clostridium perfringens and Clostridium species | L. plantarum PCS20, Lactobacillus delbrueckii DSM 20074 | 2 and 1.5 log CFU g−1, respectively | [71] |
| Dry fermented sausages | E. coli, L. monocytogenes, Salmonella spp. and sulfite-reducing clostridia | Juniperus communis L. Essential oil (EOs) | Inactivation | [72] |
| Dry cured sausage ‘Chouriço de vinho’ | Salmonella spp., L. monocytogenes and S. aureus | Bay, garlic, nutmeg, oregano, rosemary and thyme EOs | 0.1–2 log CFU g−1 | [42] |
| Dry cured sausage ‘Chouriço de vinho’ | S. aureus | Thyme EOs | Inactivation | [42] |
| Cured ham-based medium | L. monocytogenes | Cinnamon, pomegranate and strawberry extracts | 3 log CFU mL−1 | [73] |
| Ochratoxin A (OTA) Producers | ||||
|---|---|---|---|---|
| Matrix | Mold | Biocontrol Agent | Effect on Growth or Mycotoxin Production | Reference |
| Dry-cured fermented sausage medium | Penicillium nordicum | Enterococcus faecium | ↓ 1 OTA ↓ Growth | [157] |
| Dry-cured ham based medium | P. nordicum | Staphylococcus xylosus | ↓ OTA ↓ Growth | [160] |
| Fermented meat model system | P. nordicum and Penicillium verrucosum | Lactiplantibacillus plantarum and Pediococcus pentosaceus | ↓ Growth | [162] |
| Ham, industrial and traditional dry-cured sausages | P. nordicum and Aspergillus westerdijkiaie | Candida zeylanoides and Rothia mucilaginosa | ↓ OTA ↓ Growth | [159] |
| Speck | P. nordicum and Aspergillus ochraceus | Debaryomyces hansenii and Saccharomycopsis fibuligera | ↓ OTA ↓ Growth | [37] |
| Dry-cured fermented sausages | P. nordicum | D. hansenii | ↓ OTA | [31,161,163] |
| Dry-cured meat model systems | P. verrucosum, P. nordicum | D. hansenii | ↓ OTA ↓ Growth | [156,164] |
| Dry-cured fermented sausage medium | A. westerdijkiae | D. hansenii | ↓ OTA | [155] |
| Dry-cured ham based medium | P. nordicum | Penicillium chrysogenum | ↓ OTA | [158] |
| Meat model system | P. nordicum | P. chrysogenum | ↓ OTA ↓ Growth | [165] |
| Dry-cured Iberian ham | Wild mycobiota containing OTA producer strains | P. chrysogenum | ↓ OTA | [166] |
| Dry-cured fermented sausage | P. verrucosum | P. nalgiovense | ↓ OTA | [167] |
| Dry-cured fermented sausage medium | P. nordicum | Rosemary and oregano leaves | ↓ OTA | [30] |
| Dry-cured fermented sausage medium | P. nordicum | Smoked paprika “pimentón” | ↓ OTA | [44] |
| Dry-cured fermented sausages | P. nordicum | Rosemary leaves | ↓ OTA | [163] |
| Dry-cured fermented sausage medium | P. nordicum | Rosemary leaves and rosemary essential oil (EO) | ↓ OTA | [40] |
| Cheeses | P. nordicum | Lactobacillus buchneri and Lacticaseibacillus casei | ↓ OTA | [168] |
| Aflatoxins (AFs) producers | ||||
| Dry-cured ham based medium | Aspergillus flavus | S. xylosus | ↓ AFB1 ↓ Growth | [160] |
| Dry-cured ham based medium | Aspergillus parasiticus | S. xylosus | ↓ AFB1 and AFG1 ↓ Growth | [160] |
| Fermented meat model system | A. flavus and A. parasiticus | L. plantarum and P. pentosaceus | ↓ Growth | [162] |
| Dry-cured fermented sausages slices | A. parasiticus | D. hansenii + PgAFP + Pediococcus acidilactici | ↓ AFB1 and AFG1 ↓ Growth | [169] |
| Dry-cured fermented sausages; dry-cured ham | A. parasiticus | D. hansenii | ↓ AFB1 and AFG1 | [39] |
| Dry-cured meat model system | A. parasiticus | Smoked paprika“pimentón” | ↓ AFB1 and AFG1 | [44] |
| Cheese slices | A. parasiticus | D. hansenii + PgAFP + P. acidilactici | ↓ AFG1 ↓ Growth | [169] |
| Minas cheese formulation | A. flavus | Origanum vulgare EO | ↓ Spores germination | [170] |
| Cheese slices | A. flavus | Parsley EO | ↓ Growth | [171] |
| Kashar cheese | A. parasiticus | Thymol | ↓ Growth | [45] |
| Cyclopiazonic acid A (CPA) producers | ||||
| Dry-cured ham-based model | Penicillium griseofulvum | S. xylosus | ↓ CPA | [160] |
| Fermented meat model system | P. griseofulvum and Penicillium commune | Lc. plantarum and P. pentosaceus | ↓ Growth | [162] |
| Dry-fermented sausage-based medium; dry-fermented sausages | P. griseofulvum | PgAFP | ↓ CPA | [34] |
| Edam cheese | P. commune | Clove, thyme, red thyme, litsea, cumin and marjoram eOs | ↓ Growth | [43] |
| Citrinin (CIT) producer | ||||
| Arzúa-Ulloa cheese | Penicillium citrinum | Eugenol and thymol | ↓ CIT | [172] |
| Iranian and mozzarella cheeses | P. citrinum | Zataria multiflora Boiss EO | ↓ CIT | [173,174] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delgado, J.; Álvarez, M.; Cebrián, E.; Martín, I.; Roncero, E.; Rodríguez, M. Biocontrol of Pathogen Microorganisms in Ripened Foods of Animal Origin. Microorganisms 2023, 11, 1578. https://doi.org/10.3390/microorganisms11061578
Delgado J, Álvarez M, Cebrián E, Martín I, Roncero E, Rodríguez M. Biocontrol of Pathogen Microorganisms in Ripened Foods of Animal Origin. Microorganisms. 2023; 11(6):1578. https://doi.org/10.3390/microorganisms11061578
Chicago/Turabian StyleDelgado, Josué, Micaela Álvarez, Eva Cebrián, Irene Martín, Elia Roncero, and Mar Rodríguez. 2023. "Biocontrol of Pathogen Microorganisms in Ripened Foods of Animal Origin" Microorganisms 11, no. 6: 1578. https://doi.org/10.3390/microorganisms11061578
APA StyleDelgado, J., Álvarez, M., Cebrián, E., Martín, I., Roncero, E., & Rodríguez, M. (2023). Biocontrol of Pathogen Microorganisms in Ripened Foods of Animal Origin. Microorganisms, 11(6), 1578. https://doi.org/10.3390/microorganisms11061578