
Pathogenic and Opportunistic Vibrio spp. Associated with Vibriosis Incidences in the Greek Aquaculture: The Role of Vibrio harveyi as the Principal Cause of Vibriosis


Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Histopathology
2.3. Molecular Identification
2.4. Metabolic Fingerprint
3. Results
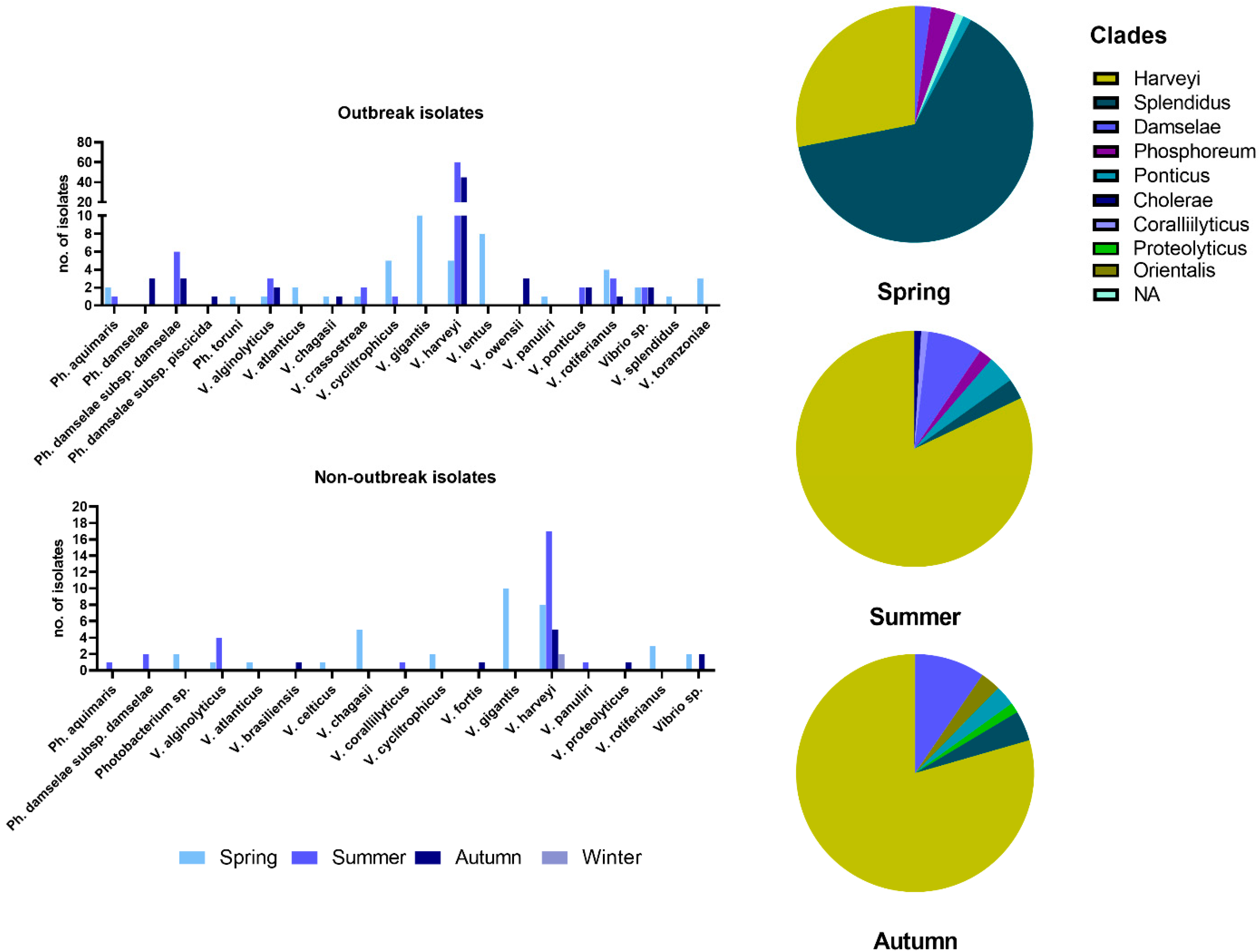
3.1. Bacterial Identification
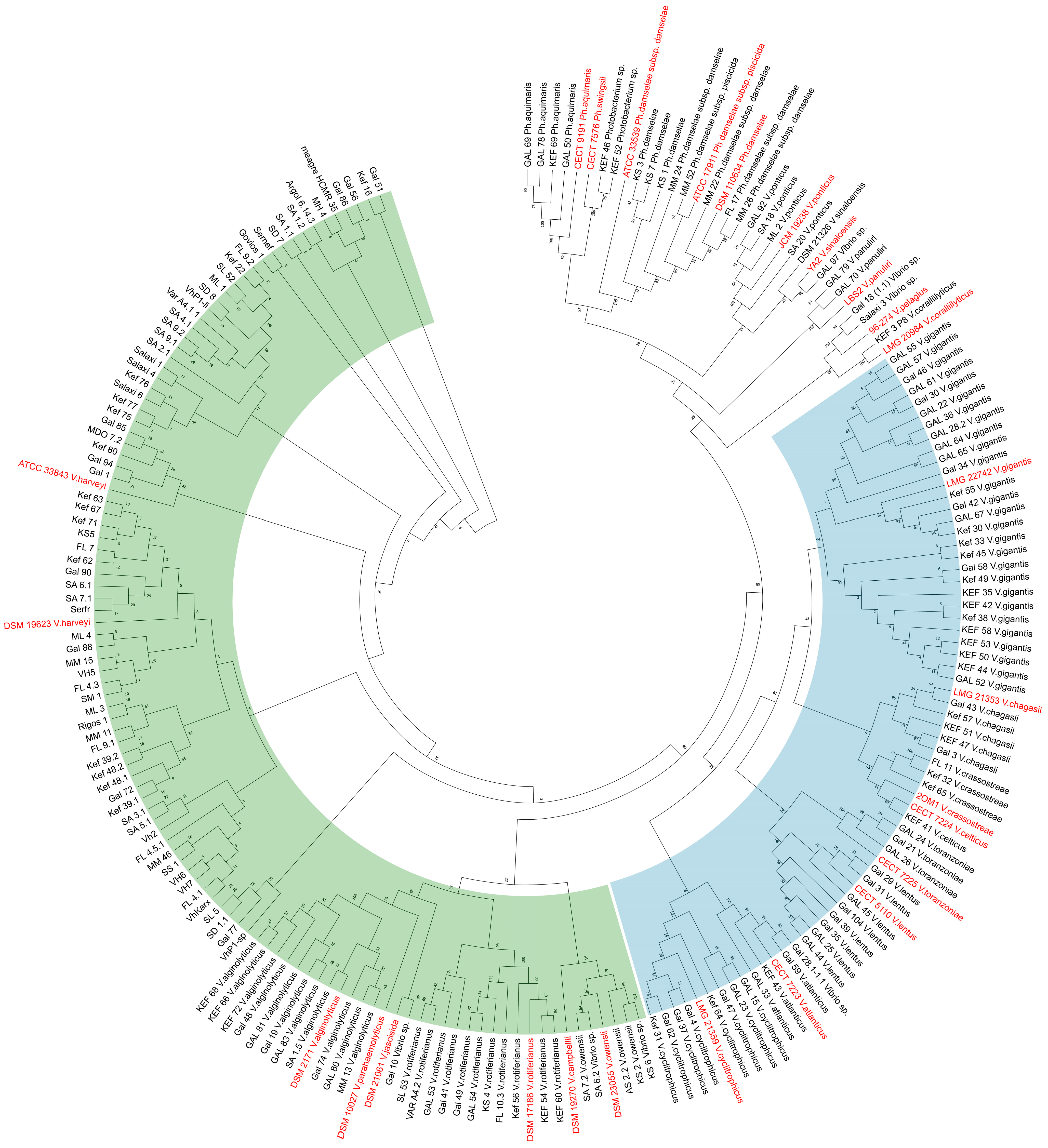
3.2. Seasonality of Vibrio Species and Phylogenetic Relationships
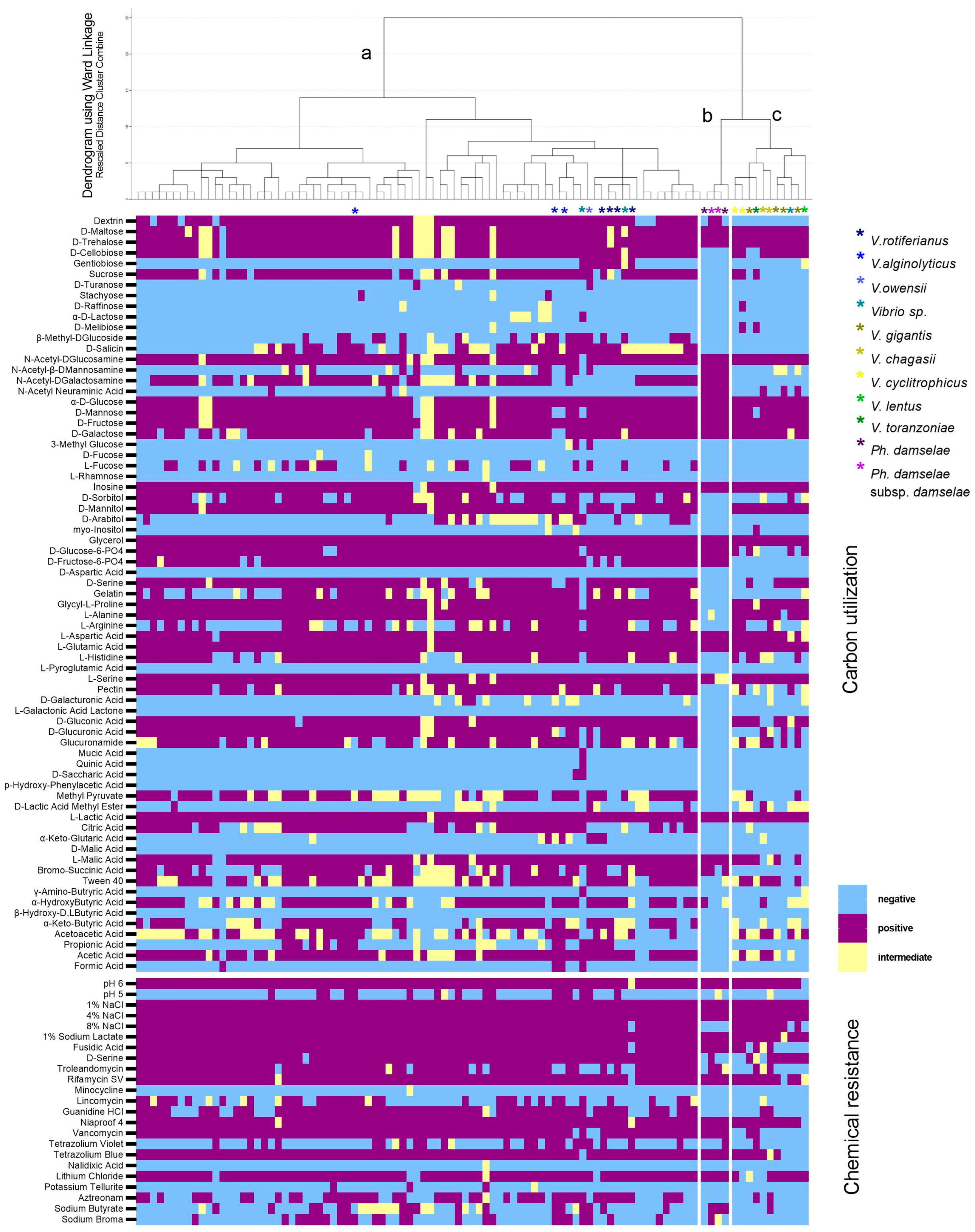
3.3. Metabolic Fingerprint
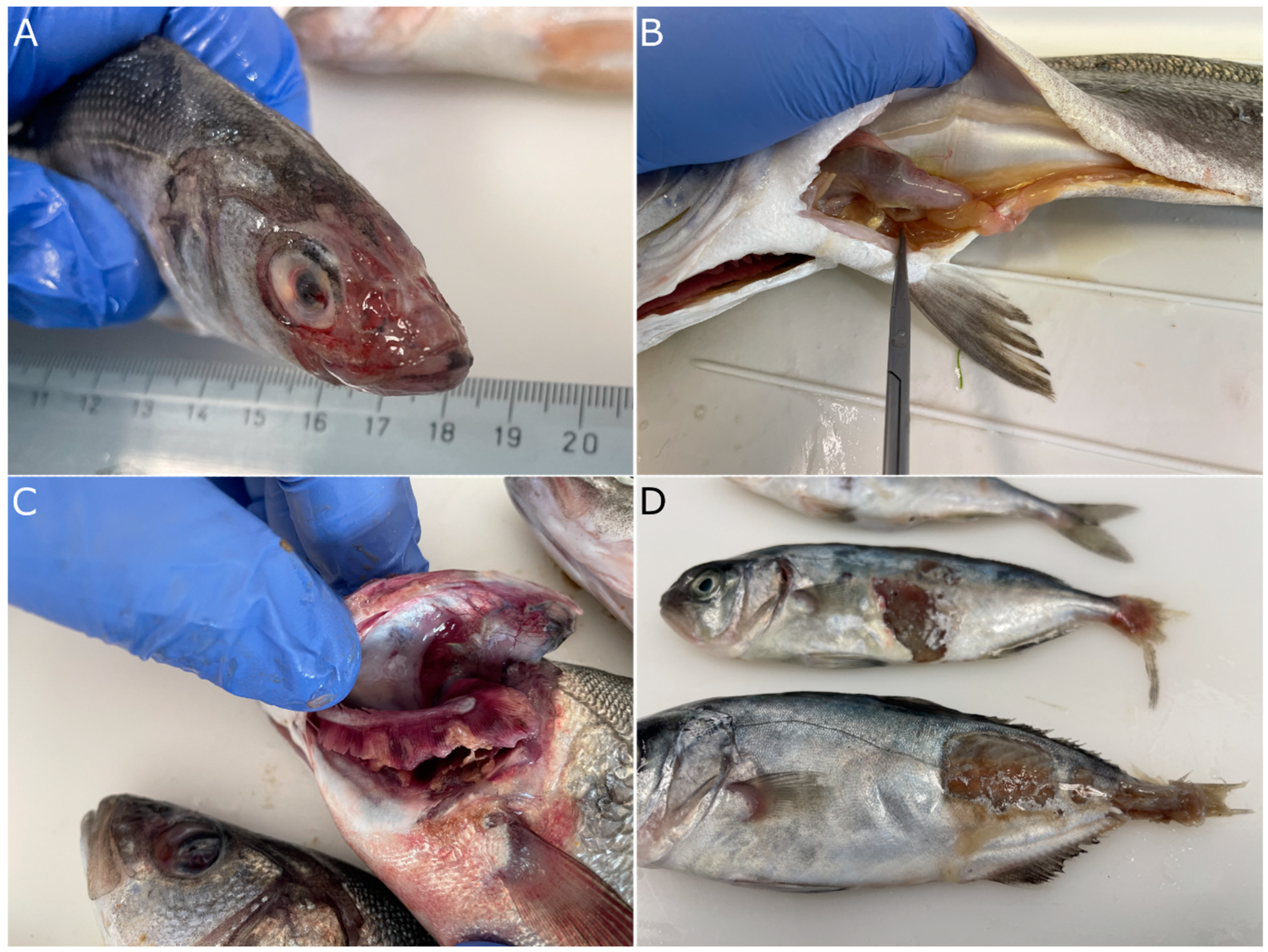
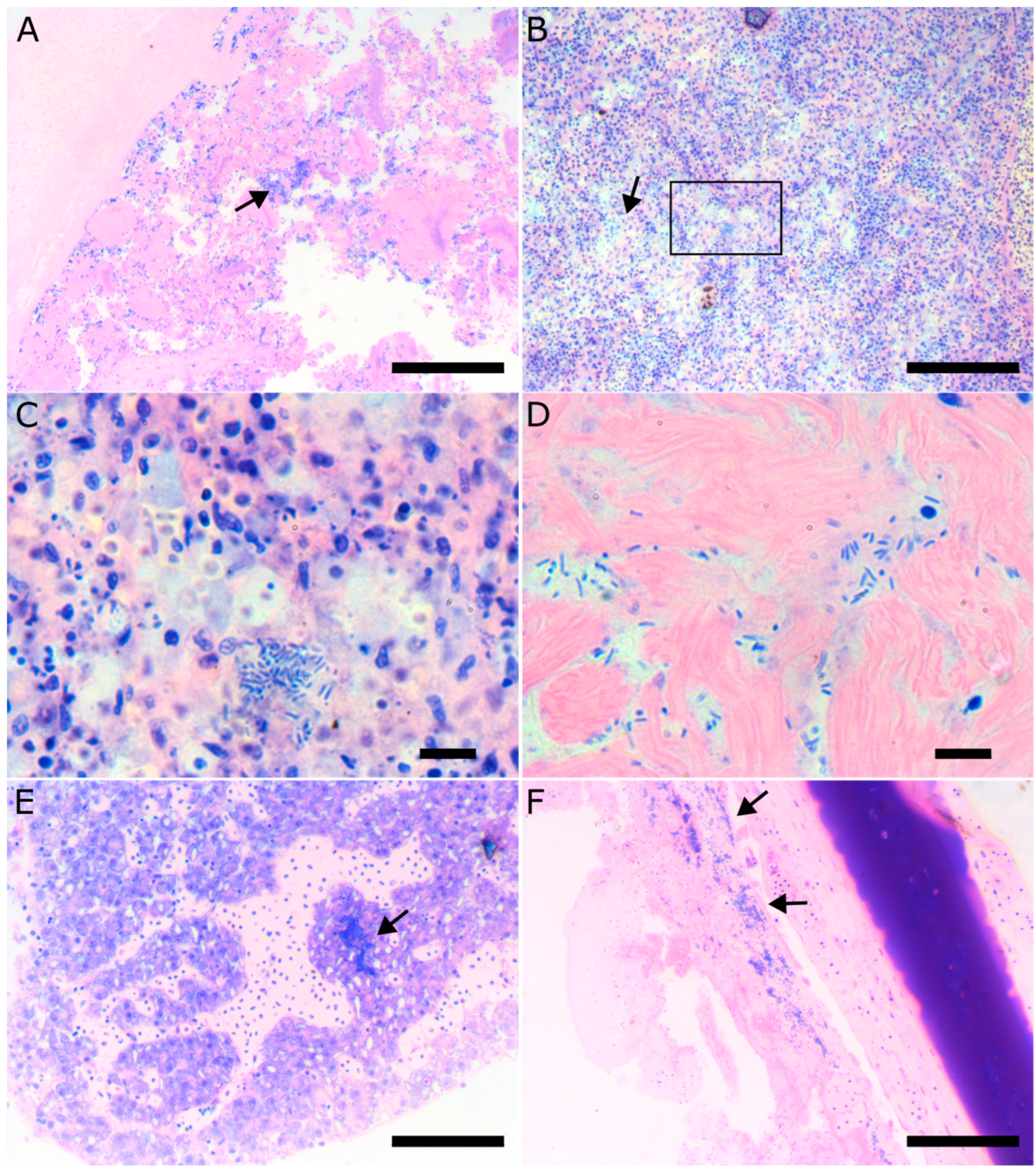
3.4. Pathology of Vibriosis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roberts, R.J. Fish Pathology; Wiley: New York, NY, USA, 2012. [Google Scholar]
- Mohd Yazid, S.H.; Mohd Daud, H.; Azmai, M.N.A.; Mohamad, N.; Mohd Nor, N. Estimating the Economic Loss Due to Vibriosis in Net-Cage Cultured Asian Seabass (Lates calcarifer): Evidence From the East Coast of Peninsular Malaysia. Front. Vet. Sci. 2021, 8, 644009. [Google Scholar] [CrossRef] [PubMed]
- Rivas, A.J.; Lemos, M.L.; Osorio, C.R. Photobacterium Damselae subsp. damselae, a Bacterium Pathogenic for Marine Animals and Humans. Front. Microbiol. 2013, 4, 283. [Google Scholar] [CrossRef]
- Pérez-Cataluña, A.; Lucena, T.; Tarazona, E.; Arahal, D.R.; Macián, M.C.; Pujalte, M.J. An MLSA Approach for the Taxonomic Update of the Splendidus Clade, a Lineage Containing Several Fish and Shellfish Pathogenic Vibrio spp. Syst. Appl. Microbiol. 2016, 39, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Ina-Salwany, M.Y.; Al-saari, N.; Mohamad, A.; Mursidi, F.A.; Mohd-Aris, A.; Amal, M.N.A.; Kasai, H.; Mino, S.; Sawabe, T.; Zamri-Saad, M. Vibriosis in Fish: A Review on Disease Development and Prevention. J. Aquat. Anim. Health 2019, 31, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Gatesoupe, F.-J.; Lambert, C.; Nicolas, J.-L. Pathogenicity of Vibrio Splendidus Strains Associated with Turbot Larvae, Scophthalmus Maximus. J. Appl. Microbiol. 1999, 87, 757–763. [Google Scholar] [CrossRef]
- Pascual, J.; Macián, M.C.; Arahal, D.R.; Garay, E.; Pujalte, M.J. Multilocus Sequence Analysis of the Central Clade of the Genus Vibrio by Using the 16S RRNA, RecA, PyrH, RpoD, GyrB, RctB and ToxR Genes. Int. J. Syst. Evol. Microbiol. 2010, 60, 154–165. [Google Scholar] [CrossRef]
- Hoffmann, M.; Monday, S.R.; Fischer, M.; Brown, E.W. Genetic and Phylogenetic Evidence for Misidentification of Vibrio Species within the Harveyi Clade. Lett. Appl. Microbiol. 2012, 54, 160–165. [Google Scholar] [CrossRef]
- Sawabe, T.; Ogura, Y.; Matsumura, Y.; Feng, G.; Rohul Amin, A.K.M.; Mino, S.; Nakagawa, S.; Sawabe, T.; Kumar, R.; Fukui, Y.; et al. Updating the Vibrio Clades Defined by Multilocus Sequence Phylogeny: Proposal of Eight New Clades, and the Description of Vibrio tritonius sp. Nov. Front. Microbiol. 2013, 4, 414. [Google Scholar] [CrossRef] [PubMed]
- Culot, A.; Grosset, N.; Bruey, Q.; Auzou, M.; Giard, J.C.; Favard, B.; Wakatsuki, A.; Baron, S.; Frouel, S.; Techer, C.; et al. Isolation of Harveyi Clade Vibrio spp. Collected in Aquaculture Farms: How Can the Identification Issue Be Addressed? J. Microbiol. Methods 2021, 180, 106106. [Google Scholar] [CrossRef]
- Urbanczyk, H.; Ogura, Y.; Hayashi, T. Taxonomic Revision of Harveyi Clade Bacteria (Family Vibrionaceae) Based on Analysis of Whole Genome Sequences. Int. J. Syst. Evol. Microbiol. 2013, 63, 2742–2751. [Google Scholar] [CrossRef] [PubMed]
- Cano-Gomez, A.; Bourne, D.G.; Hall, M.R.; Owens, L.; Høj, L. Molecular Identification, Typing and Tracking of Vibrio harveyi in Aquaculture Systems: Current Methods and Future Prospects. Aquaculture 2009, 287, 1–10. [Google Scholar] [CrossRef]
- Bellos, G.; Angelidis, P.; Miliou, H. Effect of Temperature and Seasonality Principal Epizootiological Risk Factor on Vibriosis and Photobacteriosis Outbreaks for European Sea Bass in Greece (1998–2013). J. Aquac. Res. Dev. 2015, 6, 1000338. [Google Scholar] [CrossRef]
- Cascarano, M.C.; Stavrakidis-Zachou, O.; Mladineo, I.; Thompson, K.D.; Papandroulakis, N.; Katharios, P. Mediterranean Aquaculture in a Changing Climate: Temperature Effects on Pathogens and Diseases of Three Farmed Fish Species. Pathogens 2021, 10, 1205. [Google Scholar] [CrossRef] [PubMed]
- Rigos, G.; Katharios, P. Pathological Obstacles of Newly-Introduced Fish Species in Mediterranean Mariculture: A Review. Rev. Fish Biol. Fish. 2010, 20, 47–70. [Google Scholar] [CrossRef]
- Tagliavia, M.; Salamone, M.; Bennici, C.; Quatrini, P.; Cuttitta, A. A Modified Culture Medium for Improved Isolation of Marine Vibrios. Microbiologyopen 2019, 8, e-835. [Google Scholar] [CrossRef] [PubMed]
- Conejero, M.J.U.; Hedreyda, C.T. Isolation of Partial ToxR Gene of Vibrio harveyi and Design of ToxR-Targeted PCR Primers for Species Detection. J. Appl. Microbiol. 2003, 95, 602–611. [Google Scholar] [CrossRef] [PubMed]
- Sawabe, T.; Kita-Tsukamoto, K.; Thompson, F.L. Inferring the Evolutionary History of Vibrios by Means of Multilocus Sequence Analysis. J. Bacteriol. 2007, 189, 7932–7936. [Google Scholar] [CrossRef] [PubMed]
- Cano-Gomez, A.; Høj, L.; Owens, L.; Andreakis, N. Multilocus Sequence Analysis Provides Basis for Fast and Reliable Identification of Vibrio harveyi-Related Species and Reveals Previous Misidentification of Important Marine Pathogens. Syst. Appl. Microbiol. 2011, 34, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Mougin, J.; Roquigny, R.; Travers, M.A.; Grard, T.; Bonnin-Jusserand, M.; Le Bris, C. Development of a MreB-Targeted Real-Time PCR Method for the Quantitative Detection of Vibrio harveyi in Seawater and Biofilm from Aquaculture Systems. Aquaculture 2020, 525, 735337. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and Applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Baratloo, A.; Hosseini, M.; Negida, A.; El Ashal, G. Part 1: Simple Definition and Calculation of Accuracy, Sensitivity and Specificity. Emergency 2015, 3, 48–49. [Google Scholar] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Montánchez, I.; Ogayar, E.; Plágaro, A.H.; Esteve-Codina, A.; Gómez-Garrido, J.; Orruño, M.; Arana, I.; Kaberdin, V.R. Analysis of Vibrio harveyi Adaptation in Sea Water Microcosms at Elevated Temperature Provides Insights into the Putative Mechanisms of Its Persistence and Spread in the Time of Global Warming. Sci. Rep. 2019, 9, 289. [Google Scholar] [CrossRef]
- Xu, X.; Li, H.; Qi, X.; Chen, Y.; Qin, Y.; Zheng, J.; Jiang, X. CheA, CheB, CheR, CheV, and CheY Are Involved in Regulating the Adhesion of Vibrio harveyi. Front. Cell. Infect. Microbiol. 2021, 10, 591751. [Google Scholar] [CrossRef]
- Stavrakidis-Zachou, O.; Lika, K.; Pavlidis, M.; Asaad, M.H.; Papandroulakis, N. Metabolic Scope, Performance and Tolerance of Juvenile European Sea Bass Dicentrarchus Labrax upon Acclimation to High Temperatures. PLoS ONE 2022, 17, e0272510. [Google Scholar] [CrossRef]
- Xu, W.; Gong, L.; Yang, S.; Gao, Y.; Ma, X.; Xu, L.; Chen, H.; Luo, Z. Spatiotemporal Dynamics of Vibrio Communities and Abundance in Dongshan Bay, South of China. Front. Microbiol. 2020, 11, 575287. [Google Scholar] [CrossRef]
- Sitjà-Bobadilla, A.; Pujalte, M.J.; Bermejo, A.; Garay, E.; Alvarez-Pellitero, P.; Pérez-Sánchez, J. Bacteria Associated with Winter Mortalities in Laboratory-Reared Common Dentex (Dentex dentex L.). Aquac. Res. 2007, 38, 733–739. [Google Scholar] [CrossRef]
- Austin, B.; Austin, D.; Sutherland, R.; Thompson, F.; Swings, J. Pathogenicity of Vibrios to Rainbow Trout (Oncorhynchus mykiss, Walbaum) and Artemia Nauplii. Environ. Microbiol. 2005, 7, 1488–1495. [Google Scholar] [CrossRef]
- Sohn, H.; Kim, J.; Jin, C.; Lee, J. Identification of Vibrio Species Isolated from Cultured Olive Flounder (Paralichthys olivaceus) in Jeju Island, South Korea. Fish. Aquat. Sci. 2019, 22, 14. [Google Scholar] [CrossRef]
- Mohamad, N.; Amal, M.N.A.; Yasin, I.S.M.; Zamri Saad, M.; Nasruddin, N.S.; Al-saari, N.; Mino, S.; Sawabe, T. Vibriosis in Cultured Marine Fishes: A Review. Aquaculture 2019, 512, 734289. [Google Scholar] [CrossRef]
- Korun, J. Marine Vibrios Associated with Diseased Sea Bass (Dicentrarchus labrax) in Turkey. J. Fish. Sci. 2008, 2, 66–76. [Google Scholar] [CrossRef]
- Mougin, J.; Roquigny, R.; Flahaut, C.; Bonnin-Jusserand, M.; Grard, T.; le Bris, C. Abundance and Spatial Patterns over Time of Vibrionaceae and Vibrio harveyi in Water and Biofilm from a Seabass Aquaculture Facility. Aquaculture 2021, 542, 736862. [Google Scholar] [CrossRef]
- Haldar, S.; Maharajan, A.; Chatterjee, S.; Hunter, S.A.; Chowdhury, N.; Hinenoya, A.; Asakura, M.; Yamasaki, S. Identification of Vibrio harveyi as a Causative Bacterium for a Tail Rot Disease of Sea Bream Sparus Aurata from Research Hatchery in Malta. Microbiol. Res. 2010, 165, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Turgay, E.; Steinum, T.M.; Colquhoun, D.; Karataş, S. Environmental Biofilm Communities Associated with Early-Stage Common Dentex (Dentex dente) Culture. J. Appl. Microbiol. 2019, 126, 1032–1043. [Google Scholar] [CrossRef]
- Snoussi, M.; Chaieb, K.; Mahmoud, R.; Bakhrouf, A. Quantitative Study, Identification and Antibiotics Sensitivity of Some Vibrionaceae Associated to a Marine Fish Hatchery. Ann. Microbiol. 2006, 56, 289–293. [Google Scholar] [CrossRef]
- Dong, H.T.; Taengphu, S.; Sangsuriya, P.; Charoensapsri, W.; Phiwsaiya, K.; Sornwatana, T.; Khunrae, P.; Rattanarojpong, T.; Senapin, S. Recovery of Vibrio harveyi from Scale Drop and Muscle Necrosis Disease in Farmed Barramundi, Lates calcarifer in Vietnam. Aquaculture 2017, 473, 89–96. [Google Scholar] [CrossRef]
- Avendaño-Herrera, R.; Toranzo, A.E.; Magariños, B. Tenacibaculosis Infection in Marine Fish Caused by Tenacibaculum maritimum: A Review. Dis. Aquat. Org. 2006, 71, 255–266. [Google Scholar] [CrossRef]
- Pujalte, M.J.; Sitjà-Bobadilla, A.; Macián, M.C.; Belloch, C.; Álvarez-Pellitero, P.; Pérez-Sánchez, J.; Uruburu, F.; Garay, E. Virulence and Molecular Typing of Vibrio harveyi Strains Isolated from Cultured Dentex, Gilthead Sea Bream and European Sea Bass. Syst. Appl. Microbiol. 2003, 26, 284–292. [Google Scholar] [CrossRef]
- Company, R.; Sitj, A.; Pujalte, M.J.; Garay, E.; Alvarez-Pellitero, P. Bacterial and Parasitic Pathogens in Cultured Common Dentex, Dentex dentex L. J. Fish Dis. 2002, 22, 299–309. [Google Scholar] [CrossRef]
- Castillo, D.; D’Alvise, P.; Middelboe, M.; Gram, L.; Liu, S.; Kalatzis, P.G.; Kokkari, C.; Katharios, P. Draft Genome Sequences of the Fish Pathogen Vibrio harveyi Strains VH2 and VH5. Genome Announc. 2015, 3, 2–4. [Google Scholar] [CrossRef] [PubMed]
- Al-Harbi, A.H.; Uddin, N. Bacterial Diversity of Tilapia (Oreochromis niloticus) Cultured in Brackish Water in Saudi Arabia. Aquaculture 2005, 250, 566–572. [Google Scholar] [CrossRef]
- Alcaide, E. Numerical Taxonomy of Vibrionaceae Isolated from Cultured Amberjack (Seriola dumerili) and Surrounding Water. Curr. Microbiol. 2003, 46, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Haastein, T.; Holt, G. The Occurrence of Vibrio Disease in Wild Norwegian Fish. J. Fish Biol. 1972, 4, 33–37. [Google Scholar] [CrossRef]
- Regev, Y.; Davidovich, N.; Berzak, R.; Lau, S.C.K.; Scheinin, A.P.; Tchernov, D.; Morick, D. Molecular Identification and Characterization of Vibrio Species and Mycobacterium Species in Wild and Cultured Marine Fish from the Eastern Mediterranean Sea. Microorganisms 2020, 8, 863. [Google Scholar] [CrossRef]
- Arab, S.; Nalbone, L.; Giarratana, F.; Berbar, A. Occurrence of Vibrio spp. along the Algerian Mediterranean Coast in Wild and Farmed Sparus aurata and Dicentrarchus labrax. Vet. World 2020, 13, 1199–1208. [Google Scholar] [CrossRef]
- Zhang, X.-H.; He, X.; Austin, B. Vibrio harveyi: A Serious Pathogen of Fish and Invertebrates in Mariculture. Mar. Life Sci. Technol. 2020, 2, 231–245. [Google Scholar] [CrossRef]
- Mohamad, N.; Mohd Roseli, F.A.; Azmai, M.N.A.; Saad, M.Z.; Md Yasin, I.S.; Zulkiply, N.A.; Nasruddin, N.S. Natural Concurrent Infection of Vibrio harveyi and V. alginolyticus in Cultured Hybrid Groupers in Malaysia. J. Aquat. Anim. Health 2019, 31, 88–96. [Google Scholar] [CrossRef]
- Zorrilla, I.; Chabrillón, M.; Arijo, S.; Díaz-Rosales, P.; Martínez-Manzanares, E.; Balebona, M.C.; Moriñigo, M.A. Bacteria Recovered from Diseased Cultured Gilthead Sea Bream (Sparus aurata L.) in Southwestern Spain. Aquaculture 2003, 218, 11–20. [Google Scholar] [CrossRef]
- Damir, K.; Irena, V.S.; Damir, V.; Emin, T. Occurrence, Characterization and Antimicrobial Susceptibility of Vibrio alginolyticus in the Eastern Adriatic Sea. Mar. Pollut. Bull. 2013, 75, 46–52. [Google Scholar] [CrossRef]
- Eissa, I.A.M.; Derwa, H.I.; Ismail, M.; El-lamie, M.; Dessouki, A.A.; Elsheshtawy, H.; Bayoumy, E.M. Molecular and Phenotypic Characterization of Photobacterium Damselae among Some Marine Fishes in Lake Temsah. Microb. Pathog. 2018, 114, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Labella, A.M.; Rosado, J.J.; Balado, M.; Lemos, M.L.; Borrego, J.J. Virulence Properties of Three New Photobacterium Species Affecting Cultured Fish. J. Appl. Microbiol. 2020, 129, 37–50. [Google Scholar] [CrossRef]
- Labella, A.; Manchado, M.; Alonso, M.C.; Castro, D.; Romalde, J.L.; Borrego, J.J. Molecular Intraspecific Characterization of Photobacterium damselae ssp. damselae Strains Affecting Cultured Marine Fish. J. Appl. Microbiol. 2009. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Trinanes, J.; Gonzalez-Escalona, N.; Martinez-Urtaza, J. Non-Cholera Vibrios: The Microbial Barometer of Climate Change. Trends Microbiol. 2017, 25, 76–84. [Google Scholar] [CrossRef]
- Montánchez, I.; Kaberdin, V.R. Vibrio harveyi: A Brief Survey of General Characteristics and Recent Epidemiological Traits Associated with Climate Change. Mar. Environ. Res. 2020, 154, 104850. [Google Scholar] [CrossRef]
- Izumiya, H.; Furukawa, M.; Ogata, K.; Isobe, J.; Watanabe, S.; Sasaki, M.; Ichinose, K.; Arakawa, E.; Morita, M.; Kurane, I.; et al. A Double-Quadratic Model for Predicting Vibrio Species in Water Environments of Japan. Arch. Microbiol. 2017, 199, 1293–1302. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carbon Utilization | Negative | Positive | Intermediate | Carbon Utilization | Negative | Positive | Intermediate |
|---|---|---|---|---|---|---|---|
| 3-Methyl Glucose | 97% | 1% | 1% | L-Serine | 1% | 94% | 4% |
| Acetic Acid | 7% | 81% | 13% | Methyl Pyruvate | 11% | 63% | 26% |
| Acetoacetic Acid | 26% | 33% | 40% | Mucic Acid | 100% | 0% | 0% |
| Bromo-Succinic Acid | 24% | 64% | 13% | myo-Inositol | 97% | 0% | 3% |
| Citric Acid | 24% | 67% | 10% | N-Acetyl-Neuraminic Acid | 96% | 3% | 1% |
| D-Arabitol | 68% | 14% | 18% | N-Acetyl-D-Galactosamine | 11% | 74% | 15% |
| D-Aspartic Acid | 100% | 0% | 0% | N-Acetyl-D-Glucosamine | 1% | 92% | 7% |
| D-Cellobiose | 3% | 85% | 13% | N-Acetyl-β-D-Mannosamine | 82% | 13% | 6% |
| Dextrin | 10% | 86% | 4% | Pectin | 7% | 85% | 8% |
| D-Fructose | 0% | 92% | 8% | p-Hydroxy-Phenylacetic Acid | 100% | 0% | 0% |
| D-Fructose-6-PO4 | 3% | 96% | 1% | Propionic Acid | 81% | 11% | 8% |
| D-Fucose | 97% | 0% | 3% | Quinic Acid | 100% | 0% | 0% |
| D-Galactose | 11% | 81% | 8% | Stachyose | 96% | 3% | 1% |
| D-Galacturonic Acid | 88% | 3% | 10% | Sucrose | 10% | 83% | 7% |
| D-Gluconic Acid | 1% | 93% | 6% | Tween 40 | 10% | 63% | 28% |
| D-Glucose-6-PO4 | 3% | 97% | 0% | α-D-Glucose | 3% | 90% | 7% |
| D-Glucuronic Acid | 1% | 94% | 4% | α-D-Lactose | 93% | 0% | 7% |
| D-Lactic Acid Methyl Ester | 86% | 3% | 11% | α-Hydroxy-Butyric Acid | 19% | 64% | 17% |
| D-Malic Acid | 100% | 0% | 0% | α-Keto-Butyric Acid | 39% | 51% | 10% |
| D-Maltose | 3% | 83% | 14% | α-Keto-Glutaric Acid | 93% | 3% | 4% |
| D-Mannitol | 0% | 96% | 4% | β-Hydroxy-D,L-Butyric Acid | 100% | 0% | 0% |
| D-Mannose | 3% | 90% | 7% | β-Methyl-D-Glucoside | 75% | 21% | 4% |
| D-Melibiose | 99% | 0% | 1% | γ-Amino-Butryric Acid | 100% | 0% | 0% |
| D-Raffinose | 96% | 0% | 4% | ||||
| D-Saccharic Acid | 99% | 1% | 0% | Chemical Resistance | Negative | Positive | Intermediate |
| D-Salicin | 42% | 33% | 25% | 1% Sodium Lactate | 0% | 100% | 0% |
| D-Serine | 3% | 92% | 6% | 1% NaCl | 0% | 100% | 0% |
| D-Sorbitol | 17% | 75% | 8% | 4% NaCl | 0% | 100% | 0% |
| D-Trehalose | 3% | 85% | 13% | 8% NaCl | 0% | 100% | 0% |
| D-Turanose | 97% | 1% | 1% | Aztreonam | 47% | 51% | 1% |
| Formic Acid | 99% | 1% | 0% | D-Serine | 1% | 99% | 0% |
| Gelatin | 29% | 56% | 15% | Fusidic Acid | 0% | 100% | 0% |
| Gentiobiose | 99% | 1% | 0% | Guanidine HCl | 18% | 81% | 1% |
| Glucuronamide | 10% | 75% | 15% | Lincomycin | 53% | 39% | 8% |
| Glycerol | 0% | 99% | 1% | Lithium Chloride | 7% | 92% | 1% |
| Glycyl-L-Proline | 0% | 96% | 4% | Minocycline | 99% | 0% | 1% |
| Inosine | 3% | 94% | 3% | Nalidixic Acid | 99% | 0% | 1% |
| L-Alanine | 0% | 97% | 3% | Niaproof 4 | 0% | 99% | 1% |
| L-Arginine | 44% | 42% | 14% | pH 5 | 85% | 14% | 1% |
| L-Aspartic Acid | 1% | 96% | 3% | pH 6 | 0% | 100% | 0% |
| L-Fucose | 61% | 28% | 11% | Potassium Tellurite | 94% | 4% | 1% |
| L-Galactonic Acid Lactone | 100% | 0% | 0% | Rifamycin SV | 0% | 99% | 1% |
| L-Glutamic Acid | 0% | 97% | 3% | Sodium Broma | 75% | 25% | 0% |
| L-Histidine | 17% | 76% | 7% | Sodium Butyrate | 67% | 17% | 17% |
| L-Lactic Acid | 0% | 97% | 3% | Tetrazolium Blue | 0% | 100% | 0% |
| L-Malic Acid | 3% | 92% | 6% | Tetrazolium Violet | 75% | 22% | 3% |
| L-Pyroglutamic Acid | 100% | 0% | 0% | Troleandomycin | 7% | 92% | 1% |
| L-Rhamnose | 99% | 0% | 1% | Vancomycin | 3% | 97% | 0% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Triga, A.; Smyrli, M.; Katharios, P. Pathogenic and Opportunistic Vibrio spp. Associated with Vibriosis Incidences in the Greek Aquaculture: The Role of Vibrio harveyi as the Principal Cause of Vibriosis. Microorganisms 2023, 11, 1197. https://doi.org/10.3390/microorganisms11051197
Triga A, Smyrli M, Katharios P. Pathogenic and Opportunistic Vibrio spp. Associated with Vibriosis Incidences in the Greek Aquaculture: The Role of Vibrio harveyi as the Principal Cause of Vibriosis. Microorganisms. 2023; 11(5):1197. https://doi.org/10.3390/microorganisms11051197
Chicago/Turabian StyleTriga, Adriana, Maria Smyrli, and Pantelis Katharios. 2023. "Pathogenic and Opportunistic Vibrio spp. Associated with Vibriosis Incidences in the Greek Aquaculture: The Role of Vibrio harveyi as the Principal Cause of Vibriosis" Microorganisms 11, no. 5: 1197. https://doi.org/10.3390/microorganisms11051197
APA StyleTriga, A., Smyrli, M., & Katharios, P. (2023). Pathogenic and Opportunistic Vibrio spp. Associated with Vibriosis Incidences in the Greek Aquaculture: The Role of Vibrio harveyi as the Principal Cause of Vibriosis. Microorganisms, 11(5), 1197. https://doi.org/10.3390/microorganisms11051197