

Prevalence and Antibiotic Resistance of Bacillus sp. Isolated from Raw Milk

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation of Bacillus spp. Strains
2.2. Identification by MALDI-TOF
2.3. Phenotypic Antibiotic Resistance Analysis
2.4. Statistical Analysis
3. Results
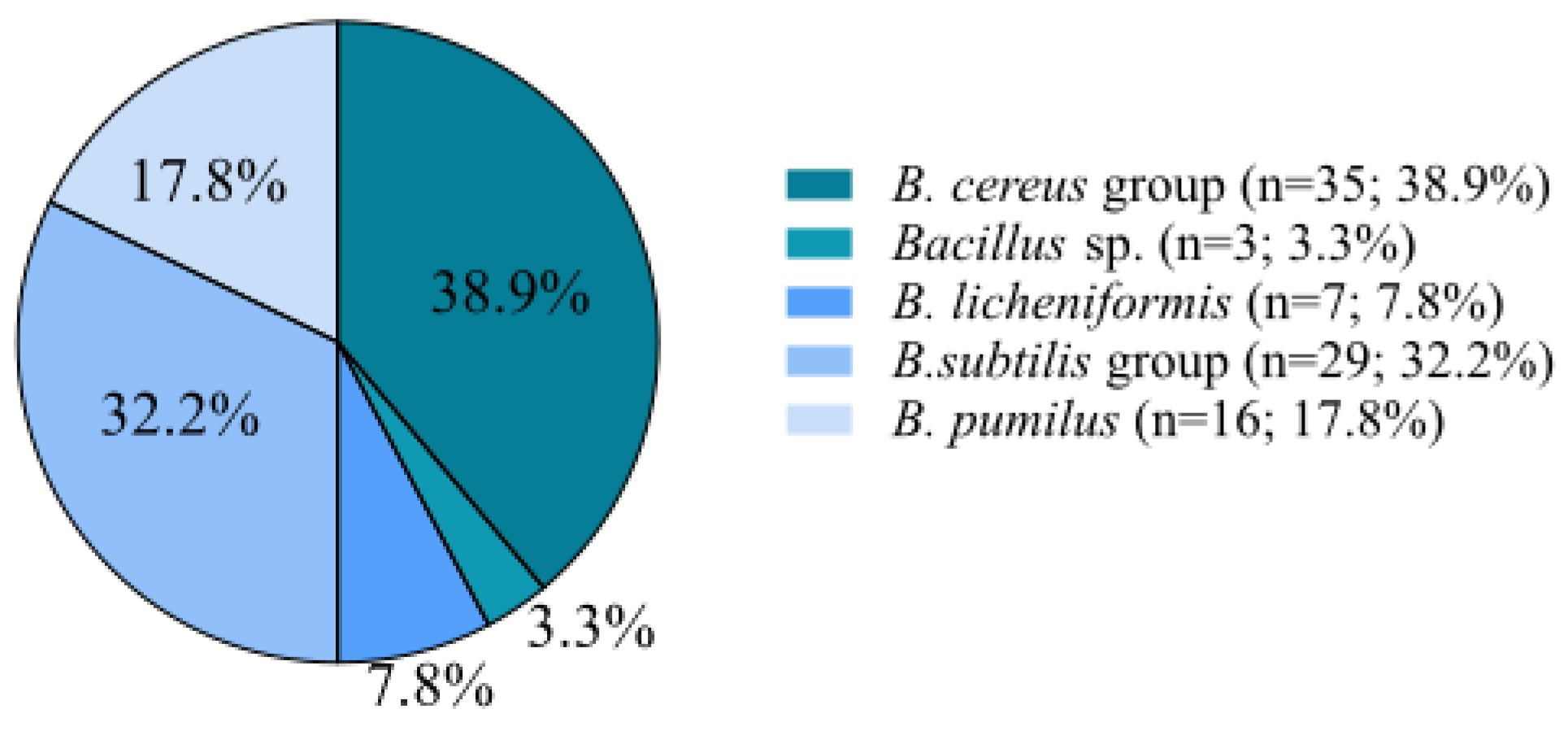
3.1. Isolation and Identification of Bacillus spp.
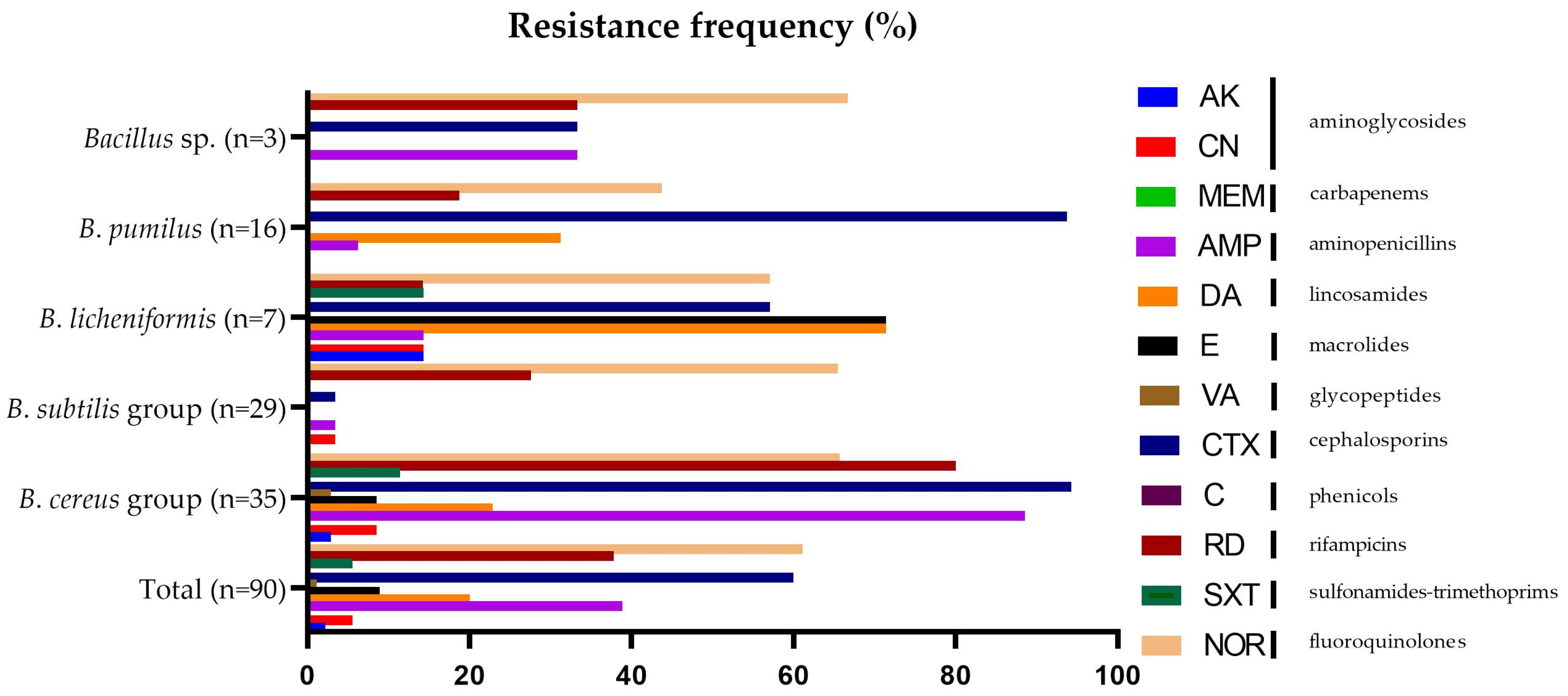
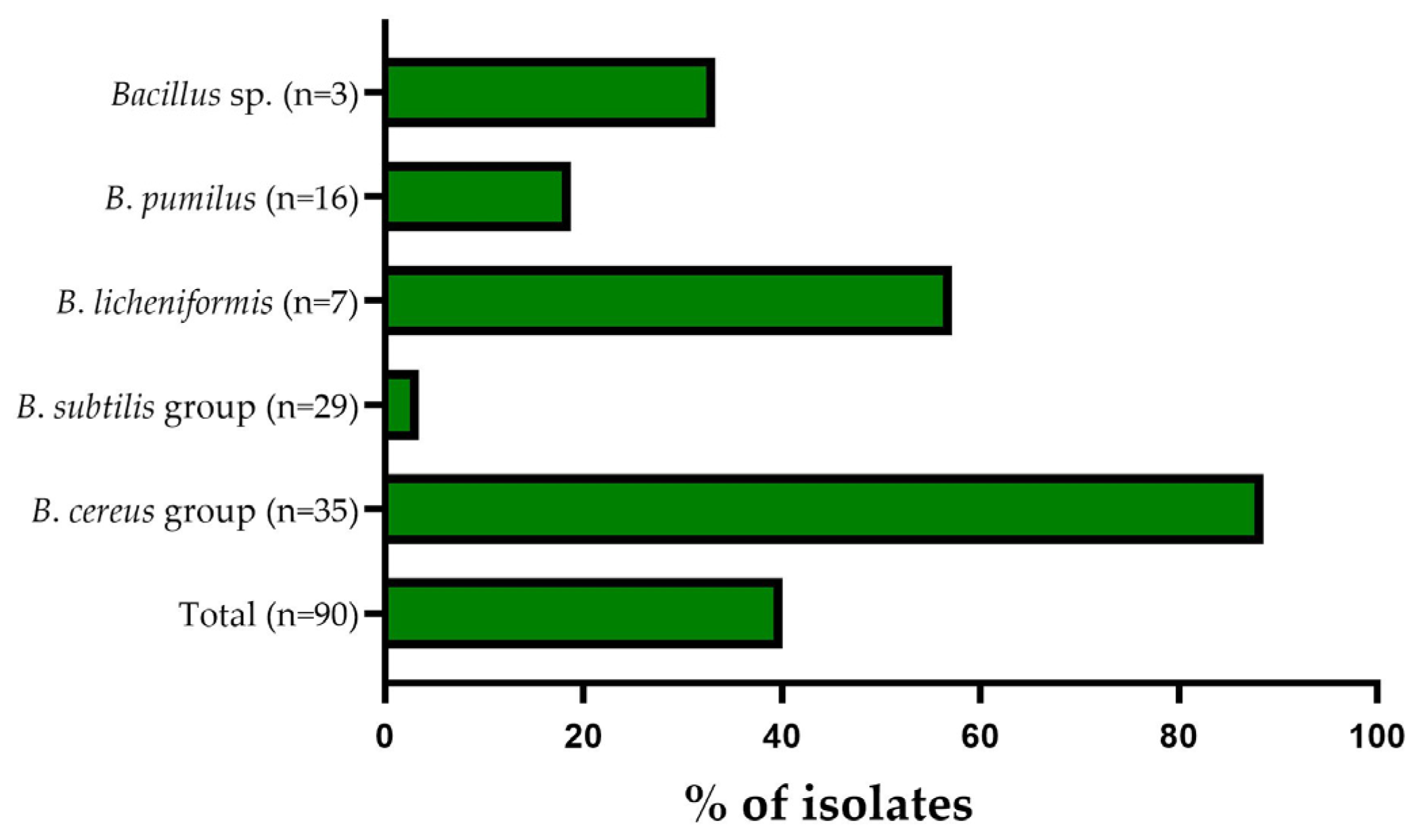
3.2. Antibiotic Resistance Pattern
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Von Neubeck, M.; Baur, C.; Krewinkel, M.; Stoeckel, M.; Kranz, B.; Stressler, T.; Fischer, L.; Hinrichs, J.; Scherer, S.; Wenning, M. Biodiversity of refrigerated raw milk microbiota and their enzymatic spoilage potential. Int. J. Food Microbiol. 2015, 211, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Velázquez-Ordoñez, V.; Valladares-Carranza, B.; Tenorio-Borroto, E.; Talavera-Rojas, M.; Antonio Varela-Guerrero, J.; Acosta-Dibarrat, J.; Puigvert, F.; Grille, L.; González Revello, Á.; Pareja, L. Microbial Contamination in Milk Quality and Health Risk of the Consumers of Raw Milk and Dairy Products. In Nutrition in Health and Disease—Our Challenges Now and Forthcoming Time; BoD—Books on Demand: Paris, France, 2019. [Google Scholar]
- Adamiak, A.; Górska, A.; Mróz, B. Bakterie psychrotrofowe w mleku surowym i jego przetworach. Zywn. Nauk. Technol. Jakosc/Food. Sci. Technol. Qual. 2015, 22, 36–48. [Google Scholar] [CrossRef]
- Yuan, H.; Han, S.; Zhang, S.; Xue, Y.; Zhang, Y.; Lu, H.; Wang, S. Microbial Properties of Raw Milk throughout the Year and Their Relationships to Quality Parameters. Foods 2022, 11, 3077. [Google Scholar] [CrossRef] [PubMed]
- Commission, T.H.E.; The, O.F.; Communities, E. European Commission Regulation (EC) no. 1662/2006 amending Regulation no. 853/2004. Regulation 2006, 2005, L320/0–L320/10. [Google Scholar]
- Wysok, B.; Wiszniewska-Łaszczych, A.; Uradziński, J.; Szteyn, J. Prevalence and antimicrobial resistance of Campylobacter in raw milk in the selected areas of Poland. Pol. J. Vet. Sci. 2011, 14, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Rolbiecki, D.; Harnisz, M. Mikroorganizmy lekooporne w środowisku przemysłu mleczarskiego. Pol. Dairy J. 2022, 10, 22–29. [Google Scholar]
- Lucey, J.A. Raw Milk Consumption: Risks and Benefits. Nutr. Today 2015, 50, 189–193. [Google Scholar] [CrossRef]
- De Jonghe, V.; Coorevits, A.; Vandroemme, J.; Heyrman, J.; Herman, L.; De Vos, P.; Heyndrickx, M. Intraspecific genotypic diversity of Bacillus species from raw milk. Int. Dairy J. 2008, 18, 496–505. [Google Scholar] [CrossRef]
- Hutchings, C.; Rajasekharan, S.K.; Reifen, R.; Shemesh, M. Mitigating milk-associated bacteria through inducing zinc ions antibiofilm activity. Foods 2020, 9, 1094. [Google Scholar] [CrossRef]
- Lücking, G.; Stoeckel, M.; Atamer, Z.; Hinrichs, J.; Ehling-Schulz, M. Characterization of aerobic spore-forming bacteria associated with industrial dairy processing environments and product spoilage. Int. J. Food Microbiol. 2013, 166, 270–279. [Google Scholar] [CrossRef]
- Granum, P.E. Spotlight on Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Lett. 2017, 364, fnx071. [Google Scholar] [CrossRef] [PubMed]
- Berthold-Pluta, A.; Pluta, A.; Garbowska, M.; Stefańska, I. Prevalence and Toxicity Characterization of Bacillus cereus in Food Products from Poland. Foods 2019, 8, 269. [Google Scholar] [CrossRef] [PubMed]
- Webb, M.D.; Barker, G.C.; Goodburn, K.E.; Peck, M.W. Risk presented to minimally processed chilled foods by psychrotrophic Bacillus cereus. Trends Food Sci. Technol. 2019, 93, 94–105. [Google Scholar] [CrossRef] [PubMed]
- European Union. The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, T. 19, e06971. [Google Scholar]
- Dubey, S.; Sharma, N.; Thakur, S.; Patel, R.; Reddy, B.M. Bacillus cereus food poisoning in Indian perspective: A review. J. Food Sci. Technol. 2021, 10, 970–975. [Google Scholar]
- Dietrich, R.; Jessberger, N.; Ehling-Schulz, M.; Märtlbauer, E.; Granum, P.E. The Food Poisoning Toxins of Bacillus cereus. Toxins 2021, 13, 98. [Google Scholar] [CrossRef]
- Jovanovic, J.; Ornelis, V.F.M.; Madder, A.; Rajkovic, A. Bacillus cereus food intoxication and toxicoinfection. Compr. Rev. Food Sci. Food Saf. 2021, 20, 3719–3761. [Google Scholar] [CrossRef]
- Stenfors Arnesen, L.P.; Fagerlund, A.; Granum, P.E. From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Rev. 2008, 32, 579–606. [Google Scholar] [CrossRef]
- Wash, P.; Batool, A.; Mulk, S.; Nazir, S.; Yasmin, H.; Mumtaz, S.; Alyemeni, M.N.; Kaushik, P.; Hassan, M.N. Prevalence of Antimicrobial Resistance and Respective Genes among Bacillus spp., a Versatile Bio-Fungicide. Int. J. Environ. Res. Public Health 2022, 19, 14997. [Google Scholar] [CrossRef]
- Zablotni, A.; Jaworski, A. Źródla Antybiotyków W SRodowiskach Naturalnych I Ich Rola Biologiczna. Postepy Hig. Med. Dosw. 2014, 68, 1040–1049. [Google Scholar] [CrossRef]
- Chajęcka-Wierzchowska, W.; Zadernowska, A.; Zarzecka, U.; Zakrzewski, A.; Gajewska, J. Enterococci from ready-to-eat food—horizontal gene transfer of antibiotic resistance genes and genotypic characterization by PCR melting profile. J. Sci. Food Agric. 2019, 99, 1172–1179. [Google Scholar] [CrossRef] [PubMed]
- Wall, B.A.; Mateus, A.; Marshall, L.; Pfeiffer, D.; Lubroth, J.; Ormel, H.J.; Otto, P.; Patriarchi, A. The Emergence of Antimicrobial Resistance in Bacteria. In Drivers, Dynamics and Epidemiology of Antimicrobial Resistance in Animal Production; Veterinary Epidemiology, Economics and Public Health Group, Department of Production and Population Health, The Royal Veterinary College: London, UK, 2016; ISBN 9789251094419. [Google Scholar]
- Byczkowska-Rostkowska, Z.; Gajewska, J.; Chajęcka-Wierzchowska, W. Antybiotyki i antybiotykooporność w przemyśle mleczarskim. Pol. Dairy J. 2022, 12, 10–15. [Google Scholar]
- Roca, I.; Akova, M.; Baquero, F.; Carlet, J.; Cavaleri, M.; Coenen, S.; Cohen, J.; Findlay, D.; Gyssens, I.; Heuer, O.E.; et al. The global threat of antimicrobial resistance: Science for intervention. New Microbes New Infect. 2015, 6, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Tóth, A.G.; Csabai, I.; Krikó, E.; Tőzsér, D.; Maróti, G.; Patai, Á.V.; Makrai, L.; Szita, G.; Solymosi, N. Antimicrobial resistance genes in raw milk for human consumption. Sci. Rep. 2020, 10, 7464. [Google Scholar] [CrossRef] [PubMed]
- Danilenko, V.; Devyatkin, A.; Marsova, M.; Shibilova, M.; Ilyasov, R.; Shmyrev, V. Common inflammatory mechanisms in covid-19 and Parkinson’s diseases: The role of microbiome, pharmabiotics and postbiotics in their prevention. J. Inflamm. Res. 2021, 14, 6349–6381. [Google Scholar] [CrossRef]
- Rahnama, H.; Azari, R.; Yousefi, M.H.; Berizi, E.; Mazloomi, S.M.; Hosseinzadeh, S.; Derakhshan, Z.; Ferrante, M.; Conti, G.O. A systematic review and meta-analysis of the prevalence of Bacillus cereus in foods. Food Control 2023, 143, 109250. [Google Scholar] [CrossRef]
- Zakrzewski, A.J.; Zarzecka, U.; Chajęcka-Wierzchowska, W.; Zadernowska, A. A Comparison of Methods for Identifying Enterobacterales Isolates from Fish and Prawns. Pathogens 2022, 11, 410. [Google Scholar] [CrossRef]
- Chajęcka-Wierzchowska, W.; Gajewska, J.; Zadernowska, A.; Randazzo, C.L.; Caggia, C. A Comprehensive Study on Antibiotic Resistance among Coagulase-Negative Staphylococci (CoNS) Strains Isolated from Ready-to-Eat Food Served in Bars and Restaurants. Foods 2023, 12, 514. [Google Scholar] [CrossRef]
- EUCAST: Breaking Point Tables for Interpretation of MICs and Zone Diameters v_12.0_Breakpoint_Tables. 2022, 1–110. Available online: http://www.eucast.org/clinical_breakpoints/ (accessed on 20 February 2023).
- Wiśniewski, P.; Zakrzewski, A.J.; Zadernowska, A.; Chajęcka-Wierzchowska, W. Antimicrobial Resistance and Virulence Characterization of Listeria monocytogenes Strains Isolated from Food and Food Processing Environments. Pathogens 2022, 11, 1099. [Google Scholar] [CrossRef]
- Gajewska, J.; Chajęcka-Wierzchowska, W.; Zadernowska, A. Occurrence and Characteristics of Staphylococcus aureus Strains along the Production Chain of Raw Milk Cheeses in Poland. Molecules 2022, 27, 6569. [Google Scholar] [CrossRef]
- Grutsch, A.A.; Nimmer, P.S.; Pittsley, R.H.; McKillip, J.L. Bacillus spp. as Pathogens in the Dairy Industry; Elsevier Inc.: Amsterdam, The Netherlands, 2018; ISBN 9780128114445. [Google Scholar]
- Gopal, N.; Hill, C.; Ross, P.R.; Beresford, T.P.; Fenelon, M.A.; Cotter, P.D. The prevalence and control of Bacillus and related spore-forming bacteria in the dairy industry. Front. Microbiol. 2015, 6, 1418. [Google Scholar] [CrossRef] [PubMed]
- Tirloni, E.; Stella, S.; Celandroni, F.; Mazzantini, D.; Bernardi, C.; Ghelardi, E. Bacillus cereus in Dairy Products and Production Plants. Foods 2022, 11, 2572. [Google Scholar] [CrossRef] [PubMed]
- Elegbeleye, J.A.; Buys, E.M. Potential spoilage of extended shelf-life (ESL) milk by Bacillus subtilis and Bacillus velezensis. LWT 2022, 153, 112487. [Google Scholar] [CrossRef]
- Elshaghabee, F.M.F.; Rokana, N.; Gulhane, R.D.; Sharma, C.; Panwar, H. Bacillus as potential probiotics: Status, concerns, and future perspectives. Front. Microbiol. 2017, 8, 1490. [Google Scholar] [CrossRef]
- Hornik, B.; Czarny, J.; Staninska-Pięta, J.; Wolko, Ł.; Cyplik, P.; Piotrowska-Cyplik, A. The raw milk microbiota from semi-subsistence farms characteristics by NGS analysis method. Molecules 2021, 26, 5029. [Google Scholar] [CrossRef]
- Li, F.; Hunt, K.; Van Hoorde, K.; Butler, F.; Jordan, K.; Tobin, J.T. Occurrence and identification of spore-forming bacteria in skim-milk powders. Int. Dairy J. 2019, 97, 176–184. [Google Scholar] [CrossRef]
- Rückert, A.; Ronimus, R.S.; Morgan, H.W. A RAPD-based survey of thermophilic bacilli in milk powders from different countries. Int. J. Food Microbiol. 2004, 96, 263–272. [Google Scholar] [CrossRef]
- Heyndrickx, M.; Scheldeman, P. Bacilli Associated with Spoilage in Dairy Products and Other Food. Appl. Syst. Bacillus Relat. 2008, 64–82. [Google Scholar] [CrossRef]
- Kumari, S.; Sarkar, P.K. Prevalence and characterization of Bacillus cereus group from various marketed dairy products in India. Dairy Sci. Technol. 2014, 94, 483–497. [Google Scholar] [CrossRef]
- World Health Organization. The World is Running Out of Antibiotics, WHO Report Confirms. 2017. Available online: https://www.who.int/news/item/20-09-2017-the-world-is-running-out-of-antibiotics-who-report-confirms (accessed on 22 May 2022).
- Gundogan, N.; Avci, E. Occurrence and antibiotic resistance of Escherichia coli, Staphylococcus aureus and Bacillus cereus in raw milk and dairy products in Turkey. Int. J. Dairy Technol. 2014, 67, 562–569. [Google Scholar] [CrossRef]
- Chang, Y.; Xie, Q.; Yang, J.; Ma, L.; Feng, H. The prevalence and characterization of Bacillus cereus isolated from raw and pasteurized buffalo milk in southwestern China. J. Dairy Sci. 2021, 104, 3980–3989. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Fang, Y.; Zhu, J.; Xu, W.; Zhu, K. Characterization of Bacillus species from market foods in Beijing, China. Processes 2021, 9, 866. [Google Scholar] [CrossRef]
- Kong, L.; Yu, S.; Yuan, X.; Li, C.; Yu, P.; Wang, J.; Guo, H.; Wu, S.; Ye, Q.; Lei, T.; et al. An Investigation on the Occurrence and Molecular Characterization of Bacillus cereus in Meat and Meat Products in China. Foodborne Pathog. Dis. 2021, 18, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Babich, O.O.; Sukhikh, S.A.; Zimina, M.I.; Milentyeva, I.S. Antibiotic activity and resistance of lactic acid bacteria and other antagonistic bacteriocin-producing microorganisms. Foods Raw Mater. 2020, 8, 377–384. [Google Scholar] [CrossRef]
- Pasvolsky, R.; Zakin, V.; Ostrova, I.; Shemesh, M. Butyric acid released during milk lipolysis triggers biofilm formation of Bacillus species. Int. J. Food Microbiol. 2014, 181, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Z.; Cui, C.; Li, X.; Yan, J.; Sun, E.; Wang, C.; Guo, H.; Hao, Y. Prevalence, antimicrobial susceptibility, and antibiotic resistance gene transfer of Bacillus strains isolated from pasteurized milk. J. Dairy Sci. 2023, 106, 75–83. [Google Scholar] [CrossRef]
- Jeong, D.W.; Lee, B.; Heo, S.; Oh, Y.; Heo, G.; Lee, J.H. Two genes involved in clindamycin resistance of Bacillus licheniformis and Bacillus paralicheniformis identified by comparative genomic analysis. PLoS ONE 2020, 15, e0231274. [Google Scholar] [CrossRef]
- Awasti, N.; Anand, S.; Djira, G. Sporulating behavior of Bacillus licheniformis strains influences their population dynamics during raw milk holding. J. Dairy Sci. 2019, 102, 6001–6012. [Google Scholar] [CrossRef]
- Manzulli, V.; Rondinone, V.; Buchicchio, A.; Serrecchia, L.; Cipolletta, D.; Fasanella, A.; Parisi, A.; Difato, L.; Iatarola, M.; Aceti, A.; et al. Discrimination of Bacillus cereus group members by maldi-tof mass spectrometry. Microorganisms 2021, 9, 1202. [Google Scholar] [CrossRef]
- Liu, Y.; Lai, Q.; Shao, Z. Genome analysis-based reclassification of Bacillus weihenstephanensis as a later heterotypic synonym of Bacillus mycoides. Int. J. Syst. Evol. Microbiol. 2018, 68, 106–112. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| No. (%) of Isolates | Antibiotic Profiles | MAR * Index | |
|---|---|---|---|
| B. cereus group (n = 35) | 1 (2.9%) | NOR | 0.08 |
| 1 (2.9%) | RD, NOR | 0.17 | |
| 2 (5.7%) | AMP, CTX | 0.17 | |
| 1 (2.9%) | AMP, CTX, NOR | 0.25 | |
| 8 (22.9%) | AMP, CTX, RD | 0.25 | |
| 1 (2.9%) | DA, CTX, NOR | 0.25 | |
| 10 (28.6%) | AMP, CTX, RD, NOR | 0.33 | |
| 2 (5.7%) | AMP, CTX, RD, SXT | 0.33 | |
| 1 (2.9%) | AMP, DA, CTX, NOR | 0.33 | |
| 1 (2.9%) | AMP, CTX, RD, SXT, NOR | 0.42 | |
| 1 (2.9%) | CN, DA, E, CTX, NOR | 0.42 | |
| 2 (5.7%) | AMP, DA, CTX, RD, NOR | 0.42 | |
| 1 (2.9%) | AMP, DA, CTX, RD, SXT, NOR | 0.5 | |
| 1 (2.9%) | CN, AMP, DA, CTX, RD, NOR | 0.5 | |
| 1 (2.9%) | CN, AMP, E, CTX, RD, NOR | 0.5 | |
| 1 (2.9%) | AK, AMP, DA, E, VA, CTX, RD, NOR | 0.67 | |
| B. licheniformis (n = 7) | 2 (28.6%) | DA, NOR | 0.17 |
| 1 (14.3%) | DA, CTX, NOR | 0.25 | |
| 1 (14.3%) | DA, E, CTX | 0.25 | |
| 1 (14.3%) | AK, DA, E, CTX | 0.33 | |
| 1 (14.3%) | CN, AMP, CTX, RD, SXT, NOR | 0.5 | |
| 1 (14.3%) | - | - | |
| B. pumilus (n = 16) | 5 (31.3%) | CTX | 0.08 |
| 4 (25.0%) | CTX, NOR | 0.17 | |
| 2 (12.5%) | DA, CTX | 0.17 | |
| 1 (6.3%) | AMP, CTX, RD | 0.25 | |
| 1 (6.3%) | DA, CTX, NOR | 0.25 | |
| 2 (12.5%) | DA, CTX, RD, NOR | 0.33 | |
| 1 (6.3%) | - | - | |
| B. subtilis group (n = 29) | 12 (41.4%) | NOR | 0.08 |
| 1 (3.4%) | RD | 0.08 | |
| 6 (20.7%) | RD, NOR | 0.17 | |
| 1 (3,4%) | CN, NOR | 0.17 | |
| 1 (3.4%) | AMP, CTX, RD | 0.25 | |
| 8 (27.6%) | - | - | |
| Bacillus sp. (n = 3) | 1 (33.3%) | NOR | 0.08 |
| 1 (33.3%) | AMP, CTX, RD, NOR | 0.33 | |
| 1 (33.3%) | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adamski, P.; Byczkowska-Rostkowska, Z.; Gajewska, J.; Zakrzewski, A.J.; Kłębukowska, L. Prevalence and Antibiotic Resistance of Bacillus sp. Isolated from Raw Milk. Microorganisms 2023, 11, 1065. https://doi.org/10.3390/microorganisms11041065
Adamski P, Byczkowska-Rostkowska Z, Gajewska J, Zakrzewski AJ, Kłębukowska L. Prevalence and Antibiotic Resistance of Bacillus sp. Isolated from Raw Milk. Microorganisms. 2023; 11(4):1065. https://doi.org/10.3390/microorganisms11041065
Chicago/Turabian StyleAdamski, Patryk, Zuzanna Byczkowska-Rostkowska, Joanna Gajewska, Arkadiusz Józef Zakrzewski, and Lucyna Kłębukowska. 2023. "Prevalence and Antibiotic Resistance of Bacillus sp. Isolated from Raw Milk" Microorganisms 11, no. 4: 1065. https://doi.org/10.3390/microorganisms11041065
APA StyleAdamski, P., Byczkowska-Rostkowska, Z., Gajewska, J., Zakrzewski, A. J., & Kłębukowska, L. (2023). Prevalence and Antibiotic Resistance of Bacillus sp. Isolated from Raw Milk. Microorganisms, 11(4), 1065. https://doi.org/10.3390/microorganisms11041065