

Effects of Bacterial Lysates and Metabolites on Collagen Homeostasis in TNF-α-Challenged Human Dermal Fibroblasts


Abstract
1. Introduction
2. Materials and Methods
2.1. HDF Cell Culture
2.2. Bacterial Cell Culture and Sample Preparation
2.3. Inflammatory Challenge
2.4. Cell Viability and Confluence
2.5. ELISA
2.5.1. Measurements of Type I Pro-Collagen and MMP-1
2.5.2. Measurements of Cytokine and Growth Factor Concentrations
2.6. Statistical Analysis
3. Results
3.1. Effects of Systemic Inflammatory Challenge
3.2. Effect Differences between Strains and Species, Lysates, and Metabolites on HDF Biomarkers
3.2.1. HDF Viability and Confluence
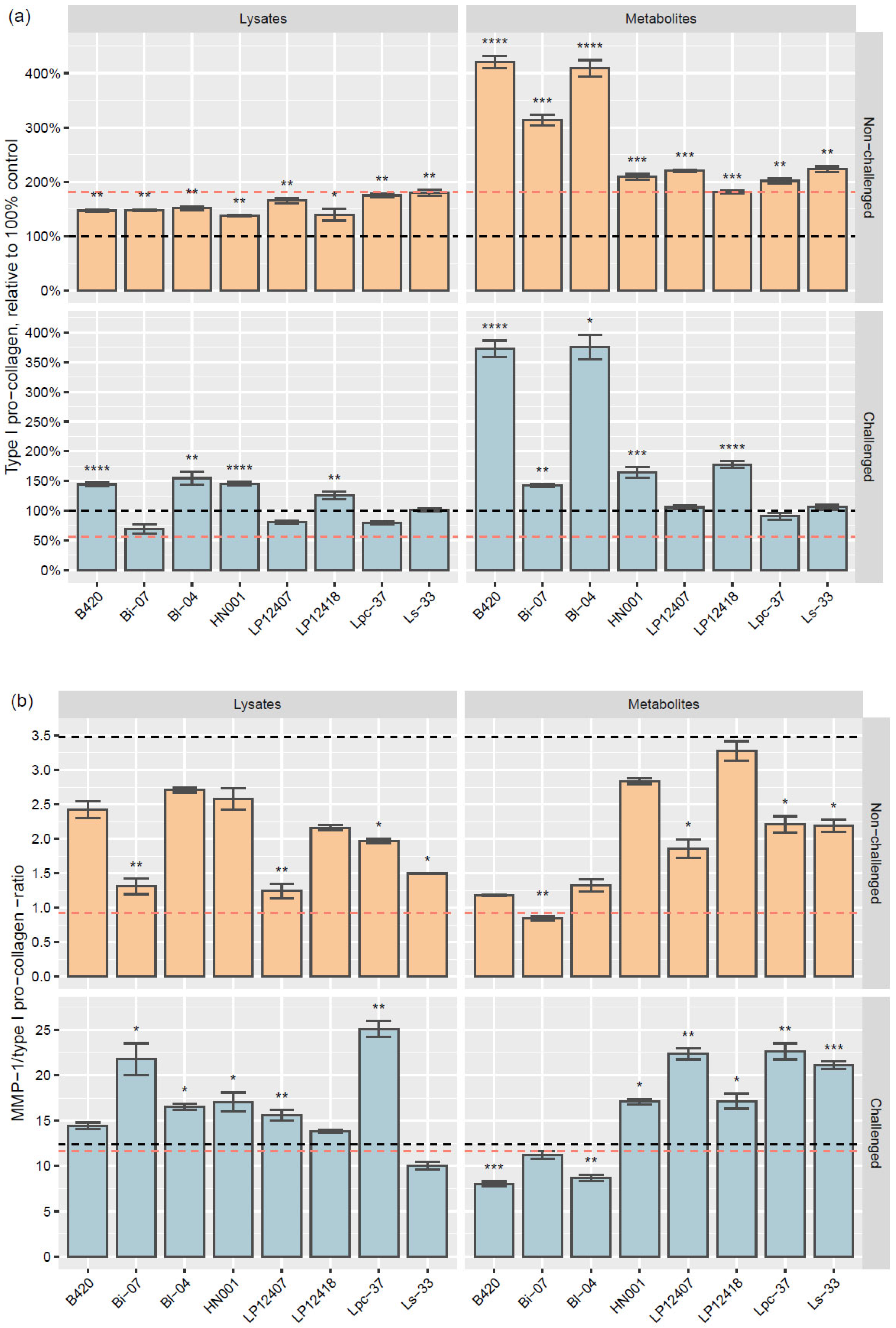
3.2.2. Type I Pro-Collagen and MMP-1/Type I Pro-Collagen Ratio
3.2.3. Cytokine Levels
3.2.4. Growth Factors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cole, M.A.; Quan, T.; Voorhees, J.J.; Fisher, G.J. Extracellular matrix regulation of fibroblast function: Redefining our perspective on skin aging. J. Cell Commun. Signal. 2018, 12, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Zalewska, A.; Głowacka, E.; Wyczółkowska, J.; Tchórzewski, H.; Narbutt, J.; Sysa-Jedrzejowska, A. Interleukin 6 and 8 levels in plasma and fibroblast cultures in psoriasis. Mediat. Inflamm. 2006, 2006, 81767. [Google Scholar] [CrossRef] [PubMed]
- Strickland, I.; Rhodes, L.E.; Flanagan, B.F.; Friedmann, P.S. TNF-alpha and IL-8 are upregulated in the epidermis of normal human skin after UVB exposure: Correlation with neutrophil accumulation and E-selectin expression. J. Investig. Dermatol. 1997, 108, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Freund, A.; Orjalo, A.V.; Desprez, P.Y.; Campisi, J. Inflammatory networks during cellular senescence: Causes and consequences. Trends Mol. Med. 2010, 16, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Pilkington, S.M.; Bulfone-Paus, S.; Griffiths, C.E.M.; Watson, R.E.B. Inflammaging and the Skin. J. Investig. Dermatol. 2021, 141, 1087–1095. [Google Scholar] [CrossRef]
- Mavrogonatou, E.; Konstantinou, A.; Kletsas, D. Long-term exposure to TNF-α leads human skin fibroblasts to a p38 MAPK- and ROS-mediated premature senescence. Biogerontology 2018, 19, 237–249. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef]
- Szántó, M.; Dózsa, A.; Antal, D.; Szabó, K.; Kemény, L.; Bai, P. Targeting the gut-skin axis—Probiotics as new tools for skin disorder management? Exp. Dermatol. 2019, 28, 1210–1218. [Google Scholar] [CrossRef]
- Apajalahti, J.H.; Kettunen, H.; Kettunen, A.; Holben, W.E.; Nurminen, P.H.; Rautonen, N.; Mutanen, M. Culture-independent microbial community analysis reveals that inulin in the diet primarily affects previously unknown bacteria in the mouse cecum. Appl. Environ. Microbiol. 2002, 68, 4986–4995. [Google Scholar] [CrossRef][Green Version]
- Putaala, H.; Tiihonen, K.; Ouwehand, A.C.; Rautonen, N. Probiotics modulate tight junction integrity and expression of junctional proteins in cultured normal human epidermal keratinocytes. Int. J. Probiotics Prebiotics 2012, 7, 81–90. [Google Scholar]
- Hsu, M.-F.; Chiang, B.-H. Effect of Bacillus subtilis natto–fermented Radix astragali on collagen production in human skin fibroblasts. Process Biochem. 2009, 44, 83–90. [Google Scholar] [CrossRef]
- Südel, K.M.; Venzke, K.; Mielke, H.; Breitenbach, U.; Mundt, C.; Jaspers, S.; Koop, U.; Sauermann, K.; Knussman-Hartig, E.; Moll, I.; et al. Novel aspects of intrinsic and extrinsic aging of human skin: Beneficial effects of soy extract. Photochem. Photobiol. 2005, 81, 581–587. [Google Scholar] [CrossRef]
- Varani, J.; Dame, M.K.; Rittie, L.; Fligiel, S.E.; Kang, S.; Fisher, G.J.; Voorhees, J.J. Decreased collagen production in chronologically aged skin: Roles of age-dependent alteration in fibroblast function and defective mechanical stimulation. Am. J. Pathol. 2006, 168, 1861–1868. [Google Scholar] [CrossRef]
- Papakonstantinou, E.; Roth, M.; Karakiulakis, G. Hyaluronic acid: A key molecule in skin aging. Dermatoendocrinology 2012, 4, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Rohani, M.G.; Parks, W.C. Matrix remodeling by MMPs during wound repair. Matrix Biol. 2015, 44–46, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Zorina, A.; Zorin, V.; Kudlay, D.; Kopnin, P. Molecular Mechanisms of Changes in Homeostasis of the Dermal Extracellular Matrix: Both Involutional and Mediated by Ultraviolet Radiation. Int. J. Mol. Sci. 2022, 23, 6655. [Google Scholar] [CrossRef]
- Xia, S.; Zhang, X.; Zheng, S.; Khanabdali, R.; Kalionis, B.; Wu, J.; Wan, W.; Tai, X. An Update on Inflamm-Aging: Mechanisms, Prevention, and Treatment. J. Immunol. Res. 2016, 2016, 8426874. [Google Scholar] [CrossRef]
- Zhuang, Y.; Lyga, J. Inflammaging in skin and other tissues—The roles of complement system and macrophage. Inflamm. Allergy Drug Targets 2014, 13, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Jarrold, B.B.; Tan, C.Y.R.; Ho, C.Y.; Soon, A.L.; Lam, T.T.; Yang, X.; Nguyen, C.; Guo, W.; Chew, Y.C.; DeAngelis, Y.M.; et al. Early onset of senescence and imbalanced epidermal homeostasis across the decades in photoexposed human skin: Fingerprints of inflammaging. Exp. Dermatol. 2022, 31, 1748–1760. [Google Scholar] [CrossRef]
- Bashir, M.M.; Sharma, M.R.; Werth, V.P. TNF-alpha production in the skin. Arch. Dermatol. Res. 2009, 301, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Ågren, M.S.; Schnabel, R.; Christensen, L.H.; Mirastschijski, U. Tumor necrosis factor-α-accelerated degradation of type I collagen in human skin is associated with elevated matrix metalloproteinase (MMP)-1 and MMP-3 ex vivo. Eur. J. Cell Biol. 2015, 94, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Choi, Y.J.; Lee, S.; Kang, K.S.; Jang, T.S.; Kim, K.H. Protective Effects of Withagenin A Diglucoside from Indian Ginseng (Withania somnifera) against Human Dermal Fibroblast Damaged by TNF-α Stimulation. Antioxidants 2022, 11, 2248. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Yu, J.S.; Phung, H.M.; Lee, J.G.; Kim, K.H.; Kang, K.S. Potential Anti-Skin Aging Effect of (-)-Catechin Isolated from the Root Bark of Ulmus davidiana var. japonica in Tumor Necrosis Factor-α-Stimulated Normal Human Dermal Fibroblasts. Antioxidants 2020, 9, 981. [Google Scholar]
- Lee, S.; Jang, T.; Kim, K.H.; Kang, K.S. Improvement of Damage in Human Dermal Fibroblasts by 3,5,7-Trimethoxyflavone from Black Ginger (Kaempferia parviflora). Antioxidants 2022, 11, 425. [Google Scholar] [CrossRef]
- Bae, J.-Y.; Choi, J.-S.; Choi, Y.-J.; Shin, S.-Y.; Kang, S.-W.; Han, S.J.; Kang, Y.-H. (−)Epigallocatechin gallate hampers collagen destruction and collagenase activation in ultraviolet-B-irradiated human dermal fibroblasts: Involvement of mitogen-activated protein kinase. Food Chem. Toxicol. 2008, 46, 1298–1307. [Google Scholar] [CrossRef]
- Lim, H.Y.; Jeong, D.; Park, S.H.; Shin, K.K.; Hong, Y.H.; Kim, E.; Yu, Y.G.; Kim, T.R.; Kim, H.; Lee, J.; et al. Antiwrinkle and Antimelanogenesis Effects of Tyndallized Lactobacillus acidophilus KCCM12625P. Int. J. Mol. Sci. 2020, 21, 1620. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.M.; Lee, D.E.; Park, S.D.; Kim, Y.T.; Kim, Y.J.; Jeong, J.W.; Jang, S.S.; Ahn, Y.T.; Sim, J.H.; Huh, C.S.; et al. Oral administration of Lactobacillus plantarum HY7714 protects hairless mouse against ultraviolet B-induced photoaging. J. Microbiol. Biotechnol. 2014, 24, 1583–1591. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, X.; Song, Y.; Zheng, B.; Wen, Z.; Gong, M.; Meng, L. Heat-Killed Lacticaseibacillus paracasei Ameliorated UVB-Induced Oxidative Damage and Photoaging and Its Underlying Mechanisms. Antioxidants 2022, 11, 1875. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, J.; Ma, M.; Zhao, Y.; Song, Y.; Zheng, B.; Wen, Z.; Gong, M.; Meng, L. Heat-Killed Lactobacillus rhamnosus ATCC 7469 Improved UVB-Induced Photoaging via Antiwrinkle and Antimelanogenesis Impacts. Photochem. Photobiol. 2023. [Google Scholar] [CrossRef]
- You, G.E.; Jung, B.J.; Kim, H.R.; Kim, H.G.; Kim, T.R.; Chung, D.K. Lactobacillus sakei lipoteichoic acid inhibits MMP-1 induced by UVA in normal dermal fibroblasts of human. J. Microbiol. Biotechnol. 2013, 23, 1357–1364. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Kim, H.J.; Kim, S.A.; Park, S.D.; Shim, J.J.; Lee, J.L. Exopolysaccharide from Lactobacillus plantarum HY7714 Protects against Skin Aging through Skin-Gut Axis Communication. Molecules 2021, 26, 1651. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Park, J.Y.; Kim, Y.; Kang, C.H. Protective Effect of Bifidobacterium animalis subs. lactis MG741 as Probiotics against UVB-Exposed Fibroblasts and Hairless Mice. Microorganisms 2022, 10, 2343. [Google Scholar] [CrossRef]
- Park, J.Y.; Lee, J.Y.; Kim, Y.; Kang, C.H. Lactic Acid Bacteria Improve the Photoprotective Effect via MAPK/AP-1/MMP Signaling Pathway on Skin Fibroblasts. Microorganisms 2022, 10, 2481. [Google Scholar] [CrossRef] [PubMed]
- Jwo, J.Y.; Chang, Y.T.; Huang, Y.C. Effects of probiotics supplementation on skin photoaging and skin barrier function: A systematic review and meta-analysis. Photodermatol. Photoimmunol. Photomed. 2022. [Google Scholar] [CrossRef]
- Topman, G.; Sharabani-Yosef, O.; Gefen, A. A method for quick, low-cost automated confluency measurements. Microsc. Microanal. 2011, 17, 915–922. [Google Scholar] [CrossRef]
- Varani, J.; Schuger, L.; Dame, M.K.; Leonard, C.; Fligiel, S.E.; Kang, S.; Fisher, G.J.; Voorhees, J.J. Reduced fibroblast interaction with intact collagen as a mechanism for depressed collagen synthesis in photodamaged skin. J. Investig. Dermatol. 2004, 122, 1471–1479. [Google Scholar] [CrossRef]
- Lee, Y.I.; Choi, S.; Roh, W.S.; Lee, J.H.; Kim, T.G. Cellular Senescence and Inflammaging in the Skin Microenvironment. Int. J. Mol. Sci. 2021, 22, 3849. [Google Scholar] [CrossRef]
- Shams, F.; Moravvej, H.; Hosseinzadeh, S.; Mostafavi, E.; Bayat, H.; Kazemi, B.; Bandehpour, M.; Rostami, E.; Rahimpour, A.; Moosavian, H. Overexpression of VEGF in dermal fibroblast cells accelerates the angiogenesis and wound healing function: In vitro and in vivo studies. Sci. Rep. 2022, 12, 18529. [Google Scholar] [CrossRef]
- Spanholtz, T.A.; Theodorou, P.; Holzbach, T.; Wutzler, S.; Giunta, R.E.; Machens, H.G. Vascular endothelial growth factor (VEGF165) plus basic fibroblast growth factor (bFGF) producing cells induce a mature and stable vascular network—a future therapy for ischemically challenged tissue. J. Surg. Res. 2011, 171, 329–338. [Google Scholar] [CrossRef]
- Werner, S.; Peters, K.G.; Longaker, M.T.; Fuller-Pace, F.; Banda, M.J.; Williams, L.T. Large induction of keratinocyte growth factor expression in the dermis during wound healing. Proc. Natl. Acad. Sci. USA 1992, 89, 6896–6900. [Google Scholar] [CrossRef] [PubMed]
- Belvedere, R.; Novizio, N.; Morello, S.; Petrella, A. The combination of mesoglycan and VEGF promotes skin wound repair by enhancing the activation of endothelial cells and fibroblasts and their cross-talk. Sci. Rep. 2022, 12, 11041. [Google Scholar] [CrossRef] [PubMed]
- Varga, J.; Rosenbloom, J.; Jimenez, S.A. Transforming growth factor beta (TGF beta) causes a persistent increase in steady-state amounts of type I and type III collagen and fibronectin mRNAs in normal human dermal fibroblasts. Biochem. J. 1987, 247, 597–604. [Google Scholar] [CrossRef] [PubMed]
- de Araújo, R.; Lôbo, M.; Trindade, K.; Silva, D.F.; Pereira, N. Fibroblast Growth Factors: A Controlling Mechanism of Skin Aging. Skin Pharmacol. Physiol. 2019, 32, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Huggenberger, R.; Detmar, M. The cutaneous vascular system in chronic skin inflammation. J. Investig. Dermatol. Symp. Proc. 2011, 15, 24–32. [Google Scholar] [CrossRef]
- Yano, K.; Oura, H.; Detmar, M. Targeted overexpression of the angiogenesis inhibitor thrombospondin-1 in the epidermis of transgenic mice prevents ultraviolet-B-induced angiogenesis and cutaneous photo-damage. J. Investig. Dermatol. 2002, 118, 800–805. [Google Scholar] [CrossRef]
- Barrientos, S.; Stojadinovic, O.; Golinko, M.S.; Brem, H.; Tomic-Canic, M. Growth factors and cytokines in wound healing. Wound Repair Regen. 2008, 16, 585–601. [Google Scholar] [CrossRef]
- McFarland, L.V.; Evans, C.T.; Goldstein, E.J.C. Strain-Specificity and Disease-Specificity of Probiotic Efficacy: A Systematic Review and Meta-Analysis. Front. Med. 2018, 5, 124. [Google Scholar] [CrossRef]
- Sengupta, R.; Altermann, E.; Anderson, R.C.; McNabb, W.C.; Moughan, P.J.; Roy, N.C. The role of cell surface architecture of lactobacilli in host-microbe interactions in the gastrointestinal tract. Mediators Inflamm. 2013, 2013, 237921. [Google Scholar] [CrossRef]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Mechanisms of Action of Probiotics. Adv. Nutr. 2019, 10, S49–S66. [Google Scholar] [CrossRef]
- Lew, L.-C.; Gan, C.-Y.; Liong, M.-T. Dermal bioactives from lactobacilli and bifidobacteria. Ann. Microbiol. 2013, 63, 1047–1055. [Google Scholar] [CrossRef]
- Liu, Q.; Yu, Z.; Tian, F.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Surface components and metabolites of probiotics for regulation of intestinal epithelial barrier. Microb Cell Fact 2020, 19, 23. [Google Scholar] [CrossRef]
- Correa-Gallegos, D.; Rinkevich, Y. Cutting into wound repair. FEBS J. 2022, 289, 5034–5048. [Google Scholar] [CrossRef] [PubMed]
- Davidson, S.; Coles, M.; Thomas, T.; Kollias, G.; Ludewig, B.; Turley, S.; Brenner, M.; Buckley, C.D. Fibroblasts as immune regulators in infection, inflammation and cancer. Nat. Rev. Immunol. 2021, 21, 704–717. [Google Scholar] [CrossRef]
- Huuskonen, L.; Lyra, A.; Lee, E.; Ryu, J.; Jeong, H.; Baek, J.; Seo, Y.; Shin, M.; Tiihonen, K.; Pesonen, T.; et al. Effects of Bifidobacterium animalis subsp. lactis Bl-04 on Skin Wrinkles and Dryness: A Randomized, Triple-Blinded, Placebo-Controlled Clinical Trial. Dermato 2022, 2, 5. [Google Scholar] [CrossRef]
- Stahl, B.; Barrangou, R. Complete genome sequences of probiotic strains Bifidobacterium animalis subsp. lactis B420 and Bi-07. J. Bacteriol. 2012, 194, 4131–4132. [Google Scholar] [CrossRef] [PubMed]
- Barrangou, R.; Briczinski, E.P.; Traeger, L.L.; Loquasto, J.R.; Richards, M.; Horvath, P.; Coûté-Monvoisin, A.C.; Leyer, G.; Rendulic, S.; Steele, J.L.; et al. Comparison of the complete genome sequences of Bifidobacterium animalis subsp. lactis DSM 10140 and Bl-04. J. Bacteriol. 2009, 191, 4144–4151. [Google Scholar] [CrossRef]
- Loquasto, J.R.; Barrangou, R.; Dudley, E.G.; Stahl, B.; Chen, C.; Roberts, R.F. Bifidobacterium animalis subsp. lactis ATCC 27673 is a genomically unique strain within its conserved subspecies. Appl. Environ. Microbiol. 2013, 79, 6903–6910. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameter | Effect of TNF-α Challenge on Parameter |
|---|---|
| Viability | + |
| Confluence | + |
| Type I pro-collagen | − |
| MMP-1/type I pro-collagen ratio | + |
| FGFb | − |
| IL-10 | + |
| IL-1α | + |
| IL-1β | + |
| IL-6 | + |
| IL-8 | + |
| KGF | + |
| TGF-β1 | − |
| TNF-α | + |
| VEGF | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huuskonen, L.; Anglenius, H.; Ahonen, I.; Tiihonen, K. Effects of Bacterial Lysates and Metabolites on Collagen Homeostasis in TNF-α-Challenged Human Dermal Fibroblasts. Microorganisms 2023, 11, 1465. https://doi.org/10.3390/microorganisms11061465
Huuskonen L, Anglenius H, Ahonen I, Tiihonen K. Effects of Bacterial Lysates and Metabolites on Collagen Homeostasis in TNF-α-Challenged Human Dermal Fibroblasts. Microorganisms. 2023; 11(6):1465. https://doi.org/10.3390/microorganisms11061465
Chicago/Turabian StyleHuuskonen, Laura, Heli Anglenius, Ilmari Ahonen, and Kirsti Tiihonen. 2023. "Effects of Bacterial Lysates and Metabolites on Collagen Homeostasis in TNF-α-Challenged Human Dermal Fibroblasts" Microorganisms 11, no. 6: 1465. https://doi.org/10.3390/microorganisms11061465
APA StyleHuuskonen, L., Anglenius, H., Ahonen, I., & Tiihonen, K. (2023). Effects of Bacterial Lysates and Metabolites on Collagen Homeostasis in TNF-α-Challenged Human Dermal Fibroblasts. Microorganisms, 11(6), 1465. https://doi.org/10.3390/microorganisms11061465