
Viable Mycobacterium avium subsp. paratuberculosis Colonizes Peripheral Blood of Inflammatory Bowel Disease Patients

 , , , , ,
, , , , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
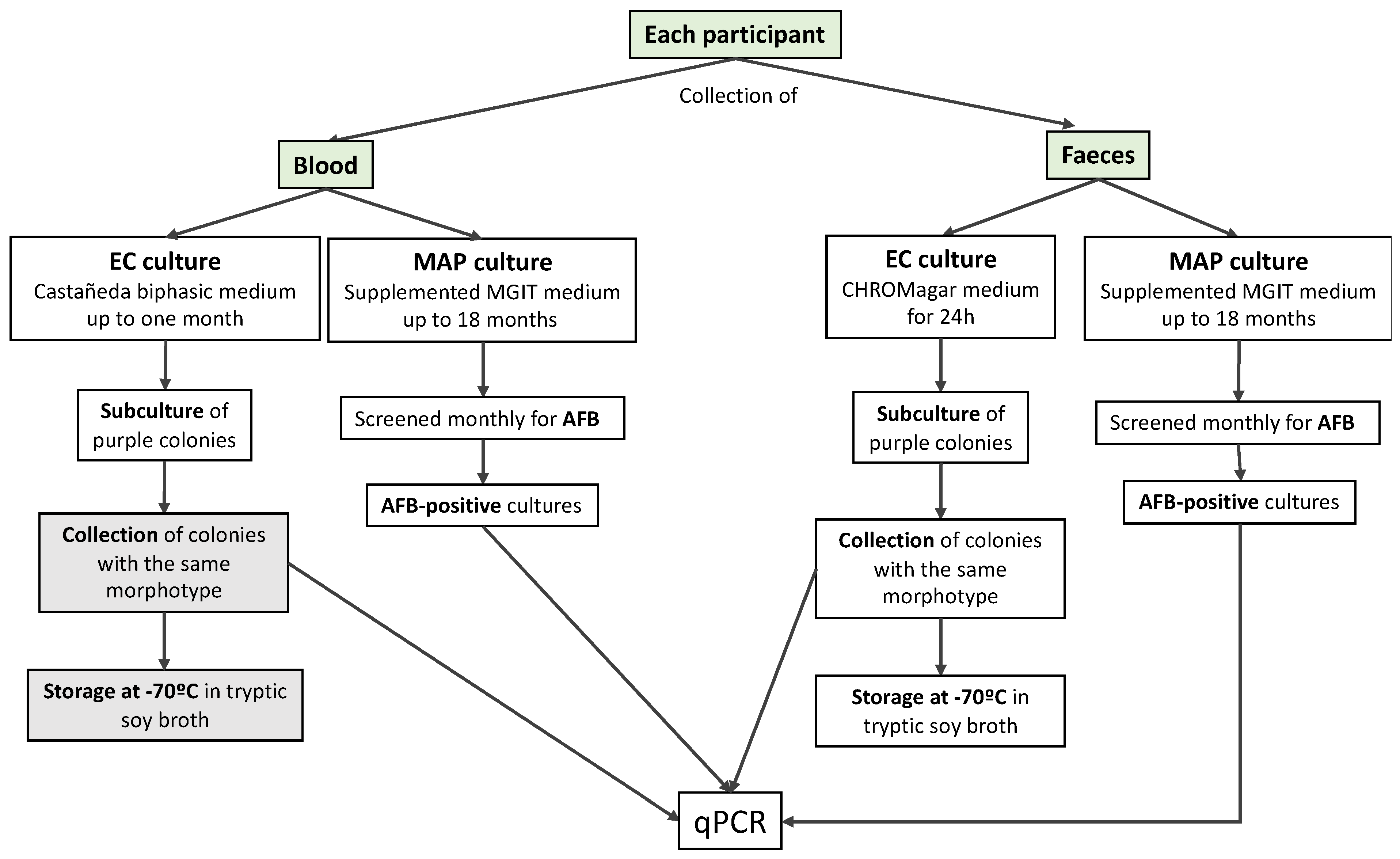
2.1. Participants and Sample Collection
2.2. Blood Cultures
2.3. Faecal Cultures
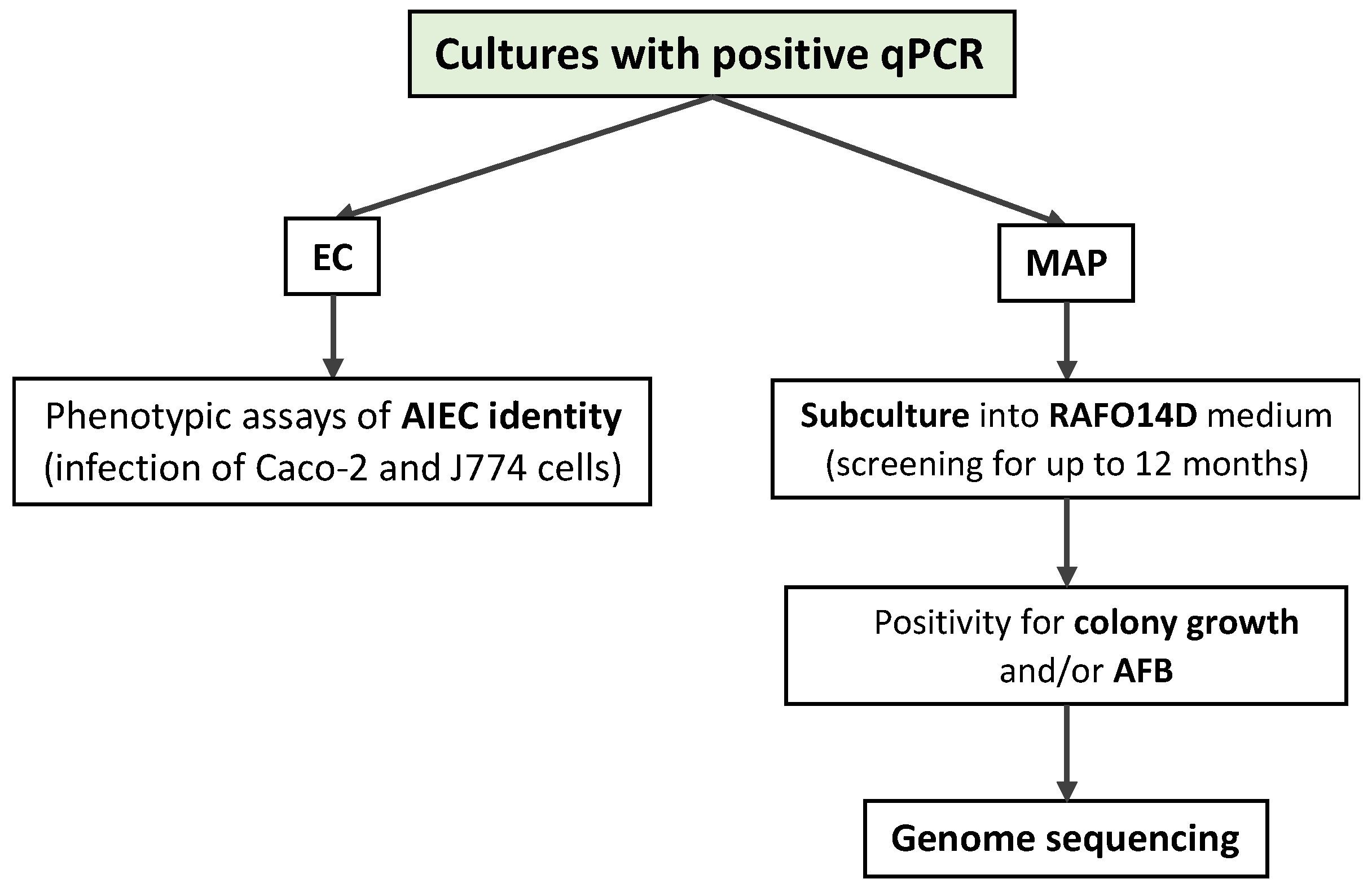
2.4. Confirmation of E. coli and MAP Identity Using Real-Time PCR
2.5. Investigation of AIEC Identity
2.6. MAP Sub-Culture and Genome Sequencing
3. Results
3.1. Characterization of the Cohort
3.2. E. coli Culture and AIEC Identity
3.3. MAP Culture
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sila, S.; Jelić, M.; Trivić, I.; Tambić Andrašević, A.; Kolaček, S.; Hojsak, I. Gut Microbiota Composition Changes Following Discontinuation of Exclusive Enteral Nutrition in Children with Crohn’s Disease. Microorganisms 2023, 11, 505. [Google Scholar] [CrossRef] [PubMed]
- Estevinho, M.M.; Leão Moreira, P.; Silva, I.; Laranjeira Correia, J.; Santiago, M.; Magro, F. A Scoping Review on Early Inflammatory Bowel Disease: Definitions, Pathogenesis, and Impact on Clinical Outcomes. Therap. Adv. Gastroenterol. 2022, 15, 17562848221142672. [Google Scholar] [CrossRef]
- Shin, S.Y.; Kim, S.; Choi, J.W.; Kang, S.-B.; Kim, T.O.; Seo, G.S.; Cha, J.M.; Chun, J.; Jung, Y.; Im, J.P.; et al. The Common and Unique Pattern of Microbiome Profiles among Saliva, Tissue, and Stool Samples in Patients with Crohn’s Disease. Microorganisms 2022, 10, 1467. [Google Scholar] [CrossRef] [PubMed]
- Rizzello, F.; Spisni, E.; Giovanardi, E.; Imbesi, V.; Salice, M.; Alvisi, P.; Valerii, M.C.; Gionchetti, P. Implications of the Westernized Diet in the Onset and Progression of IBD. Nutrients 2019, 11, 1033. [Google Scholar] [CrossRef]
- Alam, M.T.; Amos, G.C.A.; Murphy, A.R.J.; Murch, S.; Wellington, E.M.H.; Arasaradnam, R.P. Microbial Imbalance in Inflammatory Bowel Disease Patients at Different Taxonomic Levels. Gut Pathog. 2020, 12, 1. [Google Scholar] [CrossRef]
- Franzosa, E.A.; Sirota-Madi, A.; Avila-Pacheco, J.; Fornelos, N.; Haiser, H.J.; Reinker, S.; Vatanen, T.; Hall, A.B.; Mallick, H.; McIver, L.J.; et al. Gut Microbiome Structure and Metabolic Activity in Inflammatory Bowel Disease. Nat. Microbiol. 2019, 4, 293–305. [Google Scholar] [CrossRef]
- Rossi, E.; Leccese, G.; Baldelli, V.; Bibi, A.; Scalone, E.; Camilloni, C.; Paroni, M.; Landini, P. Inactivation of the Pyrimidine Biosynthesis PyrD Gene Negatively Affects Biofilm Formation and Virulence Determinants in the Crohn’s Disease-Associated Adherent Invasive Escherichia coli LF82 Strain. Microorganisms 2022, 10, 537. [Google Scholar] [CrossRef]
- Keown, D.A.; Collings, D.A.; Keenan, J.I. Uptake and Persistence of Mycobacterium avium Subsp. Paratuberculosis in Human Monocytes. Infect. Immun. 2012, 80, 3768–3775. [Google Scholar] [CrossRef]
- Negroni, A.; Costanzo, M.; Vitali, R.; Superti, F.; Bertuccini, L.; Tinari, A.; Minelli, F.; Di Nardo, G.; Nuti, F.; Pierdomenico, M.; et al. Characterization of Adherent-Invasive Escherichia coli Isolated from Pediatric Patients with Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2012, 18, 913–924. [Google Scholar] [CrossRef]
- Barnich, N.; Darfeuille-Michaud, A. Adherent-Invasive Escherichia coli and Crohn’s Disease. Curr. Opin. Gastroenterol. 2007, 23, 16–20. [Google Scholar] [CrossRef]
- Barnich, N.; Denizot, J.; Darfeuille-Michaud, A. E. coli-Mediated Gut Inflammation in Genetically Predisposed Crohn’s Disease Patients. Pathol. Biol. 2013, 61, e65–e69. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Ullah, N.; Zha, L.; Bai, Y.; Khan, A.; Zhao, T.; Che, T.; Zhang, C. Alteration of Gut Microbiota in Inflammatory Bowel Disease (IBD): Cause or Consequence? IBD Treatment Targeting the Gut Microbiome. Pathogens 2019, 8, 126. [Google Scholar] [CrossRef] [PubMed]
- Zamani, S.; Zali, M.R.; Aghdaei, H.A.; Sechi, L.A.; Niegowska, M.; Caggiu, E.; Keshavarz, R.; Mosavari, N.; Feizabadi, M.M. Mycobacterium avium Subsp. Paratuberculosis and Associated Risk Factors for Inflammatory Bowel Disease in Iranian Patients. Gut Pathog. 2017, 9, 1. [Google Scholar] [CrossRef]
- Renouf, M.J.; Cho, Y.H.; McPhee, J.B. Emergent Behavior of IBD-Associated Escherichia coli During Disease. Inflamm. Bowel Dis. 2019, 25, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, G.; Clancy, A.; Huynh, R.; Borody, T. Profound Remission in Crohn’s Disease Requiring No Further Treatment for 3–23 Years: A Case Series. Gut Pathog. 2020, 12, 16. [Google Scholar] [CrossRef] [PubMed]
- Carrière, J.; Darfeuille-Michaud, A.; Nguyen, H.T.T. Infectious Etiopathogenesis of Crohn’s Disease. World J. Gastroenterol. 2014, 20, 12102–12117. [Google Scholar] [CrossRef] [PubMed]
- Campos, N.; Magro, F.; Castro, A.R.; Cabral, J.; Rodrigues, P.; Silva, R.; Appelberg, R.; Rodrigues, S.; Lopes, S.; Macedo, G.; et al. Macrophages from IBD Patients Exhibit Defective Tumour Necrosis Factor-α Secretion but Otherwise Normal or Augmented pro-Inflammatory Responses to Infection. Immunobiology 2011, 216, 961–970. [Google Scholar] [CrossRef]
- Cirone, K.M.; Lahiri, P.; Holani, R.; Tan, Y.L.; Arrazuria, R.; De Buck, J.; Barkema, H.W.; Cobo, E.R. Synthetic Cathelicidin LL-37 Reduces Mycobacterium avium Subsp. Paratuberculosis Internalization and pro-Inflammatory Cytokines in Macrophages. Cell Tissue Res. 2020, 379, 207–217. [Google Scholar] [CrossRef]
- Nighot, P.K.; Hu, C.-A.A.; Ma, T.Y. Autophagy Enhances Intestinal Epithelial Tight Junction Barrier Function by Targeting Claudin-2 Protein Degradation. J. Biol. Chem. 2015, 290, 7234–7246. [Google Scholar] [CrossRef]
- Zhang, H.; Zheng, L.; McGovern, D.P.B.; Hamill, A.M.; Ichikawa, R.; Kanazawa, Y.; Luu, J.; Kumagai, K.; Cilluffo, M.; Fukata, M.; et al. Myeloid ATG16L1 Facilitates Host-Bacteria Interactions in Maintaining Intestinal Homeostasis. J. Immunol. 2017, 198, 2133–2146. [Google Scholar] [CrossRef]
- Schirmer, M.; Garner, A.; Vlamakis, H.; Xavier, R.J. Microbial genes and pathways in inflammatory bowel disease. Nat. Rev. Microbiol. 2019, 17, 497–511. [Google Scholar] [CrossRef]
- Chassaing, B.; Koren, O.; Carvalho, F.A.; Ley, R.E.; Gewirtz, A.T. AIEC Pathobiont Instigates Chronic Colitis in Susceptible Hosts by Altering Microbiota Composition. Gut 2014, 63, 1069–1080. [Google Scholar] [CrossRef]
- Arrazuria, R.; Elguezabal, N.; Juste, R.A.; Derakhshani, H.; Khafipour, E. Mycobacterium avium Subspecies Paratuberculosis Infection Modifies Gut Microbiota under Different Dietary Conditions in a Rabbit Model. Front. Microbiol. 2016, 7, 446. [Google Scholar] [CrossRef]
- Nazareth, N.; Magro, F.; Appelberg, R.; Silva, J.; Gracio, D.; Coelho, R.; Cabral, J.M.; Abreu, C.; Macedo, G.; Bull, T.J.; et al. Increased Viability but Decreased Culturability of Mycobacterium avium Subsp. Paratuberculosis in Macrophages from Inflammatory Bowel Disease Patients under Infliximab Treatment. Med. Microbiol. Immunol. 2015, 204, 647–656. [Google Scholar] [CrossRef]
- Nazareth, N.; Magro, F.; Machado, E.; Ribeiro, T.G.; Martinho, A.; Rodrigues, P.; Alves, R.; Macedo, G.N.; Gracio, D.; Coelho, R.; et al. Prevalence of Mycobacterium avium Subsp. Paratuberculosis and Escherichia coli in Blood Samples from Patients with Inflammatory Bowel Disease. Med. Microbiol. Immunol. 2015, 204, 681–692. [Google Scholar] [CrossRef]
- Elguezabal, N.; Bastida, F.; Sevilla, I.A.; González, N.; Molina, E.; Garrido, J.M.; Juste, R.A. Estimation of Mycobacterium avium Subsp. Paratuberculosis Growth Parameters: Strain Characterization and Comparison of Methods. Appl. Environ. Microbiol. 2011, 77, 8615–8624. [Google Scholar] [CrossRef]
- Bull, T.J.; Munshi, T.; Mikkelsen, H.; Hartmann, S.B.; Sørensen, M.R.; Garcia, J.S.; Lopez-Perez, P.M.; Hofmann, S.; Hilpert, K.; Jungersen, G. Improved Culture Medium (TiKa) for Mycobacterium avium Subspecies Paratuberculosis (MAP) Matches QPCR Sensitivity and Reveals Significant Proportions of Non-Viable MAP in Lymphoid Tissue of Vaccinated MAP Challenged Animals. Front. Microbiol. 2016, 7, 2112. [Google Scholar] [CrossRef] [PubMed]
- Castaneda, M.R. A Practical Method for Routine Blood Cultures in Brucellosis. Proc. Soc. Exp. Biol. Med. 1947, 64, 114–115. [Google Scholar] [CrossRef] [PubMed]
- Sidoti, F.; Banche, G.; Astegiano, S.; Allizond, V.; Cuffini, A.M.; Bergallo, M. Validation and Standardization of IS900 and F57 Real-Time Quantitative PCR Assays for the Specific Detection and Quantification of Mycobacterium avium Subsp. Paratuberculosis. Can. J. Microbiol. 2011, 57, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.F.; Cao, W.W.; Cerniglia, C.E. PCR Detection and Quantitation of Predominant Anaerobic Bacteria in Human and Animal Fecal Samples. Appl. Environ. Microbiol. 1996, 62, 1242–1247. [Google Scholar] [CrossRef] [PubMed]
- Bull, T.J.; McMinn, E.J.; Sidi-Boumedine, K.; Skull, A.; Durkin, D.; Neild, P.; Rhodes, G.; Pickup, R.; Hermon-Taylor, J. Detection and Verification of Mycobacterium avium Subsp. Paratuberculosis in Fresh Ileocolonic Mucosal Biopsy Specimens from Individuals with and without Crohn’s Disease. J. Clin. Microbiol. 2003, 41, 2915–2923. [Google Scholar] [CrossRef] [PubMed]
- Camprubí-Font, C.; Lopez-Siles, M.; Ferrer-Guixeras, M.; Niubó-Carulla, L.; Abellà-Ametller, C.; Garcia-Gil, L.J.; Martinez-Medina, M. Comparative Genomics Reveals New Single-Nucleotide Polymorphisms That Can Assist in Identification of Adherent-Invasive Escherichia coli. Sci. Rep. 2018, 8, 2695. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Aligning Sequence Reads, Clone Sequences and Assembly Contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. Subgroup, 1000 Genome Project Data Processing The Sequence Alignment/Map Format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Hall, B.; Holleran, G.; McNamara, D. Small Bowel Crohn’s Disease: An Emerging Disease Phenotype? Dig. Dis. 2015, 33, 42–51. [Google Scholar] [CrossRef]
- Hall, M.M.; Mueske, C.A.; Ilstrup, D.M.; Washington 2nd, J.A. Evaluation of a Biphasic Medium for Blood Cultures. J. Clin. Microbiol. 1979, 10, 673–676. [Google Scholar] [CrossRef]
- Larabi, A.; Barnich, N.; Nguyen, H.T.T. New insights into the interplay between autophagy, gut microbiota and inflammatory responses in IBD. Autophagy 2020, 16, 38–51. [Google Scholar] [CrossRef]
- Buttó, L.F.; Schaubeck, M.; Haller, D. Mechanisms of Microbe-Host Interaction in Crohn’s Disease: Dysbiosis vs. Pathobiont Selection. Front. Immunol. 2015, 6, 555. [Google Scholar] [CrossRef]
- Singh, R.; Rossini, V.; Stockdale, S.R.; Saiz-Gonzalo, G.; Hanrahan, N.; D’Souza, T.; Clooney, A.; Draper, L.A.; Hill, C.; Nally, K.; et al. An IBD-Associated Pathobiont Synergises with NSAID to Promote Colitis Which Is Blocked by NLRP3 Inflammasome and Caspase-8 Inhibitors. Gut Microbes 2023, 15, 2163838. [Google Scholar] [CrossRef]
- da Silva Santos, A.C.; Gomes Romeiro, F.; Yukie Sassaki, L.; Rodrigues, J. Escherichia coli from Crohn’s Disease Patient Displays Virulence Features of Enteroinvasive (EIEC), Enterohemorragic (EHEC), and Enteroaggregative (EAEC) Pathotypes. Gut Pathog. 2015, 7, 2. [Google Scholar] [CrossRef]
- Kittana, H.; Gomes-Neto, J.C.; Heck, K.; Juritsch, A.F.; Sughroue, J.; Xian, Y.; Mantz, S.; Segura Muñoz, R.R.; Cody, L.A.; Schmaltz, R.J.; et al. Evidence for a Causal Role for Escherichia coli Strains Identified as Adherent-Invasive (AIEC) in Intestinal Inflammation. mSphere 2023, 8, e0047822. [Google Scholar] [CrossRef] [PubMed]
- Mirsepasi-Lauridsen, H.C.; Vallance, B.A.; Krogfelt, K.A.; Petersen, A.M. Escherichia Coli Pathobionts Associated with Inflammatory Bowel Disease. Clin. Microbiol. Rev. 2019, 32, e00060-18. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-S.; Back, Y.W.; Son, Y.-J.; Kim, H.-J. Mycobacterium avium Subsp. Paratuberculosis MAP1889c Protein Induces Maturation of Dendritic Cells and Drives Th2-Biased Immune Responses. Cells 2020, 9, 944. [Google Scholar] [CrossRef] [PubMed]
- Rees, W.D.; Lorenzo-Leal, A.C.; Steiner, T.S.; Bach, H. Mycobacterium avium Subspecies Paratuberculosis Infects and Replicates within Human Monocyte-Derived Dendritic Cells. Microorganisms 2020, 8, 994. [Google Scholar] [CrossRef] [PubMed]
- Kuenstner, J.T.; Potula, R.; Bull, T.J.; Grant, I.R.; Foddai, A.; Naser, S.A.; Bach, H.; Zhang, P.; Yu, D.; Lu, X.; et al. Presence of Infection by Mycobacterium avium Subsp. Paratuberculosis in the Blood of Patients with Crohn’s Disease and Control Subjects Shown by Multiple Laboratory Culture and Antibody Methods. Microorganisms 2020, 8, 2054. [Google Scholar] [CrossRef]
- van der Sloot, K.W.J.; Voskuil, M.D.; Blokzijl, T.; Dinkla, A.; Ravesloot, L.; Visschedijk, M.C.; van Dullemen, H.M.; Festen, E.A.M.; Alizadeh, B.Z.; van Leer-Buter, C.; et al. Isotype-Specific Antibody Responses to Mycobacterium avium Paratuberculosis Antigens Are Associated With the Use of Biologic Therapy in Inflammatory Bowel Disease. J. Crohns. Colitis 2021, 15, 1253–1263. [Google Scholar] [CrossRef]
- Noth, R.; Stüber, E.; Häsler, R.; Nikolaus, S.; Kühbacher, T.; Hampe, J.; Bewig, B.; Schreiber, S.; Arlt, A. Anti-TNF-α Antibodies Improve Intestinal Barrier Function in Crohn’s Disease. J. Crohns. Colitis 2012, 6, 464–469. [Google Scholar] [CrossRef]
- Zhao, N.; Cao, J.; Xu, J.; Liu, B.; Liu, B.; Chen, D.; Xia, B.; Chen, L.; Zhang, W.; Zhang, Y.; et al. Targeting RNA with Next- and Third-Generation Sequencing Improves Pathogen Identification in Clinical Samples. Adv. Sci. 2021, 8, e2102593. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| HC | CD | UC | CIR | ||
|---|---|---|---|---|---|
| Number (n) | - | 22 | 18 | 15 | 7 |
| Age | Mean | 66.0 | 34.4 | 46.5 | 67.0 |
| Range | 14–84 | 18–65 | 18–72 | 57–83 | |
| Gender (%) | Male | 15 | 10 | 10 | 7 |
| Female | 6 | 8 | 5 | 0 | |
| Disease duration (years) | Mean | — | 10.4 | 11.0 | 4.4 |
| Range | — | 1–24 | 3–20 | 0.2–14 | |
| Montreal classification (CD) | A1 | — | 0 | — | — |
| A2 | — | 12 | — | — | |
| A3 | — | 3 | — | — | |
| L1 | — | 6 | — | — | |
| L2 | — | 4 | — | — | |
| L3 | — | 3 | — | — | |
| B1 | — | 10 | — | — | |
| B2 | — | 2 | — | — | |
| B3 | — | 4 | — | — | |
| Montreal classification (UC) | E1 | — | — | 2 | — |
| E2 | — | — | 9 | — | |
| E3 | — | — | 4 | — | |
| Harvey–Bradshaw (CD) | <5 (remission) | — | 16 | — | — |
| 5–7 (mild) | — | 2 | — | — | |
| Mayo (UC) | 0–1 (remission) | — | — | 15 | — |
| Child–Pugh classification (cirrhosis) | A | — | — | — | 3 |
| B | — | — | — | 3 | |
| C | — | — | — | 1 | |
| Therapy | ADA/IFX | — | 10 | 7 | — |
| ADA/IFX + AZA | — | 5 | 2 | — | |
| ADA/IFX + 5-ASA | — | 1 | 2 | — | |
| AZA | — | 2 | 0 | — | |
| ADA/IFX + AZA + 5-ASA | — | 0 | 4 | — |
| No Presumptive E. coli Isolates | One Presumptive E. coli Isolate | Two Presumptive E. coli Isolates (A and B) | |
|---|---|---|---|
| HC | HC3, HC4, HC19 | HC7, HC8, HC10, HC12, HC13, HC14, HC16, HC18, HC21 | HC1, HC2, HC5, HC6, HC9, HC11, HC15, HC17, HC20, HC22 |
| CIR | CIR1, CIR4, CIR7 | CIR2, CIR3, CIR5 | CIR6 |
| CD | CD5, CD7 | CD1, CD2, CD6, CD9, CD10, CD13, CD14, CD15, CD17, CD18, CD19 | CD3, CD4, CD8, CD12, CD16 |
| UC | UC3 | UC2, UC4, UC6, UC7, UC11, UC15, UC16, UC17 | UC1, UC5, UC8, UC12, UC13, UC14 |
| Confirmed E. coli-Positive Samples—n (%) * | ||
|---|---|---|
| FAECAL SAMPLES | BLOOD SAMPLES | |
| Healthy controls | 19 (86.4%) | 0 (0.0%) |
| Patients with cirrhosis | 4 (57.1%) | 0 (0.0%) |
| Crohn’s disease patients | 16 (88.9%) | 0 (0.0%) |
| Ulcerative colitis patients | 14 (93.3%) | 0 (0.0%) |
| EC Isolate | ADH-I | INV-I | REP-I |
|---|---|---|---|
| CD1 | 1.81 | 0.11 | 64.12 |
| UC5C | 0.9 | 0.17 | 188.24 |
| UC13C | 2.19 | ND | ND |
| FAECAL SAMPLES | BLOOD SAMPLES | |||
|---|---|---|---|---|
| MAP-Positive— n (%) * | Positive Sub-Culture in RAF014D | MAP-Positive— n (%) * | Positive Sub-Culture in RAF014D | |
| Healthy controls | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | ND |
| Patients with cirrhosis | 0 (0.0%) | 0 (0.0%) | 1 (14.3%) | 1 (100.0%) |
| Crohn’s disease patients | 2 (11.1%) | 1 (50.0%) | 2 (11.1%) | 1 (50.0%) |
| Ulcerative colitis patients | 1 (6.7%) | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) |
| IS900 PCR | F57 PCR | Sub-culture in RAFO14D | AFB Stain on Sub-culture | Sequenced | |||
|---|---|---|---|---|---|---|---|
| HC | Faeces | HC1 | Weak positive | Negative | NVG (12 months) | No AFB visible | No |
| CIR | Blood | CIR2 | Weak positive | Negative | NVG (12 months) | No AFB visible | No |
| CIR3 | Weak positive | Negative | NVG (12 months) | No AFB visible | No | ||
| CIR4 | Positive | Positive | Positive growth (8 months) | AFB seen | Yes | ||
| CIR6 | Low positive | Negative | Scanty growth | Suspected AFB | No | ||
| CIR7 | Weak positive | Negative | NVG (12 months) | ND | No | ||
| CD | Faeces | CD3 | Positive | Positive | Positive growth (5 months) | AFB seen | Yes |
| CD9 | Positive | Negative | NVG (12 months) | Suspected AFB | No | ||
| CD12 | Weak positive | Negative | NVG (12 months) | No AFB visible | No | ||
| CD17 | Positive | Positive | Scanty growth | Suspected AFB | No | ||
| Blood | CD3 | Positive | Positive | Positive growth (6 months) | AFB seen | Yes | |
| CD6 | Weak positive | Negative | NVG (12 months) | No AFB visible | No | ||
| CD10 | Positive | Weak positive | Scanty growth | No AFB visible | No | ||
| CD13 | Weak positive | Negative | NVG (12 months) | No AFB visible | No | ||
| UC | Faeces | UC13 | Positive | Weak positive | NVG (12 months) | No AFB visible | No |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Estevinho, M.M.; Cabeda, J.; Santiago, M.; Machado, E.; Silva, R.; Duro, M.; Pita, I.; Morais, R.; Macedo, G.; Bull, T.J.; et al. Viable Mycobacterium avium subsp. paratuberculosis Colonizes Peripheral Blood of Inflammatory Bowel Disease Patients. Microorganisms 2023, 11, 1520. https://doi.org/10.3390/microorganisms11061520
Estevinho MM, Cabeda J, Santiago M, Machado E, Silva R, Duro M, Pita I, Morais R, Macedo G, Bull TJ, et al. Viable Mycobacterium avium subsp. paratuberculosis Colonizes Peripheral Blood of Inflammatory Bowel Disease Patients. Microorganisms. 2023; 11(6):1520. https://doi.org/10.3390/microorganisms11061520
Chicago/Turabian StyleEstevinho, Maria Manuela, José Cabeda, Mafalda Santiago, Elisabete Machado, Ricardo Silva, Mary Duro, Inês Pita, Rui Morais, Guilherme Macedo, Tim J. Bull, and et al. 2023. "Viable Mycobacterium avium subsp. paratuberculosis Colonizes Peripheral Blood of Inflammatory Bowel Disease Patients" Microorganisms 11, no. 6: 1520. https://doi.org/10.3390/microorganisms11061520
APA StyleEstevinho, M. M., Cabeda, J., Santiago, M., Machado, E., Silva, R., Duro, M., Pita, I., Morais, R., Macedo, G., Bull, T. J., Magro, F., & Sarmento, A. (2023). Viable Mycobacterium avium subsp. paratuberculosis Colonizes Peripheral Blood of Inflammatory Bowel Disease Patients. Microorganisms, 11(6), 1520. https://doi.org/10.3390/microorganisms11061520