
Rarely Encountered Gram-Negative Rods and Lung Transplant Recipients: A Narrative Review
 ,
,  and
and 
Abstract
1. Introduction
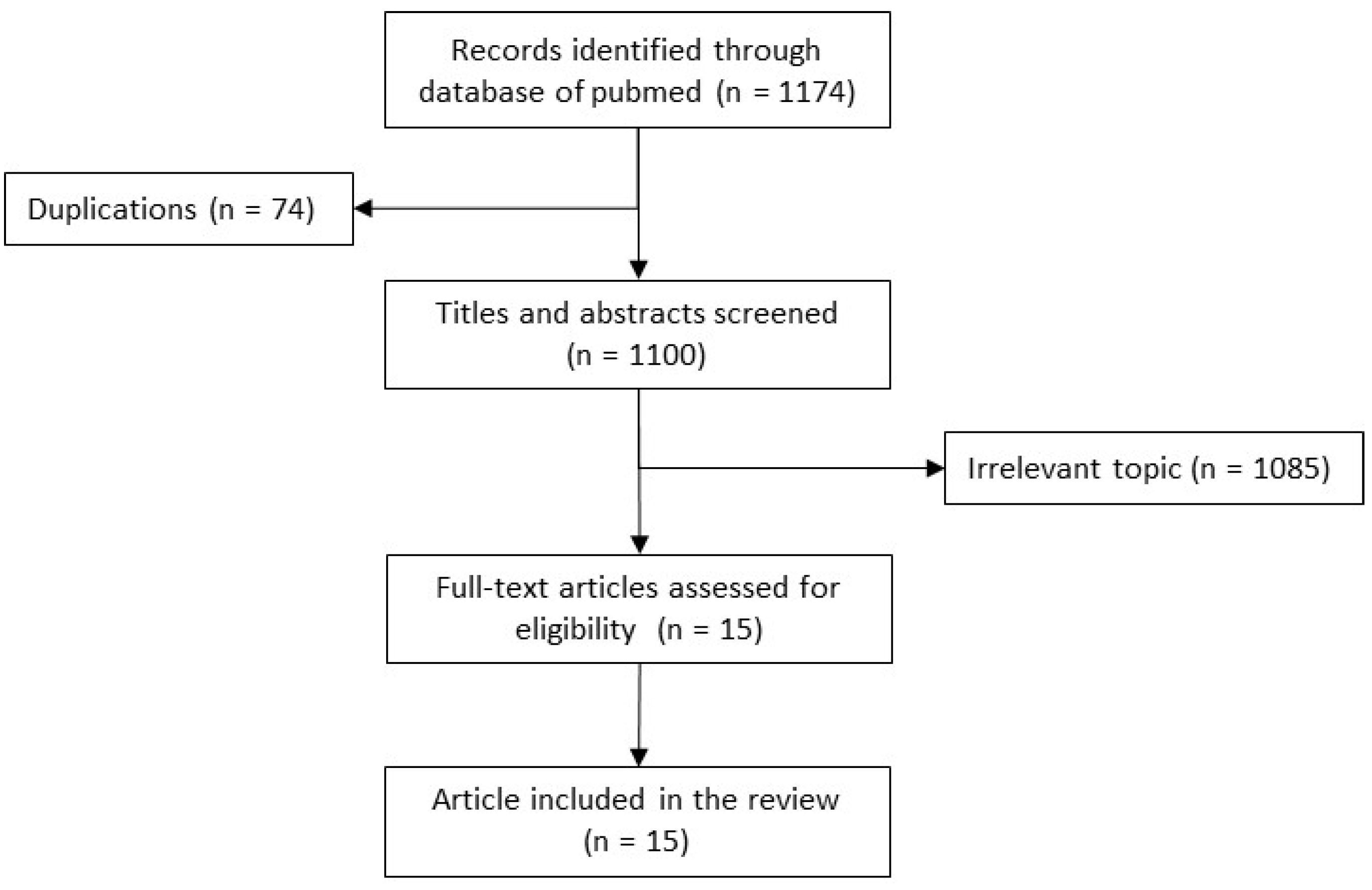
2. Methods of the Narrative Review
3. Review of Cases (Table 2)
3.1. Acetobacter indonesiensis
3.2. Chryseobacterium spp.
3.3. Elizabethkinga spp.
3.4. Inquilinus limosus
3.5. Pandoraea spp.
3.6. Bordetella spp.

{kind=link}
| No. | Age | Sex | Reason for Lung Transplantation | Infection | Isolation before Transplant | Species | Other Associated Pathogens | Past History of Antimicrobial Treatment | Medical History | Outcome | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 31 | M | Cystic fibrosis | Early post-operative pneumonia/Colonization | No | A. indonesiensis | P. aeruginosa S. aureus | Peri-operative prophylaxis with colistin, tobramycin, ceftazidime, and linezolid |
| Recovery | [21] |
| 2 | 51 | F |
| Early post-operative Pneumonia/Colonization | No | A. indonesiensis | n.a. * | Post-operative prophylaxis cotrimoxazole, vancomycin, piperacillin/tazobactam |
| Recovery | [4] |
| 3 | n.a. | n.a. | n.a. | Pneumonia 1 to 6 months after transplant | n.a. | Chryseobacterium spp. | n.a. | n.a. | n.a. | n.a. | [6] |
| 4 | 26 | M | Cystic fibrosis | Septic shock 5 years after transplant | No | Elizabethkinga spp. | P. aeruginosa | n.a. |
| Death | [7] |
| 5 | 22 | F | Cystic fibrosis | Early post-operative pulmonary infiltrate (week 1) | Yes | I. limosus | n.a. | Peri-operative prophylaxis with imipenem, tobramycin, ceftazidime, and aerosolized colistin |
| Recovery | [8] |
| 6 | 31 | M | Cystic fibrosis | Early bacteraemic lung empyema on postoperative day 38 and month 8 | Yes | I. limosus | no | Peri-operative prophylaxis with piperacillin-tazobactam, tobramycin, cotrimoxazole, nebulized amphotericin, azithromycin |
| Recovery | [9] |
| 7 | 45 | F | Cystic fibrosis | Bacteremia 8 years after lung transplantation | Yes | I. limosus | No | n.a. |
| Recovery | [10] |
| 8 | 30 | F | Cystic fibrosis | Early post-operative pleural effusion | Yes | P. apista | P. aeruginosa | n.a. |
| Recovery | [14] |
| 9 | 36 | M | Cystic fibrosis | Colonization | Yes | P. apista | n.a. | n.a. |
| Recovery | [14] |
| 10 | 21 | F | Csytic fibrosis | Early post-operative septic choc | Yes | P. nosoerga | No | Peri-operative prophylaxis piperacillin/tazobactam, tigecyclin |
| Death | [15] |
| 11 | 30 | M | End-stage pulmonary sarcoidosis complicated by nocardiosis and mycetomas. Prednisone 50 mg daily | Septic shock | No | P. pnomenusa | n.a. | Peri-operative prophylaxis Ceftazidime, vancomycin. |
| Death | [11] |
| 12 | n.a. | n.a. | Cystic fibrosis | Colonization | n.a. | P. pulmonicola | n.a. | n.a. | n.a. | Recovery | [13] |
| 13 | 30 | F | Cystic fibrosis | Colonization | Yes | P. pulmonicola | P. aeruginosa | n.a. |
| Death | [12] |
| 14 | 42 | M | Cystic fibrosis | ARDS 5 years post-transplant | no | B. pertussis | No | n.a. |
| Recovery | [18] |
| 15 | 10 | F | Cystic fibrosis | Late post-transplant pneumonia (1-year) | no | B. bronchiseptica | P. aeruginosa, A. fumigatus, K. pneumoniae, S. maltophilia. | n.a. |
| Death | [19] |
| 16 | 15 | M | Cystic fibrosis | Bronchitis 3 months post-transplant | no | B. bronchiseptica | B. cepacia | n.a. |
| Recovery | [19] |
| 17 | 51 | M | MDA-5 associated clinically amyopathic dermatomyositis. | Bronchitis 4 months post-transplant | no | B. bronchiseptica | No | n.a. |
| Recovery | [20] |
4. Issues Raised by Opportunistic GNR
4.1. Detection and Identification
4.2. Antimicrobial Resistance
4.3. Pathogenesis
4.4. Cross-Transmission and Prevention
5. Issues Raised by Bordetella spp.
6. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Waller, K.M.J.; De La Mata, N.L.; Wyburn, K.R.; Hedley, J.A.; Rosales, B.M.; Kelly, P.J.; Ramachandran, V.; Shah, K.K.; Morton, R.L.; Rawlinson, W.D.; et al. Notifiable Infectious Diseases Among Organ Transplant Recipients: A Data-Linked Cohort Study, 2000–2015. Open Forum Infect. Dis. 2022, 9, ofac337. [Google Scholar] [CrossRef] [PubMed]
- Konishi, Y.; Miyoshi, K.; Kurosaki, T.; Otani, S.; Sugimoto, S.; Yamane, M.; Oto, T.; Toyooka, S. Airway bacteria of the recipient but not the donor are relevant to post-lung transplant pneumonia. Gen. Thorac. Cardiovasc. Surg. 2020, 68, 833–840. [Google Scholar] [CrossRef] [PubMed]
- National Library of Medicine. Taxonomy Browser n.d. Available online: https://www.ncbi.nlm.nih.gov/taxonomy (accessed on 8 October 2022).
- Basu, S.S.; Delaney, M.L.; Li, N.; Onderdonk, A.B.; Bry, L. Acetobacter indonesiensis Pneumonia after Lung Transplantation. Emerg. Infect. Dis. 2018, 24, 598–599. [Google Scholar] [CrossRef]
- Bittar, F.; Reynaud-Gaubert, M.; Thomas, P.; Boniface, S.; Raoult, D.; Rolain, J.-M. Acetobacter indonesiensis pneumonia after lung transplant. Emerg. Infect. Dis. 2008, 14, 997–998. [Google Scholar] [CrossRef]
- Bae, M.; Lee, S.-O.; Jo, K.-W.; Choi, S.; Lee, J.; Chae, E.J.; Do, K.-H.; Choi, D.-K.; Choi, I.-C.; Hong, S.-B.; et al. Infections in Lung Transplant Recipients during and after Prophylaxis. Infect. Chemother. 2020, 52, 600–610. [Google Scholar] [CrossRef]
- Ramanan, P.; Razonable, R. Elizabethkingia species sepsis after lung transplantation: Case report and literature review. Transpl. Infect. Dis. 2013, 15, E229–E234. [Google Scholar] [CrossRef] [PubMed]
- Pitulle, C.; Citron, D.M.; Bochner, B.; Barbers, R.; Appleman, M.D. Novel Bacterium Isolated from a Lung Transplant Patient with Cystic Fibrosis. J. Clin. Microbiol. 1999, 37, 3851–3855. [Google Scholar] [CrossRef] [PubMed]
- Goeman, E.; Shivam, A.; Downton, T.; Glanville, A.R. Bacteremic Inquilinus limosus empyema in an Australian lung transplant patient with cystic fibrosis. J. Hearth Lung Transplant. 2015, 34, 1220–1223. [Google Scholar] [CrossRef]
- Farfour, E.; Zrounba, M.; Roux, A.; Revillet, H.; Vallée, A.; Vasse, M. Inquilinus limosus Bacteremia in Lung Transplant Recipient after SARS-CoV-2 Infection. Emerg. Infect. Dis. 2023, 29, 642–644. [Google Scholar] [CrossRef]
- Stryjewski, M.E.; LiPuma, J.J.; Messier, J.R.H.; Reller, L.B.; Alexander, B.D. Sepsis, multiple organ failure, and death due to Pandoraea pnomenusa infection after lung transplantation. J. Clin. Microbiol. 2003, 41, 2255–2257. [Google Scholar] [CrossRef]
- Kokcha, S.; Bittar, F.; Reynaud-Gaubert, M.; Mely, L.; Gomez, C.; Gaubert, J.-Y.; Thomas, P.; Rolain, J.-M. Pandoraea pulmonicola chronic colonization in a cystic fibrosis patient, France. New Microbes New Infect. 2013, 1, 27–29. [Google Scholar] [CrossRef] [PubMed]
- Caraher, E.; Collins, J.; Herbert, G.; Murphy, P.G.; Gallagher, C.G.; Crowe, M.J.; Callaghan, M.; McClean, S. Evaluation of in vitro virulence characteristics of the genus Pandoraea in lung epithelial cells. J. Med. Microbiol. 2008, 57, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, R.M.; LiPuma, J.J.; Rosenbluth, D.B.; Dunne, W.M. Chronic colonization with Pandoraea apista in cystic fibrosis patients determined by repetitive-element-sequence PCR. J. Clin. Microbiol. 2006, 44, 833–836. [Google Scholar] [CrossRef]
- Peyclit, L.; Baron, S.A.; Reynaud-Gaubert, M.; Cassir, N.; Rolain, J.M. Fatal Pandoraea nosoerga infection after combined liver-lung transplantation for cystic fibrosis: A recontamination by the pre-transplantation strain n.d. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 2403–2406. [Google Scholar] [CrossRef]
- Pimentel, J.D.; MacLeod, C. Misidentification of Pandoraea sputorum isolated from sputum of a patient with cystic fibrosis and review of Pandoraea species infections in transplant patients. J. Clin. Microbiol. 2008, 46, 3165–3168. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, I.M.; Johansen, H.K.; Frederiksen, B.; Pressler, T.; Hansen, A.; Vandamme, P.; Høiby, N.; Koch, C. Epidemic Spread of Pandoraea apista, a New Pathogen Causing Severe Lung Disease in Cystic Fibrosis Patients. Pediatr. Pulmonol. 2003, 36, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Panchabhai, T.S.; Bandyopadhyay, D.; Kapoor, A.; Akindipe, O.; Lane, C.; Krishnan, S. Acute ischemic optic neuropathy with extended prone position ventilation in a lung transplant recipient. Int. J. Crit. Illn. Inj. Sci. 2016, 6, 45–47. [Google Scholar] [CrossRef]
- Ner, Z.; Ross, L.A.; Horn, M.V.; Keens, T.G.; MacLaughlin, E.F.; Starnes, V.A.; Woo, M.S. Bordetella bronchiseptica infection in pediatric lung transplant recipients. Pediatr. Transplant. 2003, 7, 413–417. [Google Scholar] [CrossRef]
- Deitchman, A.R.; Kalchiem-Dekel, O.; Todd, N.; Reed, R.M. Rapidly progressive interstitial lung disease due to anti-melanoma differentiation associated protein-5 requiring a bilateral lung transplant, and complicated by kennel cough. Respir. Med. Case Rep. 2019, 28, 100886. [Google Scholar] [CrossRef]
- Bittar, F.; Leydier, A.; Bosdure, E.; Toro, A.; Reynaud-Gaubert, M.; Boniface, S.; Stremler, N.; Dubus, J.-C.; Sarles, J.; Raoult, D.; et al. Inquilinus limosus and cystic fibrosis. Emerg. Infect. Dis. 2008, 14, 993–995. [Google Scholar] [CrossRef]
- Hogardt, M.; Ulrich, J.; Riehn-Kopp, H.; Tümmler, B. EuroCareCF quality assessment of diagnostic microbiology of cystic fibrosis isolates. J. Clin. Microbiol. 2009, 47, 3435–3438. [Google Scholar] [CrossRef] [PubMed]
- Farfour, E.; Botterel, F.; Pozzetto, B. Infections Broncho-Pulmonaires (Hors Tuberculose et Mucoviscidose); REMIC 7; Société Française de Microbiologie: Paris, France, 2022; pp. 1–16. [Google Scholar]
- Schmoldt, S.; Latzin, P.; Heesemann, J.; Griese, M.; Imhof, A.; Hogardt, M. Clonal analysis of Inquilinus limosus isolates from six cystic fibrosis patients and specific serum antibody response. J. Med. Microbiol. 2006, 55, 1425–1433. [Google Scholar] [CrossRef] [PubMed]
- Chiron, R.; Marchandin, H.; Counil, F.; Jumas-Bilak, E.; Freydière, A.-M.; Bellon, G.; Husson, M.-O.; Turck, D.; Brémont, F.; Chabanon, G.; et al. Clinical and microbiological features of Inquilinus sp. isolates from five patients with cystic fibrosis. J. Clin. Microbiol. 2005, 43, 3938–3943. [Google Scholar] [CrossRef] [PubMed]
- McHugh, K.E.; Rhoads, D.D.; Wilson, D.A.; Highland, K.B.; Richter, S.S.; Procop, G.W. Inquilinus limosus in pulmonary disease: Case report and review of the literature. Diagn. Microbiol. Infect. Dis. 2016, 86, 446–449. [Google Scholar] [CrossRef] [PubMed]
- Adley, C.; Saieb, F. Comparison of bioMérieux API 20NE and Remel RapID NF Plus, identification systems of type strains of Ralstonia pickettii. Lett. Appl. Microbiol. 2005, 41, 136–140. [Google Scholar] [CrossRef]
- Ryan, M.P.; Pembroke, J.T.; Adley, C.C. Genotypic and phenotypic diversity of Ralstonia pickettii and Ralstonia insidiosa isolates from clinical and environmental sources including High-purity Water. Diversity in Ralstonia pickettii. BMC Microbiol. 2011, 11, 194. [Google Scholar] [CrossRef]
- Wellinghausen, N.; Köthe, J.; Wirths, B.; Sigge, A.; Poppert, S.; Johnson, J.R.; Scheutz, F.; Ulleryd, P.; Kuskowski, M.A.; O’Bryan, T.T.; et al. Superiority of molecular techniques for identification of gram-negative, oxidase-positive rods, including morphologically nontypical Pseudomonas aeruginosa, from patients with cystic fibrosis. J. Clin. Microbiol. 2005, 43, 3895–3900. [Google Scholar] [CrossRef]
- Salvador-García, C.; Yagüe-Guirao, G.; Pastor-Vivero, M.D.; Sáez-Nieto, J.A. Chronic colonization of Inquilinus limosus in a patient with cystic fibrosis: First report in Spain. Enferm. Infecc. Microbiol. Clin. 2013, 31, 414–415. [Google Scholar] [CrossRef]
- Bittar, F.; Richet, H.; Dubus, J.-C.; Reynaud-Gaubert, M.; Stremler, N.; Sarles, J.; Raoult, D.; Rolain, J.-M. Molecular detection of multiple emerging pathogens in sputa from cystic fibrosis patients. PLoS ONE 2008, 3, e2908. [Google Scholar] [CrossRef]
- Kidd, T.J.; Ramsay, K.A.; Hu, H.; Bye, P.T.P.; Elkins, M.R.; Grimwood, K.; Harbour, C.; Marks, G.B.; Nissen, M.D.; Robinson, P.J.; et al. Low rates of Pseudomonas aeruginosa misidentification in isolates from cystic fibrosis patients. J. Clin. Microbiol. 2009, 47, 1503–1509. [Google Scholar] [CrossRef]
- Brizendine, K.D.; Baddley, J.W.; Pappas, P.G.; Leon, K.J.; Rodriguez, J.M. Fatal Burkholderia gladioli infection misidentified as Empedobacter brevis in a lung transplant recipient with cystic fibrosis. Transpl. Infect. Dis. 2012, 14, E13–E18. [Google Scholar] [CrossRef]
- Buechler, C.; Neidhöfer, C.; Hornung, T.; Neuenhoff, M.; Parčina, M. Detection and characterization of clinical bordetella trematum isolates from chronic wounds. Pathogens 2021, 10, 966. [Google Scholar] [CrossRef] [PubMed]
- Bosshard, P.P.; Zbinden, R.; Abels, S.; Böddinghaus, B.; Altwegg, M.; Böttger, E.C. 16S rRNA gene sequencing versus the API 20 NE system and the VITEK 2 ID-GNB card for identification of nonfermenting Gram-negative bacteria in the clinical laboratory. J. Clin. Microbiol. 2006, 44, 1359–1366. [Google Scholar] [CrossRef] [PubMed]
- Souza, H.A.P.H.D.M.D.; Dalla-Costa, L.M.; Vicenzi, F.J.; De Souza, D.C.; Riedi, C.A.; Filho, N.A.R.; Pillonetto, M. MALDI-TOF: A useful tool for laboratory identification of uncommon glucose non-fermenting gram-negative bacteria associated with cystic fibrosis. J. Med. Microbiol. 2014, 63, 1148–1153. [Google Scholar] [CrossRef] [PubMed]
- Farfour, E.; Leto, J.; Barritault, M.; Barberis, C.; Meyer, J.; Dauphin, B.; Le Guern, A.-S.; Leflèche, A.; Badell, E.; Guiso, N.; et al. Evaluation of the andromas matrix-assisted laser desorption ionization-time of flight mass spectrometry system for identification of aerobically growing gram-positive bacilli. J. Clin. Microbiol. 2012, 50, 2702–2707. [Google Scholar] [CrossRef] [PubMed]
- Rocca, M.F.; Barrios, R.; Zintgraff, J.; Martínez, C.; Irazu, L.; Vay, C.; Prieto, M. Utility of platforms Viteks MS and Microflex LT for the identification of complex clinical isolates that require molecular methods for their taxonomic classification. PLoS ONE 2019, 14, e0218077. [Google Scholar] [CrossRef]
- Bittar, F.; Rolain, J.-M. Detection and accurate identification of new or emerging bacteria in cystic fibrosis patients. Clin. Microbiol. Infect. 2010, 16, 809–820. [Google Scholar] [CrossRef]
- Lau, S.K.P.; Chow, W.-N.; Foo, C.-H.; Curreem, S.O.T.; Lo, G.C.-S.; Teng, J.L.L.; Chen, J.H.K.; Ng, R.H.Y.; Wu, A.K.L.; Cheung, I.Y.Y.; et al. Elizabethkingia anophelis bacteremia is associated with clinically significant infections and high mortality. Sci. Rep. 2016, 6, 26045. [Google Scholar] [CrossRef]
- Cheng, Y.-H.; Perng, C.-L.; Jian, M.-J.; Lee, S.-Y.; Sun, J.-R.; Shang, H.-S. Multicentre study evaluating matrix-assisted laser desorption ionization–time of flight mass spectrometry for identification of clinically isolated Elizabethkingia species and analysis of antimicrobial susceptibility. Clin. Microbiol. Infect. 2019, 25, 340–345. [Google Scholar] [CrossRef]
- Liang, C.-Y.; Yang, C.-H.; Lai, C.-H.; Huang, Y.-H.; Lin, J.-N. Genomic Features, Comparative Genomic Analysis, and Antimicrobial Susceptibility Patterns of Chryseobacterium arthrosphaerae Strain ED882-96 Isolated in Taiwan. Genes 2019, 10, 309. [Google Scholar] [CrossRef]
- Coward, A.; Kenna, D.T.; Woodford, N.; Turton, J.F.; Armstrong, M.; Auckland, C.; Bowler, I.; Burns, P.; Cargill, J.; Carroll, M.; et al. Structured surveillance of Achromobacter, Pandoraea and Ralstonia species from patients in England with cystic fibrosis. J. Cyst. Fibros. 2020, 19, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Coenye, T.; LiPuma, J.J. Use of the gyrB gene for the identification of Pandoraea species. FEMS Microbiol. Lett. 2002, 208, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Coenye, T.; Liu, L.; Vandamme, P.; LiPuma, J.J. Identification of Pandoraea species by 16S ribosomal DNA-based PCR assays. J. Clin. Microbiol. 2001, 39, 4452–4455. [Google Scholar] [CrossRef] [PubMed]
- Coenye, T.; Vandamme, P.; LiPuma, J.J. Infection by Ralstonia species in cystic fibrosis patients: Identification of R. pickettii and R. mannitolilytica by polymerase chain reaction. Emerg. Infect. Dis. 2002, 8, 692–696. [Google Scholar] [CrossRef] [PubMed]
- Maroye, P.; Doermann, H.; Rogues, A.; Gachie, J.; Mégraud, F. Investigation of an outbreak of Ralstonia pickettii in a paediatric hospital by RAPD. J. Hosp. Infect. 2000, 44, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Sfeir, M.M. Antimicrobial susceptibility testing for glucose-nonfermenting gram-negative bacteria: The tip of the iceberg. Antimicrob. Agents Chemother. 2020, 64, e00011-20. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters Version 12.0, Valid from 2022-01-01 2022. Available online: http://www.eucast.org (accessed on 8 October 2022).
- CLSI M100-ED30:2020; Performance Standards for Antimicrobial Susceptibility Testing, 30th ed. Clinical and laboratory Standard Institute: Wayne, PA, USA, 2020.
- Kiratisin, P.; Koomanachai, P.; Kowwigkai, P.; Pattanachaiwit, S.; Aswapokee, N.; Leelaporn, A. Early-onset prosthetic valve endocarditis caused by Inquilinus sp. Diagn. Microbiol. Infect. Dis. 2006, 56, 317–320. [Google Scholar] [CrossRef]
- Wellinghausen, N.; Essig, A.; Sommerburg, O. Inquilinus limosus in patients with cystic fibrosis, Germany. Emerg. Infect. Dis. 2005, 11, 457–459. [Google Scholar] [CrossRef]
- Lenhart-Pendergrass, P.M.; Caverly, L.J.; Wagner, B.D.; Sagel, S.D.; Nick, J.A.; LiPuma, J.J.; Martiniano, S.L. Clinical characteristics and outcomes associated with Inquilinus infection in cystic fibrosis. J. Cyst. Fibros. 2021, 20, 310–315. [Google Scholar] [CrossRef]
- Cicatiello, A.G.; Iula, D.V.; Pagliuca, C.; Pastore, G.; Pagliarulo, C.; Catania, M.R.; Colicchio, R.; Picardi, M.; Raia, V.; Salvatore, P. Identification of Inquilinus limosus in cystic fibrosis: A first report in Italy. New Microbiol. 2014, 37, 567–571. [Google Scholar]
- Kohlmann, R.; Barenberg, K.; Anders, A.; Gatermann, S.G. Acetobacter indonesiensis Bacteremia in Child with Metachromatic Leukodystrophy. Emerg. Infect. Dis. 2016, 22, 1681–1683. [Google Scholar] [CrossRef] [PubMed]
- Pino, M.; Power, P.; Gutkind, G.; Di Conza, J.A. INQ-1, a chromosome-encoded AmpC β-lactamase from Inquilinus limosus. J. Antimicrob. Chemother. 2014, 69, 560–562. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Schneider, I.; Queenan, A.M.; Bauernfeind, A. Novel carbapenem-hydrolyzing oxacillinase OXA-62 from Pandoraea pnomenusa. Antimicrob. Agents Chemother. 2006, 50, 1330–1335. [Google Scholar] [CrossRef] [PubMed]
- Schneider, I.; Bauernfeind, A. Intrinsic carbapenem-hydrolyzing oxacillinases from members of the genus pandoraea. Antimicrob. Agents Chemother. 2015, 59, 7136–7141. [Google Scholar] [CrossRef] [PubMed]
- Fluit, A.C.; Bayjanov, J.R.; Aguilar, M.D.; Cantón, R.; Tunney, M.M.; Elborn, J.S.; van Westreenen, M.; Ekkelenkamp, M.B. Characterization of clinical Ralstonia strains and their taxonomic position. Antonie van Leeuwenhoek 2021, 114, 1721–1733. [Google Scholar] [CrossRef]
- Chang, Y.; Zhang, D.; Niu, S.; Chen, Q.; Lin, Q.; Zhang, X. Mbls, rather than efflux pumps, led to carbapenem resistance in fosfomycin and aztreonam/avibactam resistant elizabethkingia anophelis. Infect. Drug Resist. 2021, 14, 315–327. [Google Scholar] [CrossRef]
- González, L.J.; Vila, A.J. Carbapenem resistance in Elizabethkingia meningoseptica is mediated by metallo-β-lactamase BlaB. Antimicrob. Agents Chemother. 2012, 56, 1686–1692. [Google Scholar] [CrossRef]
- Woodford, N.; Palepou, M.-F.I.; Babini, G.S.; Holmes, B.; Livermore, D.M. Carbapenemases of Chryseobacterium (Flavobacterium) meningosepticum: Distribution of blaB and characterization of a novel metallo-β-lactamase gene, blaB3, in the type strain, NCTC 10016. Antimicrob. Agents Chemother. 2000, 44, 1448–1452. [Google Scholar] [CrossRef]
- Damas, M.S.F.; Ferreira, R.L.; Campanini, E.B.; Soares, G.G.; Campos, L.C.; Laprega, P.M.; da Costa, A.S.; Freire, C.C.D.M.; Pitondo-Silva, A.; Cerdeira, L.T.; et al. Whole genome sequencing of the multidrug-resistant Chryseobacterium indologenes isolated from a patient in Brazil. Front. Med. 2022, 9, 2143. [Google Scholar] [CrossRef]
- Wang, T.; Jiang, X.; Feng, C.; Li, A.; Dong, H.; Wu, S.; Zheng, B. Whole genome sequencing uncovers a novel IND-16 metallo-β-lactamase from an extensively drug-resistant Chryseobacterium indologenes strain J31. Gut Pathog. 2016, 8, 47. [Google Scholar] [CrossRef]
- Cimmino, T.; Rolain, J.-M. Whole genome sequencing for deciphering the resistome of Chryseobacterium indologenes, an emerging multidrug-resistant bacterium isolated from a cystic fibrosis patient in Marseille, France. New Microbes New Infect. 2016, 12, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Zhang, Q.; Gu, Z. Molecular diversity of chromosomal metallo-β-lactamase genes in Elizabethkingia genus. Int. J. Antimicrob. Agents 2020, 56, 105978. [Google Scholar] [CrossRef] [PubMed]
- Pino, M.; Di Conza, J.; Gutkind, G. Draft genome sequence of Inquilinus limosus strain MP06, a multidrug-resistant clinical isolate. Braz. J. Microbiol. 2015, 46, 943–944. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.-L.; Ee, R.; Yong, D.; Yu, C.-Y.; Ang, G.-Y.; Tee, K.-K.; Yin, W.-F.; Chan, K.-G. Complete Genome Sequence Analysis of Pandoraea pnomenusa Type Strain DSM 16536(T) Isolated from a Cystic Fibrosis Patient. Front. Microbiol. 2016, 7, 109. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, D.; Yang, Y.; Su, J.; Xu, X.; Wang, M.; Chen, Y.; Li, Y. Clinical and molecular characteristics of Chryseobacterium indologenes isolates at a teaching hospital in Shanghai, China. Ann. Transl. Med. 2021, 9, 668. [Google Scholar] [CrossRef]
- Shinha, T.; Ahuja, R. Bacteremia due to Elizabethkingia meningoseptica. IDCases 2015, 2, 13–15. [Google Scholar] [CrossRef]
- Zdziarski, P.; Paściak, M.; Rogala, K.; Korzeniowska-Kowal, A.; Gamian, A. Elizabethkingia miricola as an opportunistic oral pathogen associated with superinfectious complications in humoral immunodeficiency: A case report. BMC Infect. Dis. 2017, 17, 763. [Google Scholar] [CrossRef]
- Arbune, M.; Fotea, S.; Nechita, A.; Stefanescu, V. Emerging Infection with Elizabethkingia meningoseptica in Neonate. A Case Report. J. Crit. Care Med. 2018, 4, 96–100. [Google Scholar] [CrossRef]
- Ceyhan, M.; Yıldırım, I.; Tekelı, A.; Yurdakok, M.; Us, E.; Altun, B.; Kutluk, T.; Cengiz, A.B.; Gurbuz, V.; Barın, C.; et al. A Chryseobacterium meningosepticum outbreak observed in 3 clusters involving both neonatal and non-neonatal pediatric patients. Am. J. Infect. Control. 2008, 36, 453–457. [Google Scholar] [CrossRef]
- Olbrich, P.; Rivero-Garvía, M.; Falcón-Neyra, M.D.; Lepe, J.A.; Cisneros, J.M.; Márquez-Rivas, J.; Neth, O. Chryseobacterium indologenes central nervous system infection in infancy: An emergent pathogen? Infection 2014, 42, 179–183. [Google Scholar] [CrossRef]
- Hayes, D.; Murphy, B.S.; Kuhn, R.J.; Anstead, M.I.; Feola, D.J. Mucoid Inquilinus limosus in a young adult with cystic fibrosis. Pediatr. Pulmonol. 2009, 44, 619–621. [Google Scholar] [CrossRef] [PubMed]
- Watson, L.; Andersson, M.I.; Ives, A.; Hull, J.; Chapman, S.J.; Flight, W.G. Familial cluster of Inquilinus limosus infection among three brothers with cystic fibrosis. Respir. Med. Case Rep. 2021, 34, 101487. [Google Scholar] [CrossRef]
- Poore, T.S.; Virella-Lowell, I.; Guimbellot, J.S. Potential pathogenicity of Inquilinus limosus in a pediatric patient with cystic fibrosis. Pediatr. Pulmonol. 2018, 53, E21–E23. [Google Scholar] [CrossRef] [PubMed]
- Millard, A.D.; Westblade, L.F.; LiPuma, J.J.; Vavikolanu, K.; Read, T.D.; Pallen, M.; Burd, E.M.; Constantinidou, C.I. Draft Genome Sequence of the Pandoraea apista LMG 16407 Type Strain. Genome Announc. 2015, 3, e01300-15. [Google Scholar] [CrossRef]
- Mwanza, E.P.; Hugo, A.; Charimba, G.; Hugo, C.J. Pathogenic Potential and Control of Chryseobacterium Species from Clinical, Fish, Food and Environmental Sources. Microorganisms 2022, 10, 895. [Google Scholar] [CrossRef]
- Zajmi, A.; Teo, J.; Yeo, C.C. Epidemiology and Characteristics of Elizabethkingia spp. Infections in Southeast Asia. Microorganisms 2022, 10, 882. [Google Scholar] [CrossRef]
- Herasimenka, Y.; Cescutti, P.; Impallomeni, G.; Rizzo, R. Exopolysaccharides produced by Inquilinus limosus, a new pathogen of cystic fibrosis patients: Novel structures with usual components. Carbohydr. Res. 2007, 342, 2404–2415. [Google Scholar] [CrossRef]
- Maunders, E.; Welch, M. Matrix exopolysaccharides; the sticky side of biofilm formation. FEMS Microbiol. Lett. 2017, 364, 120. [Google Scholar] [CrossRef]
- Kuttel, M.; Ravenscroft, N.; Foschiatti, M.; Cescutti, P.; Rizzo, R. Conformational properties of two exopolysaccharides produced by Inquilinus limosus, a cystic fibrosis lung pathogen. Carbohydr. Res. 2012, 350, 40–48. [Google Scholar] [CrossRef]
- Costello, A.; Herbert, G.; Fabunmi, L.; Schaffer, K.; Kavanagh, K.; Caraher, E.M.; Callaghan, M.; McClean, S. Virulence of an emerging respiratory pathogen, genus Pandoraea, in vivo and its interactions with lung epithelial cells. J. Med. Microbiol. 2011, 60, 289–299. [Google Scholar] [CrossRef]
- Lin, P.-Y.; Chiu, C.-H.; Chu, C.; Tang, P.; Su, L.-H. Invasion of murine respiratory tract epithelial cells by Chryseobacterium meningosepticum and identification of genes present specifically in an invasive strain. New Microbiol. 2006, 29, 55–62. [Google Scholar] [PubMed]
- Lin, P.-Y.; Chen, H.-L.; Huang, C.-T.; Su, L.-H.; Chiu, C.-H. Biofilm production, use of intravascular indwelling catheters and inappropriate antimicrobial therapy as predictors of fatality in Chryseobacterium meningosepticum bacteraemia. Int. J. Antimicrob. Agents 2010, 36, 436–440. [Google Scholar] [CrossRef] [PubMed]
- Puah, S.M.; Fong, S.P.; Kee, B.P.; Puthucheary, S.; Chua, K.H. Molecular identification and biofilm-forming ability of Elizabethkingia species. Microb. Pathog. 2022, 162, 105345. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-C.; Lo, H.-H.; Hsieh, H.-Y.; Chang, S.-M. Identification, epidemiological relatedness, and biofilm formation of clinical Chryseobacterium indologenes isolates from central Taiwan. J. Microbiol. Immunol. Infect. 2015, 48, 559–564. [Google Scholar] [CrossRef]
- Hibbing, M.E.; Fuqua, C.; Parsek, M.R.; Peterson, S.B. Bacterial competition: Surviving and thriving in the microbial jungle. Nat. Rev. Microbiol. 2009, 8, 15–25. [Google Scholar] [CrossRef]
- Rogers, G.B.; Hoffman, L.R.; Whiteley, M.; Daniels, T.W.; Carroll, M.P.; Bruce, K.D. Revealing the dynamics of polymicrobial infections: Implications for antibiotic therapy. Trends Microbiol. 2010, 18, 357–364. [Google Scholar] [CrossRef]
- Magalhães, A.P.; Lopes, S.P.; Pereira, M.O. Insights into Cystic Fibrosis Polymicrobial Consortia: The Role of Species Interactions in Biofilm Development, Phenotype, and Response to In-Use Antibiotics. Front. Microbiol. 2017, 7, 2146. [Google Scholar] [CrossRef]
- O’brien, T.J.; Figueroa, W.; Welch, M. Decreased efficacy of antimicrobial agents in a polymicrobial environment. ISME J. 2022, 16, 1694–1704. [Google Scholar] [CrossRef]
- Coman, I.; Bilodeau, L.; Lavoie, A.; Carricart, M.; Tremblay, F.; Zlosnik, J.; Berthiaume, Y. Ralstonia mannitolilytica in cystic fibrosis: A new predictor of worse outcomes. Respir. Med. Case Rep. 2017, 20, 48–50. [Google Scholar] [CrossRef]
- Cooke, R.; O’neill, W.; Xu, J.; Moore, J.; Elborn, J. Inquilinus limosus isolated from a cystic fibrosis patient: First UK report. Br. J. Biomed. Sci. 2007, 64, 127–129. [Google Scholar] [CrossRef]
- Degand, N.; Lotte, R.; Le Butor, C.D.; Segonds, C.; Thouverez, M.; Ferroni, A.; Vallier, C.; Mély, L.; Carrère, J. Epidemic spread of Pandoraea pulmonicola in a cystic fibrosis center. BMC Infect. Dis. 2015, 15, 583. [Google Scholar] [CrossRef] [PubMed]
- The UK Cystic Fibrosis Trust Infection Control Group. Standards for the Clinical Care of Children and Adults with Cystic Fibrosis in the UK; The UK Cystic Fibrosis Trust Infection Control Group: Bromley, UK, 2011. [Google Scholar]
- The UK Cystic Fibrosis Trust Infection Control Group. The Burkholderia cepacia Complex—Suggestions for Prevention and Infection Control; The UK Cystic Fibrosis Trust Infection Control Group: Bromley, UK, 2004. [Google Scholar]
- Saiman, L.; Siegel, J.D.; Lipuma, J.J.; Brown, R.; Bryson, E.A.; Chambers, M.J.; Downer, V.S.; Fliege, J.; Hazle, L.A.; Jain, M.; et al. Infection Prevention and Control Guideline for Cystic Fibrosis: 2013 Update. Infect. Control. Hosp. Epidemiology 2014, 35, s1–s67. [Google Scholar] [CrossRef] [PubMed]
- Smyth, A.R.; Smith, S.J.; Rowbotham, N.J. Infection prevention and control in cystic fibrosis: One size fits all The argument against. Paediatr. Respir. Rev. 2019, 36, 94–96. [Google Scholar] [CrossRef]
- Stockwell, R.E.; Wood, M.E.; Ballard, E.; Moore, V.; Wainwright, C.E.; Bell, S.C. Current infection control practices used in Australian and New Zealand cystic fibrosis centers. BMC Pulm. Med. 2020, 20, 16–18. [Google Scholar] [CrossRef]
- Saiman, L.; Zhou, J.J.; Jiang, X.; Kosorok, M.R.; Muhlebach, M.S. Surveying Cystic Fibrosis Care Centers to Assess Adoption of Infection Prevention and Control Recommendations. Infect. Control. Hosp. Epidemiol. 2018, 39, 647–651. [Google Scholar] [CrossRef] [PubMed]
- Walter, J.; Foley, P.; Yason, C.; Vanderstichel, R.; Muckle, A. Prevalence of feline herpesvirus-1, feline calicivirus, chlamydia felis, and bordetella bronchiseptica in a population of shelter cats on Prince Edward island. Can. J. Vet. Res. 2020, 84, 181–188. [Google Scholar]
- Berger, A.; Willi, B.; Meli, M.L.; Boretti, F.S.; Hartnack, S.; Dreyfus, A.; Lutz, H.; Hofmann-Lehmann, R. Feline calicivirus and other respiratory pathogens in cats with Feline calicivirusrelated symptoms and in clinically healthy cats in Switzerland. BMC Veter. Res. 2015, 11, 282. [Google Scholar] [CrossRef]
- Binns, S.H.; Dawson, S.; Speakman, A.J.; Cuevas, L.E.; Gaskell, C.J.; Hart, C.A.; Morgan, K.L.; Gaskell, R.M. Prevalence and risk factors for feline Bordetella bronchiseptica infection. Veter. Rec. 1999, 144, 575–580. [Google Scholar] [CrossRef]
- Day, M.; Carey, S.; Clercx, C.; Kohn, B.; MarsilIo, F.; Thiry, E.; Freyburger, L.; Schulz, B.; Walker, D. Aetiology of Canine Infectious Respiratory Disease Complex and Prevalence of its Pathogens in Europe. J. Comp. Pathol. 2020, 176, 86–108. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Vaccine Scheduler—Pertussis: Recommended Vaccinations n.d. Available online: https://vaccine-schedule.ecdc.europa.eu/Scheduler/ByDisease?SelectedDiseaseId=3&SelectedCountryIdByDisease=-1 (accessed on 8 October 2022).
- Saul, N.; Wang, K.; Bag, S.; Baldwin, H.; Alexander, K.; Chandra, M.; Thomas, J.; Quinn, H.; Sheppeard, V.; Conaty, S. Effectiveness of maternal pertussis vaccination in preventing infection and disease in infants: The NSW Public Health Network case-control study. Vaccine 2018, 36, 1887–1892. [Google Scholar] [CrossRef]
- Rowe, S.L.; Tay, E.L.; Franklin, L.J.; Stephens, N.; Ware, R.; Kaczmarek, M.C.; Lester, R.A.; Lambert, S.B. Effectiveness of parental cocooning as a vaccination strategy to prevent pertussis infection in infants: A case-control study. Vaccine 2018, 36, 2012–2019. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.E.; Rendall, J.C.; Millar, B.C. A doggy tale: Risk of zoonotic infection with Bordetella bronchiseptica for cystic fibrosis (CF) patients from live licenced bacterial veterinary vaccines for cats and dogs. J. Clin. Pharm. Ther. 2022, 47, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.E.; Rendall, J.C.; Millar, B.C. Does Bordetella pertussis vaccine offer any cross-protection against Bordetella bronchiseptica? Implications for pet owners with cystic fibrosis. J. Clin. Pharm. Ther. 2021, 46, 1194–1198. [Google Scholar] [CrossRef] [PubMed]

| Family | Genera (Number of Report) |
|---|---|
| Acebobacteraceae | Acetobacter (2) |
| Alcaligenaceae | Achromobacter *, Bordetella (4) |
| Burkholderiacaea | Burkholderia *, Pandoraea (6) |
| Flavobacteriaceae | None |
| Pseudomonaceae | Pseudomonas * |
| Rhodospirillaceae | Inquilinus (3) |
| Xanthomonadaceae | Stenotrophomonas * |
| Weeksellaceae | Chryseobacterium (1), Elizabethkinga (1) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farfour, E.; Roux, A.; Sage, E.; Revillet, H.; Vasse, M.; Vallée, A. Rarely Encountered Gram-Negative Rods and Lung Transplant Recipients: A Narrative Review. Microorganisms 2023, 11, 1468. https://doi.org/10.3390/microorganisms11061468
Farfour E, Roux A, Sage E, Revillet H, Vasse M, Vallée A. Rarely Encountered Gram-Negative Rods and Lung Transplant Recipients: A Narrative Review. Microorganisms. 2023; 11(6):1468. https://doi.org/10.3390/microorganisms11061468
Chicago/Turabian StyleFarfour, Eric, Antoine Roux, Edouard Sage, Hélène Revillet, Marc Vasse, and Alexandre Vallée. 2023. "Rarely Encountered Gram-Negative Rods and Lung Transplant Recipients: A Narrative Review" Microorganisms 11, no. 6: 1468. https://doi.org/10.3390/microorganisms11061468
APA StyleFarfour, E., Roux, A., Sage, E., Revillet, H., Vasse, M., & Vallée, A. (2023). Rarely Encountered Gram-Negative Rods and Lung Transplant Recipients: A Narrative Review. Microorganisms, 11(6), 1468. https://doi.org/10.3390/microorganisms11061468