

Strain Streptomyces sp. P-56 Produces Nonactin and Possesses Insecticidal, Acaricidal, Antimicrobial and Plant Growth-Promoting Traits

 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Microorganism
2.2. Identification of Strain P-56
2.3. Obtaining Laboratory Sample of the Metabolic Complex
2.4. Insecticidal Activity of DEE
2.5. Isolation and Purification of the Toxin
2.6. Identification of the Toxin
2.7. Method of Growing Crystals for X-ray Analysis
2.8. Acaricidal Activity of DEE
2.9. Antifungal and Antibacterial Activity of Strain P-56
2.10. Acetylene Reduction Activity of Strain P-56
2.11. Phytohormone Production by Strain P-56
2.12. Utilization of ACC by Strain P-56
2.13. Solubilization of Phosphates by Strain P-56
2.14. Production of Siderophores by Strain P-56
2.15. Bioassay with Plants
2.16. Statistical Analysis
3. Results
3.1. Identification of Strain P-56
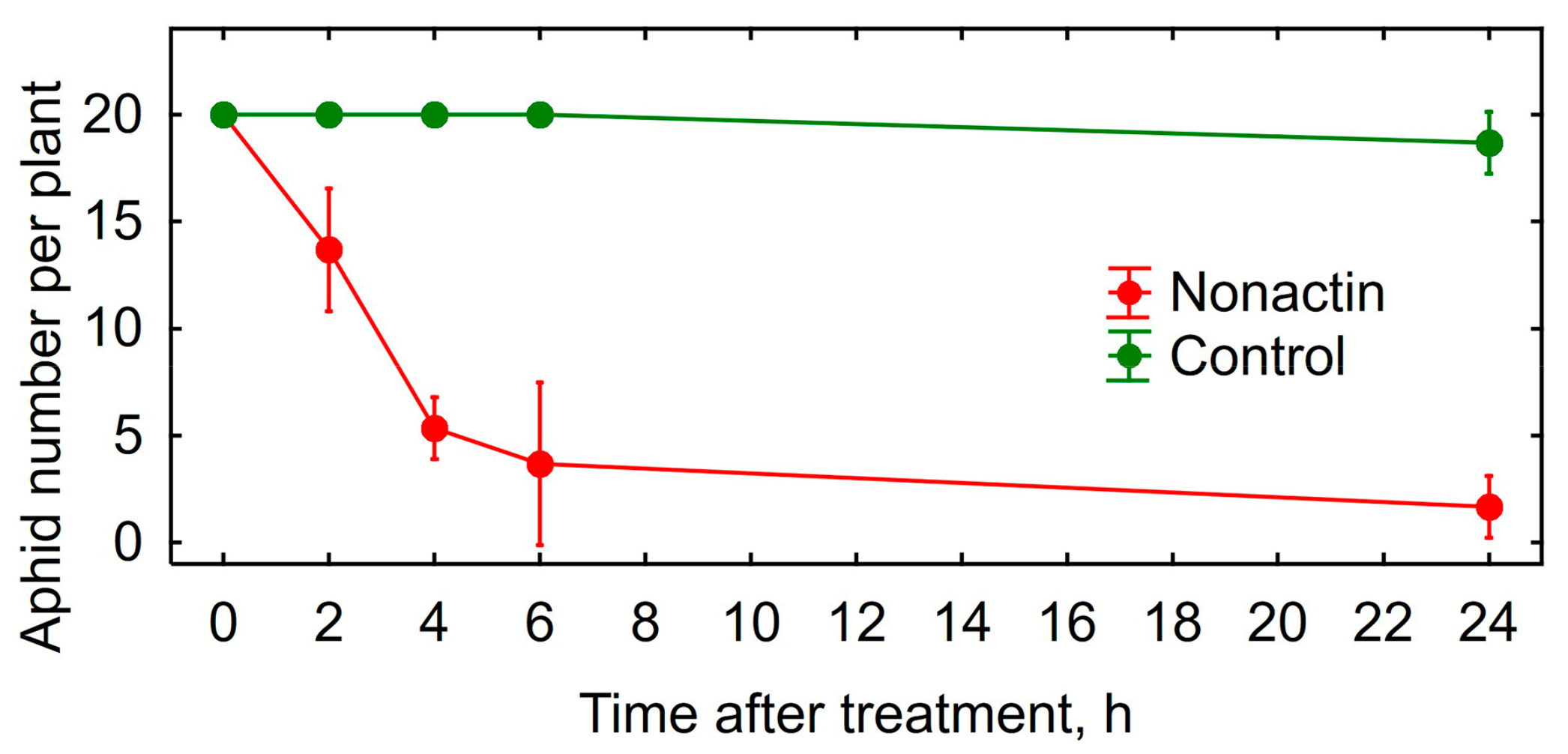
3.2. Insecticidal Activity
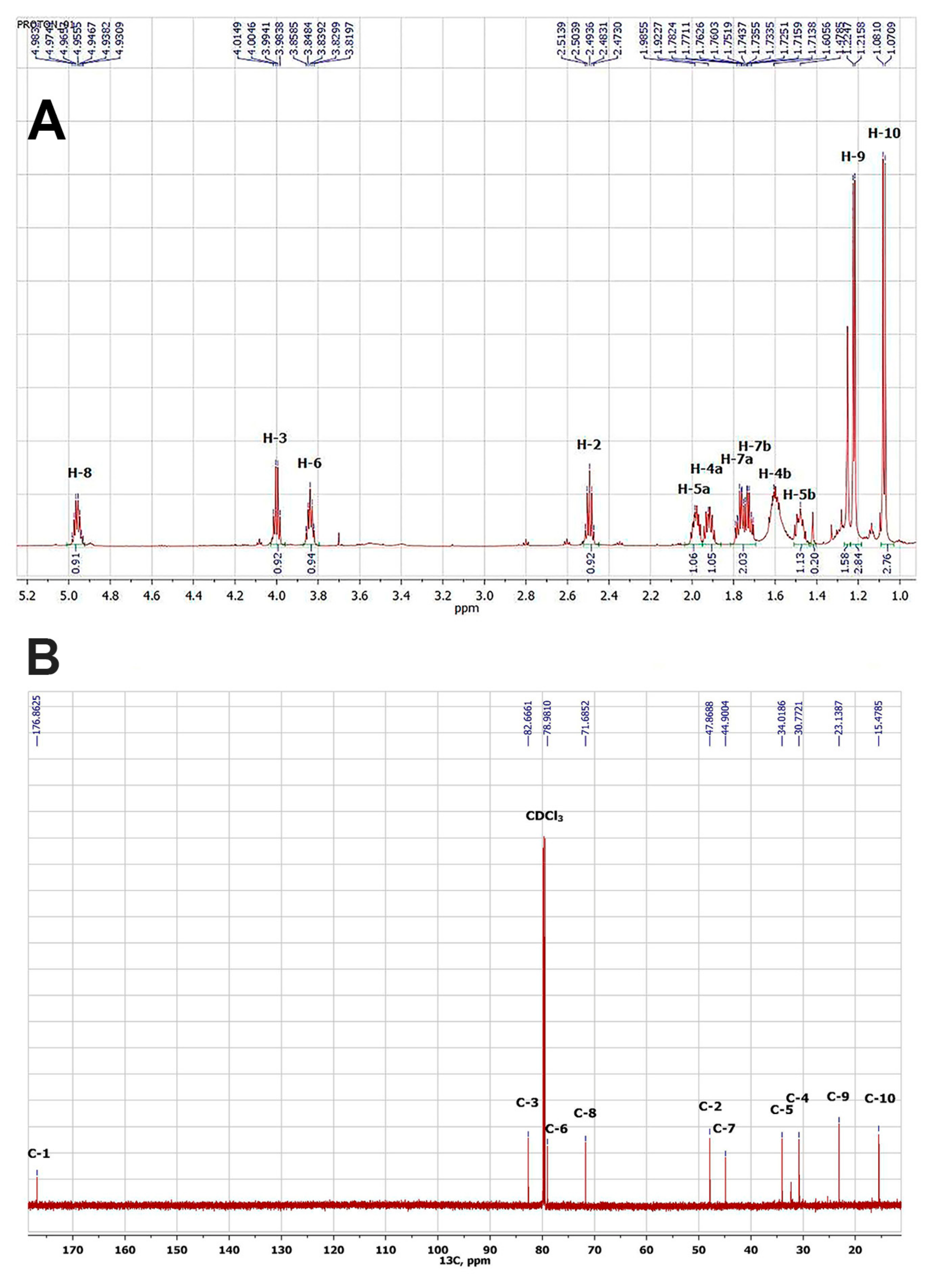
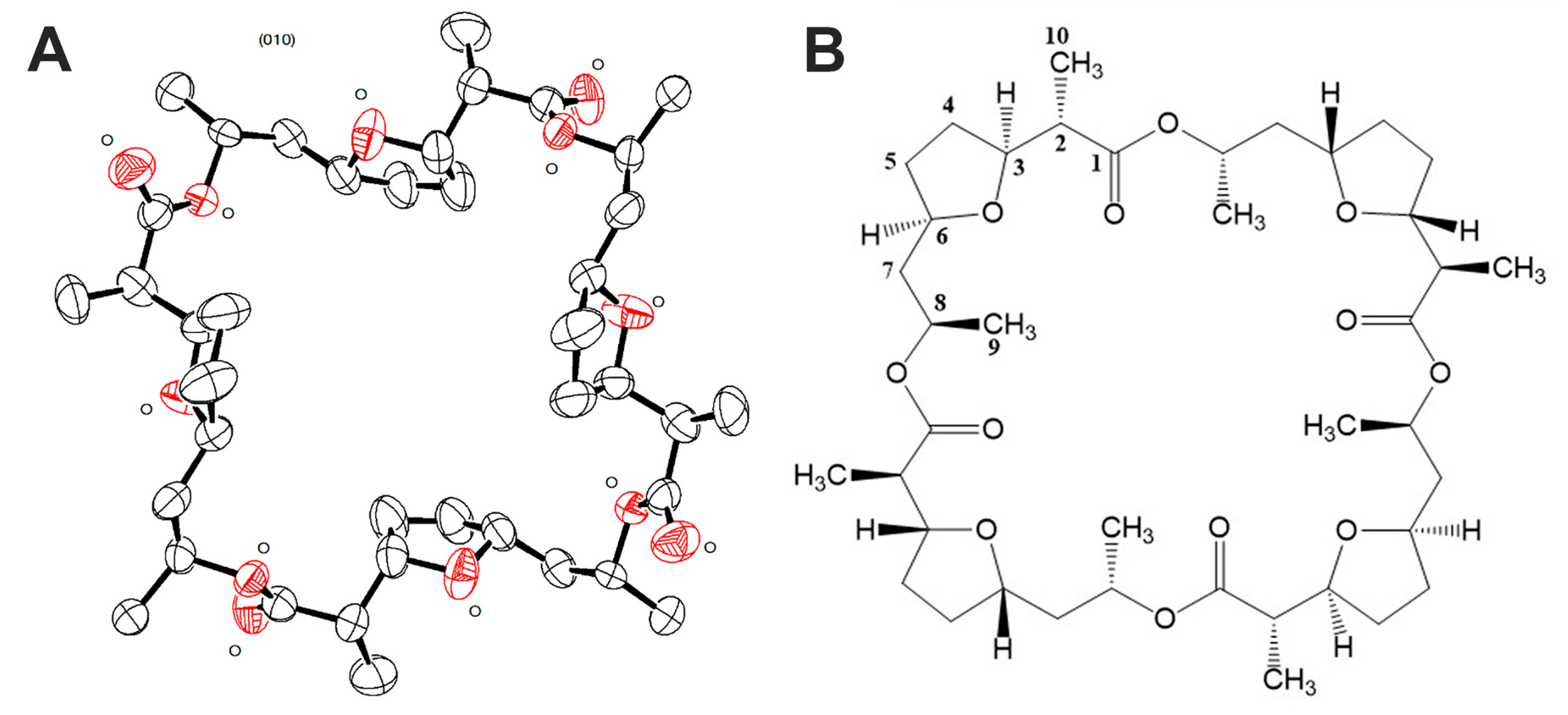
3.3. Isolation and Structural Identification of the Active Substance
3.4. Acaricidal Activity
3.5. Antagonistic Properties
3.6. Plant Growth-Promoting Traits
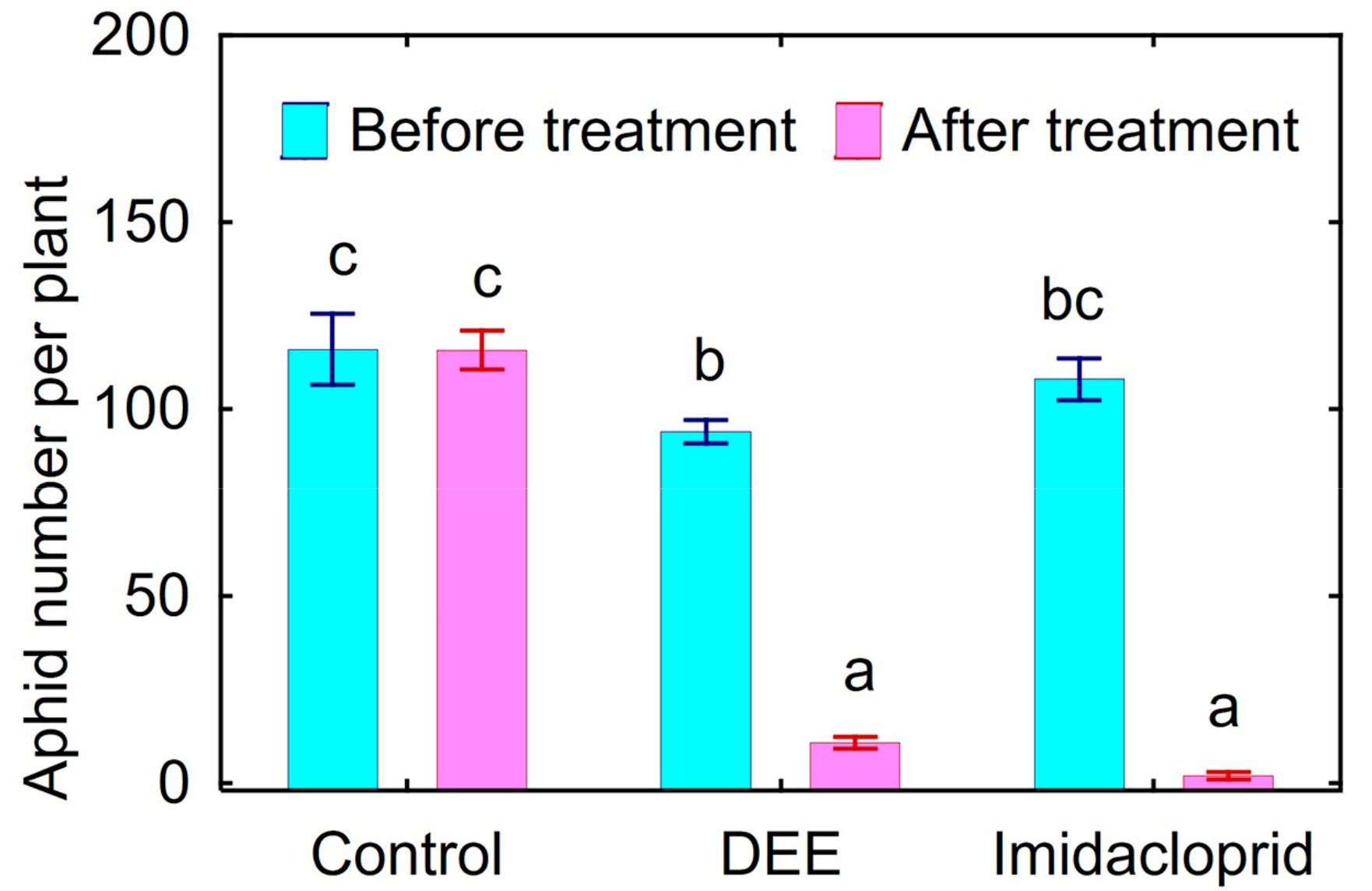
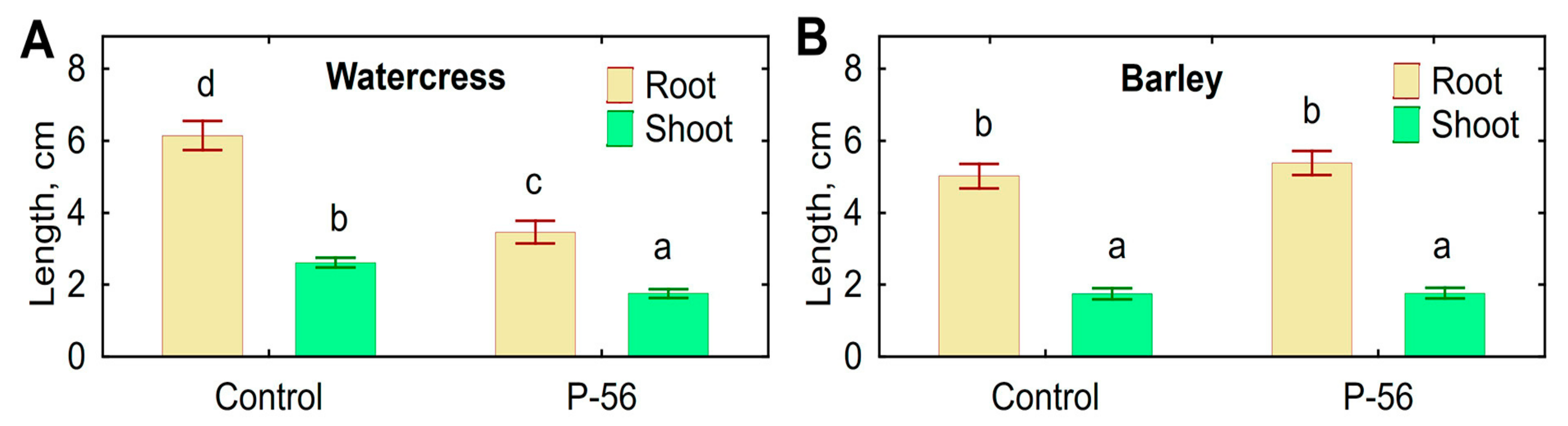
3.7. Biotests with Plants
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maithani, D.; Sharma, A.; Gangola, S.; Choudhary, P.; Bhatt, P. Insights into applications and strategies for discovery of microbial bioactive metabolites. Microbiol. Res. 2022, 261, 127053. [Google Scholar] [CrossRef] [PubMed]
- Ossai, J.; Khatabi, B.; Nybo, S.E.; Kharel, M.K. Renewed interests in the discovery of bioactive actinomycete metabolites driven by emerging technologies. J. Appl. Microbiol. 2022, 132, 59–77. [Google Scholar] [CrossRef] [PubMed]
- Yuzikhin, O.S.; Shaposhnikov, A.I.; Konnova, T.A.; Syrova, D.S.; Hamo, H.; Ermekkaliev, T.S.; Shevchenko, V.P.; Shevchenko, K.V.; Gogoleva, N.E.; Nizhnikov, A.A.; et al. Isolation and characterization of 1-Hydroxy-2,6,6-trimethyl-4-oxo-2-cyclohexene-1-acetic acid, a metabolite in bacterial transformation of abscisic acid. Biomolecules 2022, 12, 1508. [Google Scholar] [CrossRef] [PubMed]
- Copping, L.G.; Menn, J.J. Biopesticides: A review of their action, applications and efficacy. Pest. Manag. Sci. 2000, 56, 651–676. [Google Scholar] [CrossRef]
- Chattopadhyay, P.; Banerjee, G.; Mukherjee, S. Recent trends of modern bacterial insecticides for pest control practice in integrated crop management system. 3 Biotech 2017, 7, 60. [Google Scholar] [CrossRef]
- Dewi, T.K.; Agustiani, D.; Antonius, S. Secondary metabolites production by actinomycetes and their antifungal activity. KnE Life Sci. 2017, 3, 256. [Google Scholar] [CrossRef]
- Salwan, R.; Sharma, V. Molecular and biotechnological aspects of secondary metabolites in actinobacteria. Microbiol. Res. 2019, 231, 126374. [Google Scholar] [CrossRef]
- Mingbo, Q.U.; Merzendorfer, H.; Moussian, B.; Yang, Q. Bioinsecticides as future mainstream pest control agents: Opportunities and challenges. Fron. Agr Sci. Eng. 2022, 9, 82–97. [Google Scholar] [CrossRef]
- Zenova, G.M. Soil Actinomycetes; Publishing House of Moscow State University: Moscow, Russia, 1992; p. 78. ISBN 5-211-02902-X. (In Russian) [Google Scholar]
- Olanrewaju, O.S.; Babalola, O.O. Streptomyces: Implications and interactions in plant growth promotion. Appl. Microbiol. Biotechnol. 2019, 103, 1179–1188. [Google Scholar] [CrossRef]
- Zizka, Z. Biological effects of macrotetrolide antibiotics and nonactic acids. Folia Microbiol. 1998, 43, 7–14. [Google Scholar] [CrossRef]
- Watve, M.G.; Tickoo, R.; Jog, M.M.; Bhole, B.D. How many antibiotics are produced by the genus Streptomyces? Arch. Microbiol. 2001, 176, 386–390. [Google Scholar] [CrossRef]
- Degtyareva, E.A.; Vinogradova, K.A.; Alexandrova, A.V.; Filonenko, V.A.; Kozhevin, P.A. Soil streptomycetes as potential biofungicides. Moscow Univ. Soil Sci. Bull. 2009, 64, 73–77. [Google Scholar] [CrossRef]
- Waksman, S.A.; Schatz, A.; Reynods, D.M. Production of antibiotic substances by actinomycetes. Ann. N. Y. Acad. Sci. 2010, 1213, 112–124. [Google Scholar] [CrossRef]
- Procópio, R.E.; Silva, I.R.; Martins, M.K.; Azevedo, J.L.; Araújo, J.M. Antibiotics produced by Streptomyces. Braz. J. Infect. Dis. 2012, 16, 466–471. [Google Scholar] [CrossRef]
- Siegwart, M.; Graillot, B.; Blachere Lopez, C.; Besse, S.; Bardin, M.; Nicot, P.C.; Lopez-Ferber, M. Resistance to bio-insecticides or how to enhance their sustainability: A review. Front. Plant Sci. 2015, 6, 381. [Google Scholar] [CrossRef]
- Belakhov, V.V.; Boikova, I.V.; Novikova, I.I.; Kolodyaznya, V.A. Results of examination of the biological activity of nonmedical antibiotics with a view to finding environmentally friendly pesticides for plant protection. Russ. J. Gen. Chem. 2018, 88, 2982–2989. [Google Scholar] [CrossRef]
- Katarzyn, J.; Aexander, R.; Guspiel, A.; Ziemskaj, O.J. Secondary metabolites of actinomycetes and their antibacterial, antifungal and antiviral properties. Pol. J. Microbiol. 2018, 67, 259–272. [Google Scholar] [CrossRef]
- Pavlyusin, V.A.; Novikova, I.I.; Boikova, I.V. Microbiological control in phytosanitary optimization technologies for agroecosystems: Research and practice (review). Agric. Biol. 2020, 55, 421–438. [Google Scholar] [CrossRef]
- Jizba, J.; Sedmera, P.; Zima, J.; Beran, M.; Blumauerová, M.; Kandybin, N.V.; Samoukina, G.V. Macrotetrolide antibiotics produced by Streptomyces globisporus. Folia Microbiol. 1991, 36, 437–443. [Google Scholar] [CrossRef]
- Crevelin, E.J.; Crotti, A.E.; Zucchi, T.D.; Melo, I.S.; Moraes, L.A. Dereplication of Streptomyces sp. AMC 23 polyether ionophore antibiotics by accurate-mass electrospray tandem mass spectrometry. J. Mass Spectrom. 2014, 49, 1117–1126. [Google Scholar] [CrossRef]
- Harinantenaina Rakotondraibe, L.; Rasolomampianina, R.; Park, H.Y.; Li, J.; Slebodnik, C.; Brodie, P.J.; Blasiak, L.C.; Hill, R.; TenDyke, K.; Shen, Y.; et al. Antiproliferative and antiplasmodial compounds from selected Streptomyces species. Bioorg. Med. Chem. Lett. 2015, 25, 5646–5649. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Zheng, S. Efficient production of nonactin by Streptomyces griseus subsp. griseus. Can. J. Microbiol. 2016, 62, 711–714. [Google Scholar] [CrossRef] [PubMed]
- Kusche, B.R.; Phillips, J.B.; Priestley, N.D. Nonactin biosynthesis: Setting limits on what can be achieved with precursor-directed biosynthesis. Bioorg. Med. Chem. Lett. 2009, 19, 1233–1235. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhang, J.; He, J. Structures, synthesis and biological activities of nonactic acid and its derivatives. Curr. Med. Chem. 2021, 28, 8673–8691. [Google Scholar] [CrossRef]
- Jizba, J.; Přikrylová, V.; Ujhelyiová, L.; Varkonda, Š. Insecticidal properties of nonactic acid and homononactic acid, the precursors of macrotetrolide antibiotics. Folia Microbiol. 1992, 37, 299–303. [Google Scholar] [CrossRef]
- Jizba, J.; Skibová, I. Regulation of biosynthesis of pesticidal metabolic complexes in Streptomyces griseus. Folia Microbiol. 1994, 39, 119–128. [Google Scholar] [CrossRef]
- Kaaniche, F.; Hamed, A.; Elleuch, L.; Chakchouk-Mtibaa, A.; Smaoui, S.; Karray-Rebai, I.; Koubaa, I.; Arcile, G.; Allouche, N.; Mellouli, L. Purification and characterization of seven bioactive compounds from the newly isolated Streptomyces cavourensis TN638 strain via solid-state fermentation. Microb. Pathog. 2020, 142, 104106. [Google Scholar] [CrossRef]
- Schrey, S.D.; Tarkka, M.T. Friends and foes: Streptomycetes as modulators of plant disease and symbiosis. Antonie van Leeuwenhoek 2008, 94, 11–19. [Google Scholar] [CrossRef]
- Seipke, R.F.; Kaltenpoth, M.; Hutchings, M.I. Streptomyces as symbionts: An emerging and widespread theme? FEMS Microbiol. Rev. 2012, 36, 862–876. [Google Scholar] [CrossRef]
- Viaene, T.; Langendries, S.; Beirinckx, S.; Maes, M.; Goormachtig, S. Streptomyces as a plant’s best friend? FEMS Microbiol. Ecol. 2016, 92, fiw119. [Google Scholar] [CrossRef]
- Aldesuquy, H.S.; Mansour, F.A.; Abo-Hamed, S.A. Effect of the culture filtrates of Streptomyces on growth and productivity of wheat plants. Folia Microbiol. 1998, 43, 465–470. [Google Scholar] [CrossRef]
- Dimkpa, C.; Svatos, A.; Merten, D.; Büchel, G.; Kothe, E. Hydroxamate siderophores produced by Streptomyces acidiscabies E13 bind nickel and promote growth in cowpea (Vigna unguiculata L.) under nickel stress. Can. J. Microbiol. 2008, 54, 163–172. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Vadlamudi, S.; Bandikinda, P.; Sathya, A.; Vijayabharathi, R.; Rupela, O.; Kudapa, H.; Katta, K.; Varshney, R.K. Evaluation of Streptomyces strains isolated from herbal vermicompost for their plant growth-promotion traits in rice. Microbiol. Res. 2014, 169, 40–48. [Google Scholar] [CrossRef]
- Kaur, T.; Manhas, R.K. Antifungal, insecticidal, and plant growth promoting potential of Streptomyces hydrogenans DH16. J. Basic Microbiol. 2014, 54, 1175–1185. [Google Scholar] [CrossRef]
- Terra, L.; Ratcliffe, N.; Castro, H.C.; Vicente, A.C.P.; Dyson, P. Biotechnological potential of Streptomyces siderophores as new antibiotics. Curr. Med. Chem. 2021, 28, 1407–1421. [Google Scholar] [CrossRef]
- Jog, R.; Pandya, M.; Nareshkumar, G.; Rajkumar, S. Mechanism of phosphate solubilization and antifungal activity of Streptomyces spp. isolated from wheat roots and rhizosphere and their application in improving plant growth. Microbiology 2014, 160 Pt 4, 778–788. [Google Scholar] [CrossRef]
- Anwar, S.; Ali, B.; Sajid, I. Screening of rhizospheric actinomycetes for various in-vitro and in-vivo plant growth promoting (PGP) traits and for agroactive compounds. Front. Microbiol. 2016, 7, 1334. [Google Scholar] [CrossRef]
- Alwahshi, K.J.; Purayil, G.P.; Saeed, E.E.; Abufarajallah, H.A.; Aldhaheri, S.J.; AbuQamar, S.F.; El-Tarabily, K.A. The 1-aminocyclopropane-1-carboxylic acid deaminase-producing Streptomyces violaceoruber UAE1 can provide protection from sudden decline syndrome on date palm. Front. Plant Sci. 2022, 13, 904166. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Srinivas, V.; Chand, U.; Pratyusha, S.; Samineni, S. Streptomyces consortia-mediated plant growth-promotion and yield performance in chickpea. 3 Biotech 2022, 12, 318. [Google Scholar] [CrossRef]
- Holt, J.G. (Ed.) Bergey’s Mannual of Determinative Bacteriology, 9th ed.; Lippincott Williams and Wilkins: Baltimore, MD, USA, 1994; p. 787. ISBN 13 978-0683006032. [Google Scholar]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef]
- Safronova, V.; Belimov, A.; Sazanova, A.; Chirak, E.; Kuznetsova, I.; Andronov, E.; Pinaev, A.; Tsyganova, A.; Seliverstova, E.; Kitaeva, A.; et al. Two broad host range rhizobial strains isolated from relict legumes have various complementary effects on symbiotic parameters of co-inoculated plants. Front. Microbiol. 2019, 10, 514. [Google Scholar] [CrossRef] [PubMed]
- Dolomanov, O.V.; Bourhis, L.J.; Gildea, R.J.; Howard, J.A.; Puschmann, H. OLEX2: A complete structure solution, refinement and analysis program. J. Appl. Crystallogr. 2009, 42, 339–341. [Google Scholar] [CrossRef]
- Rennie, R.J. A single medium for the isolation of acetylene-reducing (nitrogen-fixing) bacteria from soils. Can. J. Microbiol. 1981, 27, 8–14. [Google Scholar] [CrossRef]
- Belimov, A.A.; Hontzeas, N.; Safronova, V.I.; Demchinskaya, S.V.; Piluzza, G.; Bullitta, S.; Glick, B.R. Cadmium-tolerant plant growth-promoting bacteria associated with the roots of Indian mustard (Brassica juncea L. Czern.). Soil Biol. Biochem. 2005, 37, 241–250. [Google Scholar] [CrossRef]
- Brazhnikova, Y.V.; Shaposhnikov, A.I.; Sazanova, A.L.; Belimov, A.A.; Mukasheva, T.D.; Ignatova, L.V. Phosphate mobilization by culturable fungi and their capacity to increase soil p availability and promote barley growth. Curr. Microbiol. 2022, 79, 240. [Google Scholar] [CrossRef] [PubMed]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Saitô, H.; Tabeta, R.; Yokoi, M. High-resolution 13C NMR study of free and metal-complexed ionophores in the solid state: Conformation and dynamics of the macrocyclic ring and the effect of intermolecular short contact of methyl groups on spin-lattice relaxation times and displacements of chemical shifts. Magn. Reson. Chem. 1988, 26, 775–786. [Google Scholar] [CrossRef]
- Grimme, S.; Bannwarth, C.; Dohm, S.; Hansen, A.; Pisarek, J.; Pracht, P.; Seibert, J.; Neese, F. Fully automated quantum-chemistry-based computation of spin–spin-coupled nuclear magnetic resonance spectra. Angew. Chem. Int. Ed. 2017, 56, 14763–14769. [Google Scholar] [CrossRef]
- Corbaz, R.; Ettlinger, L.; Gäumann, E.; Keller-Schierlein, W.; Kradolfer, F.; Neipp, L.; Prelog, V.; Zähner, H. Stoffwechselprodukte von Actinomyceten. 3. Mitteilung. Nonactin. Helv. Chim. Acta 1955, 38, 1445–1448. [Google Scholar] [CrossRef]
- Bérdy, J. Handbook of Antibiotic Compounds; CRC Press: Boca Raton, FL, USA, 1980; p. 327. ISBN 0849334500. [Google Scholar]
- Islam, M.T.; Laatsch, H.; von Tiedemann, A. Inhibitory effects of macrotetrolides from Streptomyces spp. on zoosporogenesis and motility of peronosporomycete zoospores are likely linked with enhanced ATPase activity in mitochondria. Front. Microbiol. 2016, 7, 1824. [Google Scholar] [CrossRef]
- Guzei, I.A.; Wang, C.; Zhan, Y.; Dolomanov, O.V.; Cheng, Y.Q. Pseudomerohedrally twinned monoclinic structure of unfolded ‘free’ nonactin: Comparative analysis of its large conformational change upon encapsulation of alkali metal ions. Acta Crystallogr. C Struct. Chem. 2009, 65 Pt 10, o521–o524. [Google Scholar] [CrossRef]
- Sheldrick, G.M. SHELXT–Integrated space-group and crystal-structure determination. Acta Crystallogr. A Found Adv. 2015, 71, 3–8. [Google Scholar] [CrossRef]
- Sheldrick, G.M. Crystal structure refinement with SHELXL. Acta Crystallogr. C Struct. Chem. 2015, 71, 3–8. [Google Scholar] [CrossRef]
- El-Tarabily, K.A. Promotion of tomato (Lycopersicon esculentum Mill.) plant growth by rhizosphere competent 1-aminocyclopropane-1-carboxylic acid deaminase–producing Streptomycete actinomycetes. Plant Soil 2008, 308, 161–174. [Google Scholar] [CrossRef]
- Glick, B.R.; Karaturovic, D.M.; Newell, P.C. A novel procedure for rapid isolation of plant growth promoting pseudomonas. Can. J. Microbiol. 1995, 41, 533–536. [Google Scholar] [CrossRef]
- Belimov, A.A.; Safronova, V.I.; Sergeyeva, T.A.; Egorova, T.N.; Matveyeva, V.A.; Tsyganov, V.E.; Borisov, A.Y.; Tikhonovich, I.A.; Kluge, C.; Preisfeld, A.; et al. Characterisation of plant growth-promoting rhizobacteria isolated from polluted soils and containing 1-aminocyclopropane-1-carboxylate deaminase. Can. J. Microbiol. 2001, 47, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Bignell, D.R.D.; Fyans, J.K.; Cheng, Z. Phytotoxins produced by plant pathogenic Streptomyces species. J. Appl. Microbiol. 2014, 116, 223–235. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | Strain Name | Diameter of the Growth Inhibition Zone, mm |
|---|---|---|
| Bacteria | ||
| Bacillus subtilis | VIZR-B10 | 13 ± 1 ab |
| Clavibacfer michiganense subsp. michiganense | VIZR-13a | 35 ± 3 d |
| Erwinia caratovora subsp. caratovora | VIZR: 160 | 22 ± 3 c |
| Escherichia coli | T-047 | 0 |
| Pseudomonas fluoresceus | VKM-B-894 | 0 |
| Staphylococcus aureus | 209p | 12 ± 1 ab |
| Streptomyces globisporus | VKM-Ac-179 | 10 ± 1 a |
| Xanthomonas campestris pv. vesicatoria | VIZR-322 | 0 |
| Fungi | ||
| Alternaria solani | VIZR-32 | 24 ± 3 c |
| Ascochyta melonis | VIZR-2-21 | 0 |
| Aspergillus niger | ATCC16404 | 0 |
| Fusarium graminearum | COP | 12 ± 1 ab |
| Fusarium redolens | VIZR-11 | 15 ± 1 b |
| Fusarium sambucinum | VIZR-18 | 12 ± 1 ab |
| Fusarium solani | VIZR-38 | 20 ± 3 c |
| Penicillium granulatum | ATCC-10450 | 0 |
| Rhizoctonia solani | VIZR-28 | 15 ± 2 b |
| Saccharomyces cerevisiae | K-12 | 0 |
| Sclerotinia libertiana | VIZR-32-1 | 25 ± 3 c |
| Trichoderma gypseum | VIZR-G-24 | 10 ± 2 a |
| Aphid Species | Lethal Concentrations, mg mL−1 | ||
|---|---|---|---|
| LC50 | LC90 | LC98 | |
| Medoura viciae | 1.1 | 4.2 | 6.0 |
| Aphis gossypii | 0.6 | 3.2 | 4.7 |
| Myzus persicae | 0.3 | 3.5 | 5.4 |
| Acyrthosiphon pisum | 1.8 | 6.9 | 10.0 |
| Neomyzus. circumtlexus | 2.6 | 5.0 | 6.5 |
| Fraction Collection Time | Starting Solvents Ratio, Vol % | Final Solvents Ratio, Vol % | Fraction Number | Fraction Volume, mL | Aphid Mortality, % |
|---|---|---|---|---|---|
| 0 min–2 min 55 s | A 100 | A 100 | 1 | 45 | 37 |
| 2 min 55 s–3 min | A 100 | A–B 90:10 | |||
| 3 min–5 min 55 s | A–B 90:10 | A–B 90:10 | 2 | 45 | 42 |
| 5 min 55 s–6 min | A–B 90:10 | A–B 80:20 | |||
| 6 min–8 min 55 s | A–B 80:20 | A–B 80:20 | 3 | 45 | 40 |
| 8 min 55 s–9 min | A–B 80:20 | A–B 70:30 | |||
| 9 min–11 min 55 s | A–B 70:30 | A–B 70:30 | 4 | 45 | 93 |
| 11 min 55 s–12 min | A–B 70:30 | A–B 60:40 | |||
| 12 min–14 min 55 s | A–B 60:40 | A–B 60:40 | 5 | 45 | 100 |
| 14 min 55 s–15 min | A–B 60:40 | A–B 50:50 | |||
| 15 min–17 min 55 s | A–B 50:50 | A–B 50:50 | 6 | 45 | 92 |
| 17 min 55 s–18 min | A–B 50:50 | A–B 40:60 | |||
| 18 min–20 min 55 s | A–B 40:60 | A–B 40:60 | 7 | 45 | 87 |
| 20 min 55 s–21 min | A–B 40:60 | A–B 30:70 | |||
| 21 min–23 min 55 s | A–B 30:70 | A–B 30:70 | 8 | 45 | 57 |
| 23 min 55 s–24 min | A–B 30:70 | A–B 20:80 | |||
| 24 min–26 min 55 s | A–B 20:80 | A–B 20:80 | 9 | 45 | 42 |
| 26 min 55 s–27 min | A–B 20:80 | A–B 10:90 | |||
| 27 min–29 min 55 s | A–B 10:90 | A–B 10:90 | 10 | 45 | 40 |
| 29 min 55 s–30 min | A–B 10:90 | B 100 | |||
| 30 min–32 min 55 s | B 100 | B 100 | 11 | 45 | 32 |
| 32 min 55 s–33 min | B 100 | B 100 | |||
| 33 min–36 min | C 100 | C 100 | 12 | 45 | 34 |
| 36 min–39 min | C 100 | C 100 | 13 | 45 | 0 |
| Formula | C40H64O12 |
| Molar mass | 736.91 |
| Diffractometer | Kappa Apex II (Bruker AXS) |
| Temperature (K) | 100.0 |
| Radiation source | CuKα (λ = 1.54178 Å) |
| Space group | P2/n |
| Cell parameters: a, b, c (E) | 28.5271(9), 5.6924(2), 28.5193(9) |
| б, в, г (°) | 90, 113.044(2), 90 |
| Volume (E3) | 4261.6(2) |
| Z | 4 |
| The range of angles 2I (°) | 3.366–105.908 |
| Resolution (E) | 26.2508–0.9658 |
| Index range | −29 ≤ h ≤ 29, −5 ≤ k ≤ 5, −29 ≤ l ≤ 28 |
| Total reflexes | 68422 |
| Independent reflexes | 4887 |
| Completeness (%) | 98.97 |
| Average I/y(I) | 52.64 |
| Rint | 0.0449 |
| Rsigma | 0.0190 |
| Data/parameters | 4887/478 |
| R [I > =2y (I)] | R1 = 0.1067, wR2 = 0.2448 |
| Treatments | Tetranychus urticae Mortality, % |
|---|---|
| Tap water | Nd |
| 1.25 g DEE L−1 | 56 ± 8 a |
| 2.5 g DEE L−1 | 89 ± 5 b |
| 5.0 g DEE L−1 | 98 ± 4 b |
| 10.0 g DEE L−1 | 100 ± 3 b |
| 20.0 g DEE L−1 | 95 ± 5 b |
| Fitoverm (2.0 g Aversectin-C L−1) | 95 ± 5 b |
| Treatments | Seed Germination, % | Root Length, mm | ||
|---|---|---|---|---|
| Barley | Watercress | Barley | Watercress | |
| Control | 58 ± 5 a | 94 ± 2 b | 108 ± 5 b | 33 ± 2 a |
| Supernatant | 73 ± 6 a | 85 ± 5 b | 89 ± 4 a | 31 ± 3 a |
| 2 g L−1 DEE | 45 ± 6 a | 85 ± 3 b | 105 ± 5 b | 37 ± 2 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boykova, I.; Yuzikhin, O.; Novikova, I.; Ulianich, P.; Eliseev, I.; Shaposhnikov, A.; Yakimov, A.; Belimov, A. Strain Streptomyces sp. P-56 Produces Nonactin and Possesses Insecticidal, Acaricidal, Antimicrobial and Plant Growth-Promoting Traits. Microorganisms 2023, 11, 764. https://doi.org/10.3390/microorganisms11030764
Boykova I, Yuzikhin O, Novikova I, Ulianich P, Eliseev I, Shaposhnikov A, Yakimov A, Belimov A. Strain Streptomyces sp. P-56 Produces Nonactin and Possesses Insecticidal, Acaricidal, Antimicrobial and Plant Growth-Promoting Traits. Microorganisms. 2023; 11(3):764. https://doi.org/10.3390/microorganisms11030764
Chicago/Turabian StyleBoykova, Irina, Oleg Yuzikhin, Irina Novikova, Pavel Ulianich, Igor Eliseev, Alexander Shaposhnikov, Alexander Yakimov, and Andrey Belimov. 2023. "Strain Streptomyces sp. P-56 Produces Nonactin and Possesses Insecticidal, Acaricidal, Antimicrobial and Plant Growth-Promoting Traits" Microorganisms 11, no. 3: 764. https://doi.org/10.3390/microorganisms11030764
APA StyleBoykova, I., Yuzikhin, O., Novikova, I., Ulianich, P., Eliseev, I., Shaposhnikov, A., Yakimov, A., & Belimov, A. (2023). Strain Streptomyces sp. P-56 Produces Nonactin and Possesses Insecticidal, Acaricidal, Antimicrobial and Plant Growth-Promoting Traits. Microorganisms, 11(3), 764. https://doi.org/10.3390/microorganisms11030764