
Comparative Genomic Analysis and Physiological Properties of Limosilactobacillus fermentum SMFM2017-NK2 with Ability to Inflammatory Bowel Disease


Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Lactic Acid Bacteria
2.2. Development of a Colitis Mouse Model and Administration of Lactic Acid Bacteria
2.3. Weight Measurement
2.4. Determination of Disease Activity Index (DAI)
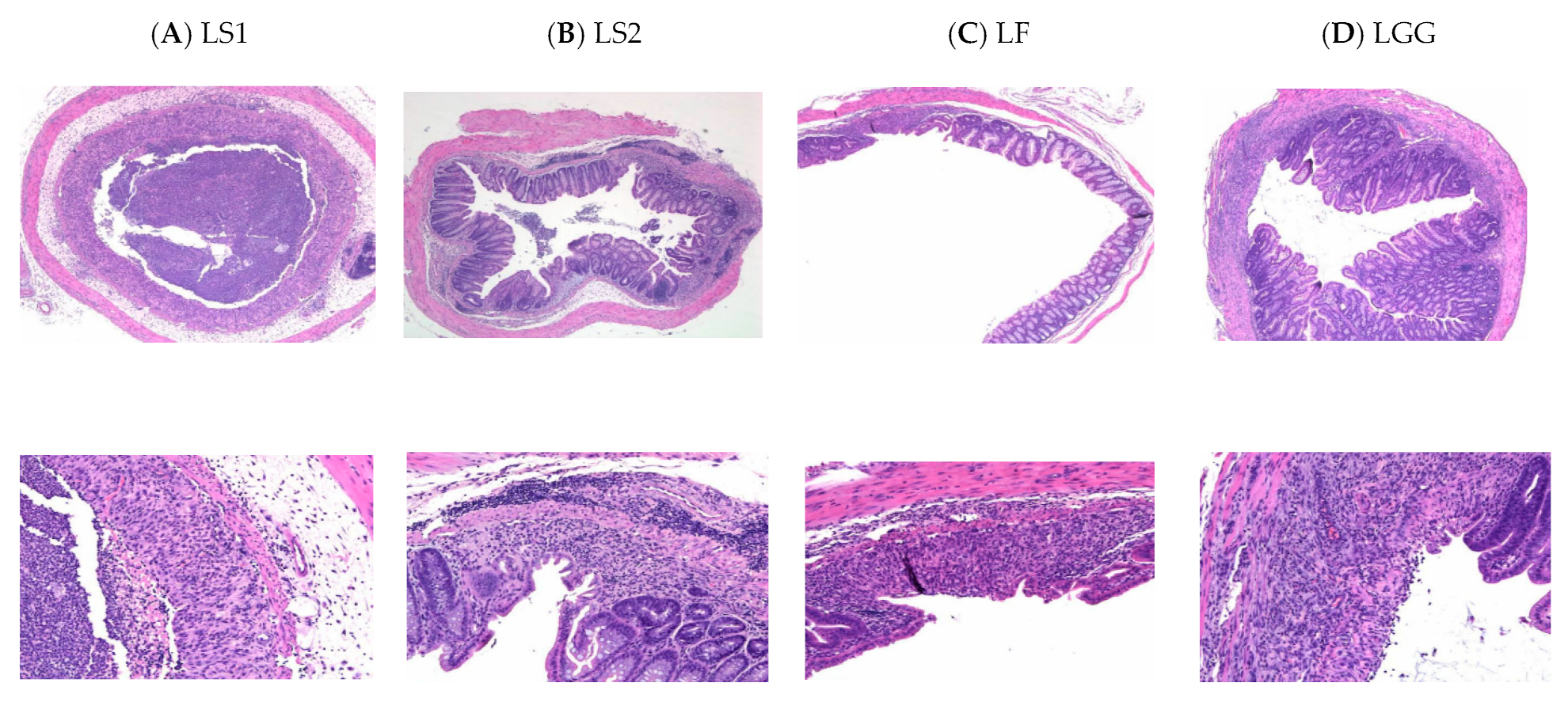
2.5. Histopathological Analysis of Colon Tissue
2.6. Analysis of Inflammation-Related Gene Expression in the Colon
2.7. Analysis of Biochemical Characteristics
2.8. Observation of Bacterial Morphology
2.9. Analysis of Cytotoxicity
2.10. Evaluation of Heat Stability
2.11. DNA Library Preparation and Sequencing
2.12. Gene Prediction and Taxonomic Profiling
2.13. Orthologous Genes Clustering
2.14. Analysis of Single Nucleotide Polymorphism (SNP) and Insertion/Deletion (InDel)
2.15. Statistical Analysis
3. Results and Discussion
3.1. Body Weight
3.2. Length of Large and Small Intestine
3.3. DAI Score during Administration of Lactic Acid Bacteria
3.4. Histopathological Features of Colon
3.5. Gene Expression of Proinflammatory Cytokines in Colon
3.6. Biochemical Characteristics
3.7. Morphological Characteristics
3.8. Cytotoxicity
3.9. Heat Stability
3.10. General Features of L. fermentum SMFM2017-NK2 Genome
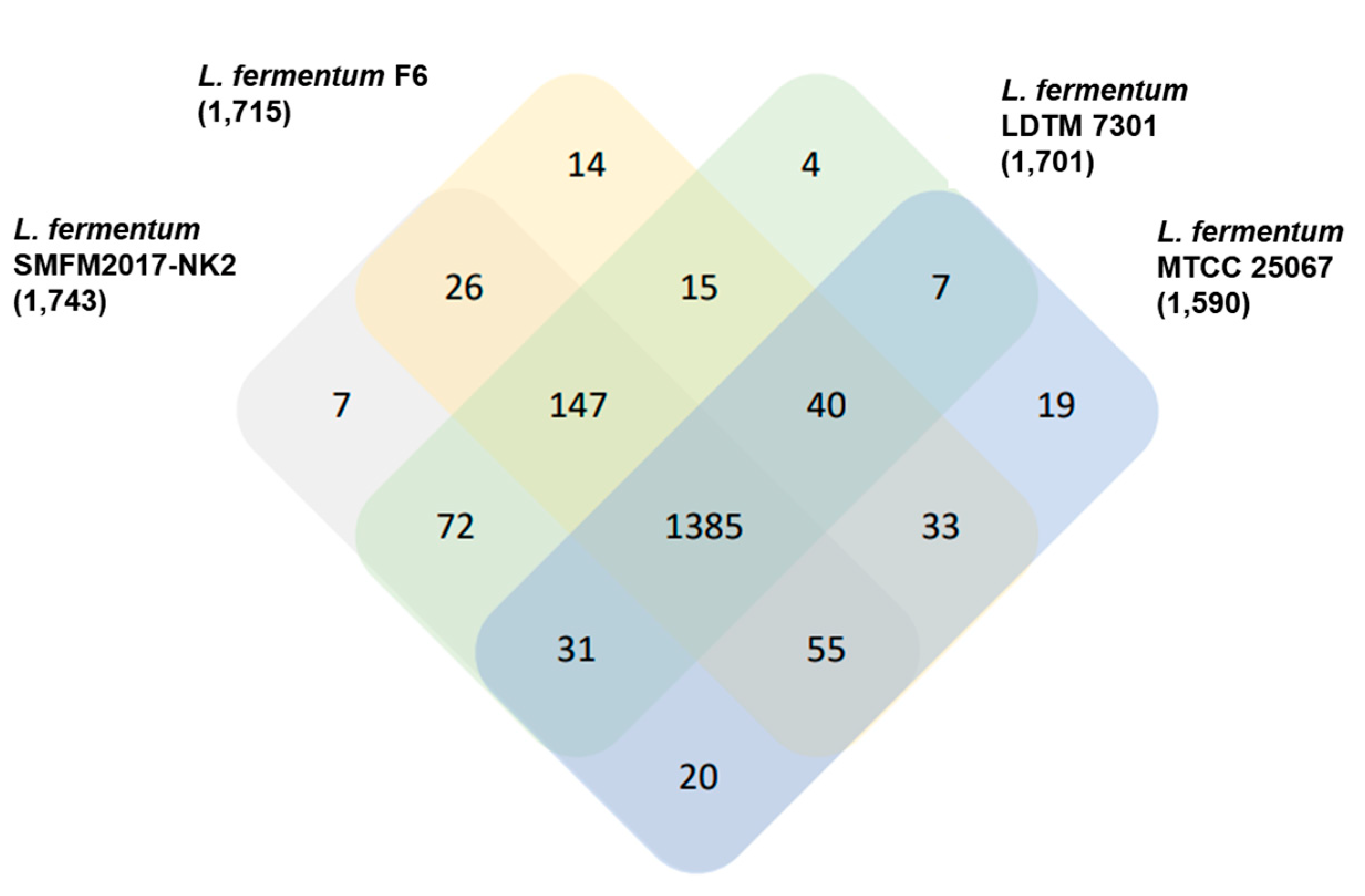
3.11. Orthologous Gene Cluster of L. fermentum SMFM2017-NK2
3.12. Genomic Variation of L. fermentum SMFM2017-NK2
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kim, M.J.; Chung, H.J. Quality Characteristics and Antioxidant Activities of Yanggaeng added with Pleurotus eryngii Powder. J. East. Asian Soc. Diet. Life 2017, 27, 69–77. [Google Scholar] [CrossRef]
- Lee, K.H.; Rhee, K.H. Anti-Inflammatory Effects of Glycyrrhiza glabra Linne Extract in a Dextran Sulfate Sodium-Induced Colitis Mouse Model. Korean J. Food Nutr. 2010, 23, 435–439. [Google Scholar]
- Ha, J.E.; Jang, E.J.; Im, S.G.; Sohn, H.S. Medication use and drug expenditure in inflammatory bowel disease: Based on Korean National Health Insurance Claims Data (2010–2014). Korean J. Clin. Pharm. 2019, 29, 79–88. [Google Scholar] [CrossRef]
- Chang, B.Y.; Han, J.H.; Cha, B.S.; Ann, S.H.; Kim, S.Y. Optimization of Culture Condition for Enhancing the Probiotics Functions. J. Food Hyg. Saf. 2015, 30, 295–301. [Google Scholar] [CrossRef]
- Kim, J.M. Roles of enteric microbial composition and metabolism in health and diseases. Korean J. Gastroenterol. 2013, 62, 191–205. [Google Scholar] [CrossRef]
- Park, J.S.; Joe, I.; Rhee, P.D.; Jeong, C.S.; Jeong, G. A lactic acid bacterium isolated from kimchi ameliorates intestinal inflammation in DSS-induced colitis. J. Microbiol. 2017, 55, 304–310. [Google Scholar] [CrossRef]
- Mardini, H.E.; Grigorian, A.Y. Probiotic mix VSL#3 is effective adjunctive therapy for mild to moderately active ulcerative colitis: A meta-analysis. Inflamm. Bowel Dis. 2014, 20, 1562–1567. [Google Scholar]
- Jung, M.Y.; Park, Y.H.; Kim, H.S.; Poo, H.R.; Chang, Y.H. Probiotic property of Lactobacillus pentosus Miny-148 isolated from human feces. Korean J. Microbiol. 2009, 45, 177–184. [Google Scholar]
- Jang, H.J. Potential Use of Lactic acid Bacteria Isolated from Kimchi as Probiotics. Master’s Thesis, Sookmyung Women’s University Graduate School, Seoul, Republic of Korea, 2018. [Google Scholar]
- Erben, U.; Loddenkemper, C.; Doerfel, K.; Spieckermann, S.; Haller, D.; Heimesaat, M.M.; Zeitz, M.; Seigmund, B.; Kühl, A.A. A guide to histomorphological evaluation of intestinal inflammation in mouse models. Int. J. Clin. Exp. Pathol. 2014, 7, 4557. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Kim, K.A.; Gu, W.; Lee, I.A.; Joh, E.H.; Kim, D.H. High fat diet-induced gut microbiota exacerbates inflammation and obesity in mice via the TLR4 signaling pathway. PLoS ONE 2012, 7, e47713. [Google Scholar] [CrossRef]
- Ranatunga, D.C.; Ramakrishnan, A.; Uprety, P.; Wang, F.; Zhang, H.; Margolick, J.B.; Brayton, C.; Bream, J.H. IL-10–expressing CD4+ T Cells in colitis. J. Immunol. 2012, 189, 1243–1252. [Google Scholar] [CrossRef]
- Song, M.; Baik, H.W.; Hong, S.G.; Sung, M.K. Wheat bran arabinoxylan supplementation alleviates 5-fluorouracil induced mucositis and myelosuppression in BALB/c mice. J. Funct. Foods 2016, 21, 312–320. [Google Scholar] [CrossRef]
- Weisberg, S.P.; McCann, D.; Desa, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef]
- Murray, L.J.; Lee, R.; Martens, C. In vivo cytokine gene 82 expression in T cell subsets of the autoimmune MRL/Mp-lpr/lpr mouse. Eur. J. Immunol. 1990, 20, 163–170. [Google Scholar] [CrossRef]
- Shin, Y.W.; Bae, E.A.; Kim, S.S.; Kim, D.H. Effect of ginsenoside Rb1 and compound K in chronic oxazolone-induced mouse dermatitis. Int. Immunopharmacol. 2005, 5, 1183–1191. [Google Scholar] [CrossRef]
- Mardis, E.R. Next-generation DNA sequencing methods. Annu. Rev. Genom. Hum. Genet. 2008, 9, 387–402. [Google Scholar] [CrossRef]
- Peng, Y.; Leung, H.C.; Yiu, S.M.; Chin, F.Y. IDBA-UD: A de novo assembler for single-cell and metagenomic sequencing data with highly uneven depth. J. Bioinform. 2012, 28, 1420–1428. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. J. Bioinform. 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Hyatt, D.; Chen, G.L.; LoCascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef] [PubMed]
- Laslett, D.; Canback, B. ARAGORN, a program to detect tRNA genes and tmRNA genes in nucleotide sequences. Nucleic Acids Res. 2004, 32, 11–16. [Google Scholar] [CrossRef]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef] [PubMed]
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive metagenomic visualization in a Web browser. BMC Bioinform. 2011, 12, 385. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Stoeckert, C.J.; Roos, D.S. OrthoMCL: Identification of ortholog groups for eukaryotic genomes. Genome Res. 2003, 13, 2178–2189. [Google Scholar] [CrossRef] [PubMed]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef]
- Jung, H.C. Animal model for inflammatory bowel disease. Korean J. Gastroenterol. 2001, 37, 69–75. [Google Scholar]
- Ahn, Y.T.; Lee, H.; Lee, J.H.; Kim, D.H.; Huh, C.S. Anti-colitic Effect of Lactic Acid Bacteria R&B Balance®. Curr. Top. Lact. Acid. Bact. Probiotics. 2013, 1, 70–78. [Google Scholar]
- Lee, H.Y.; Ahn, Y.T.; Park, S.H.; Ahn, Y.M.; Shim, J.J.; Lee, J.; Lee, J.S.; Surh, Y.J.; Huh, C.S.; Kim, D.H. Evaluation of Anti-Colitic Effect of Chung-Jang-Hwan (C-mix) in Mice. Biomol Ther. 2011, 19, 52–58. [Google Scholar] [CrossRef]
- Myung, N.Y. The improving effect of Zizyphus Jujube on dextran sulfate sodium-induced colitis in mice. Korea J. Herbology. 2015, 30, 35–40. [Google Scholar] [CrossRef]
- Fan, H.; Qiu, M.Y.; Mei, J.J.; Shen, G.X.; Liu, S.L.; Chen, R. Effects of four regulating-intestine prescriptions on pathology and ultrastructure of colon tissue in rats with ulcerative colitis. World J. Gastroenterol. 2005, 11, 4800. [Google Scholar] [CrossRef]
- Chassaing, B.; Aitken, J.D.; Malleshappa, M.; Vijay-Kumar, M. Dextran sulfate sodium (DSS)-induced colitis in mice. Curr. Protoc. Immunol. 2014, 104, 15.25.1–15.25.14. [Google Scholar] [CrossRef]
- Chen, Z.; Yi, L.; Pan, Y.; Long, X.; Mu, J.; Yi, R.; Zhao, X. Lactobacillus fermentum ZS40 ameliorates inflammation in mice with ulcerative colitis induced by dextran sulfate sodium. Front. Pharmacol. 2021, 12, 700217. [Google Scholar] [CrossRef]
- Zhou, X.; Liu, H.; Zhang, J.; Mu, J.; Zalan, Z.; Hegyi, F. Protective effect of Lactobacillus fermentum CQPC04 on dextran sulfate sodium-induced colitis in mice is associated with modulation of the nuclear factor-kappaB signaling pathway. J. Dairy Sci. 2019, 102, 9570–9585. [Google Scholar] [CrossRef]
- Fang, L.; Hui, H.; Fulejia, Z.; Xianrong, Z.; Zhennai, Y.; Xin, Z. A Mixture of Lactobacillus fermentum HFY06 and arabinoxylan ameliorates dextran sulfate dodium-induced acute ulcerative colitis in mice. J. Inflamm. 2021, 14, 6575–6585. [Google Scholar]
- Wang, Y.; Wu, J.; Lv, M.; Shao, Z.; Hungwe, M.; Wang, J.; Bai, X.; Xie, J.; Wang, Y.; Geng, W. Metabolism characteristics of lactic acid bacteria and the expanding applications in food industry. Front. Bioeng. Biotechnol. 2021, 9, 612285. [Google Scholar] [CrossRef] [PubMed]
- Hayek, S.A.; Ibrahim, S.A. Current limitations and challenges with lactic acid bacteria: A review. Food Sci. Nutr. 2013, 4, 73–87. [Google Scholar] [CrossRef]
- Hertel, C.; Schmidt, G.; Fischer, M.; Oellers, K.; Hammes, W.P. Oxygen-dependent regulation of the expression of the catalase gene katA of Lactobacillus sakei LTH677. Appl. Environ. Microbiol. 1998, 64, 1359–1365. [Google Scholar] [CrossRef]
- Cho, M.K.; Kim, K.; Kim, C.H.; Lee, T.K.; Kim, K.Y. Isolation and characterization of Lactobacillus fermentum YL-3 as a poultry probiotic. Kor. J. Appl. Microbiol. Biotechnol. 2000, 28, 279–284. [Google Scholar]
- Shields, P.; Cathcart, L. Oxidase Test Protocol. Am. Soc. Microbiol. 2010, 1–9. [Google Scholar]
- Yoon, J.A.; Shin, K.O. Studies on the function of lactic acid bacteria and related yeasts in probiotics: A review. Kor. J. Food Nutr. 2015, 30, 395–404. [Google Scholar]
- Seo, J.H.; Lee, H. Characteristic and immunomodulating activity of lactic acid bacteria for the potential probiotics. Korean J. Food Sci. Technol. 2007, 39, 681–687. [Google Scholar]
- Xu, M.L.; Hu, J.H.; Wang, L.; Kim, H.S.; Jin, C.W.; Cho, D.H. Antioxidant and Anti-diabetes Activity of Extracts from Machilus thunbergii S. et Z. Korean J. Med. Crop. Sci. 2010, 18, 34−39. [Google Scholar]
- Oh, Y.J.; Lee, M.S.; Ahn, Y.J.; Ahn, J.H. Application of Measurement Method for Lactic Acid Bacteria Content on Coccus Including Yoghurt. Curr. Top. Lact. Acid Bac. Probio. 2015, 3, 34–36. [Google Scholar] [CrossRef]
- Gabaldón, T.; Koonin, E.V. Functional and evolutionary implications of gene orthology. Nat. Rev. Genet. 2013, 14, 360–366. [Google Scholar] [CrossRef]
- Ijaq, J.; Malik, G.; Kumar, A.; Das, P.S.; Meena, N.; Bethi, N.; Sundararajan, V.S.; Suravajhala, P. A model to predict the function of hypothetical proteins through a nine-point classification scoring schema. BMC Bioinform. 2019, 20, 1–8. [Google Scholar] [CrossRef]
- Kosinski, J.; Feder, M.; Bujnicki, J.M. The PD-(D/E) XK superfamily revisited: Identification of new members among proteins involved in DNA metabolism and functional predictions for domains of (hitherto) unknown function. BMC Bioinform. 2005, 6, 172. [Google Scholar] [CrossRef]
- Davalos-Garcia, M.; Conter, A.; Toesca, I.; Gutierrez, C.; Cam, K. Regulation of osmC Gene Expression by the Two-Component System rcsB-rcsC in Escherichia coli. J. Bacteriol. 2001, 183, 5870–5876. [Google Scholar] [CrossRef]
- SnpEff. Available online: https://pcingola.github.io/SnpEff/se_introduction/ (accessed on 29 December 2020).
- FAO/WHO. Joint FAO/WHO Working Group Report on Drafting Guidelines for the Evaluation of Probiotics in Food; FAO: Rome, Italy, 2002. [Google Scholar]
- Vitetta, L.; Coulson, S.; Thomsen, M.; Nguyen, T.; Hall, S. Probiotics, D-Lactic acidosis, oxidative stress and strain specificity. Gut Microbes. 2017, 8, 311–322. [Google Scholar] [CrossRef]
- Begley, M.; Hill, C.; Gahan, C.G. Bile salt hydrolase activity in probiotics. Appl. Environ. Microbiol. 2006, 72, 1729–1738. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′->3′) | References | |
|---|---|---|---|
| β-actin | Forward | CCGTGAAAAGATGACCCAGATC | [12] |
| Reverse | CACAGCCTGGATGGCTACGT | ||
| TNF-α | Forward | TCTTCTCATTCCTGCTTGTGG | [13] |
| Reverse | GGTCTGGGGCATAGAAGGA | ||
| IL-1β | Forward | AACCTGCTGGTGTGTGACGTTC | [12] |
| Reverse | CAGCACGAGGCTTTTTTGTTGT | ||
| IL-6 | Forward | ACCAGAGGAAATTTTGAATAGGC | [14] |
| Reverse | TGATGCACTTGCAGAAAACA | ||
| INOS | Forward | AATCTTGGAGCGAGTTGTGG | [15] |
| Reverse | CAGGAAGTAGGGAGGGCTTG | ||
| IFN-γ | Forward | AGCGGCTGACTGAACTCAGATTGTAG | [16] |
| Reverse | GTCACAGTTTTCAGCTGTATAGGG | ||
| COX-2 | Forward | TGTATCCCCCCACAGTCAAAGACAC | [17] |
| Reverse | GTGCTCCCGAAGCCAGATGG | ||
| Histopathological Features | Group | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LS1 | LS2 | LF | LGG | |||||||||||||
| Infiltration of inflammatory cells | 3 | 3 | 3 | 0 | 3 | 2 | 1 | 0 | 1 | 3 | 0 | 0 | 0 | 1 | 3 | 3 |
| Depth of inflammatory cell infiltration | 2 | 2 | 2 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 1 | 3 |
| Crypt | 2 | 2 | 3 | 0 | 1 | 2 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 2 |
| Edema | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Mucoprotein | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Hemorrhage | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Pseudomembrane | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sum | 23 | 13 | 8 | 16 | ||||||||||||
| Orthologous Group | Gene | Start Position | End Position | Sequence Length (bp) | Type | Function |
|---|---|---|---|---|---|---|
| H001716 | GENE_01478 | 382 | 591 | 210 | CDS | hypothetical protein |
| GENE_02186 | 160,903 | 161,112 | 210 | CDS | hypothetical protein | |
| H001717 | GENE_01485 | 3011 | 3388 | 378 | CDS | hypothetical protein |
| GENE_02175 | 156,223 | 156,648 | 426 | CDS | hypothetical protein | |
| H001737 | GENE_02180 | 159,325 | 159,477 | 153 | CDS | hypothetical protein |
| GENE_04647 | 106,749 | 106,883 | 135 | CDS | hypothetical protein | |
| H001777 | GENE_03909 | 32 | 433 | 402 | CDS | hypothetical protein |
| GENE_04681 | 120,449 | 120,838 | 390 | CDS | hypothetical protein | |
| H001778 | GENE_03910 | 426 | 1274 | 849 | CDS | hypothetical protein |
| GENE_04680 | 119,608 | 120,456 | 849 | CDS | PD-(D/E)XK nuclease superfamily protein | |
| H001789 | GENE_04624 | 78,078 | 78,485 | 408 | CDS | hypothetical protein |
| GENE_05049 | 900 | 1265 | 366 | CDS | hypothetical protein | |
| H001794 | GENE_05028 | 1293 | 1730 | 438 | CDS | OsmC-like protein |
| GENE_05029 | 1742 | 2164 | 423 | CDS | OsmC-like protein |
| Region of the Genome | Count | Percent (%) |
|---|---|---|
| Upstream | 15,522 | 44.26 |
| Downstream | 16,302 | 46.48 |
| Exon | 2648 | 7.55 |
| Intergenic | 596 | 1.70 |
| Transcript | 1 | 0.00 |
| None | 1 | 0.00 |
| Effect | Position | Reference Allel | Alternative Allele | Depth | Codon | Amino Acid | Gene | Transcript |
|---|---|---|---|---|---|---|---|---|
| Frameshift_variant | 16,086 | C | CG | 369 | c.635dupG | p.Val213fs | Gene_gene17 | gene17 |
| 135,033 | C | CG | 391 | c.96dupG | p.Pro33fs | Gene_gene151 | gene151 | |
| 157,379 | T | TG | 319 | c.516dupG | p.Arg173fs | Gene_gene175 | gene175 | |
| 327,600 | A | AC | 216 | c.538dupC | p.Arg180fs | BGV76_RS01760 | Transcript_gene350 | |
| 401,859 | C | CA | 716 | c.524dupT | p.Ile176fs | BGV76_RS02115 | Transcript_gene421 | |
| 402,254 | A | ATCGC | 580 | c.129_130insGCGA | p.Tyr44fs | BGV76_RS02115 | Transcript_gene421 | |
| 402,280 | GA | G | 638 | c.103delT | p.Ser35fs | BGV76_RS02115 | Transcript_gene421 | |
| 531,259 | CT | C | 308 | c.209delA | p.Glu70fs | Gene_gene559 | gene559 | |
| 579,296 | G | GC | 288 | c.126dupC | p.Ser43fs | Gene_gene613 | gene613 | |
| 606,522 | C | CG | 321 | c.968dupG | p.Cys324fs | Gene_gene637 | gene637 | |
| 619,737 | A | AAT | 59 | c.4_5insAT | p.Ser2fs | Gene_gene646 | gene646 | |
| 664,789 | T | TCCTAAATTGCAAGATTAAGTGAGCCACCCGGCCACGGGAG | 93 | c.410_411insCTCCCGTGGCCGGGTGGCTCACTTAATCTTGCAATTTAGG | p.Gln137fs | tnpA | Transcript_gene696 | |
| 727,563 | T | TC | 274 | c.557dupG | p.Val187fs | Gene_gene763 | gene763 | |
| 881,163 | G | GC | 311 | c.698dupG | p.Lys234fs | Gene_gene905 | gene905 | |
| 1,039,055 | AG | A | 427 | c.1167delC | p.Ser389fs | BGV76_RS05400 | Transcript_gene1078 | |
| 1,076,406 | CG | C | 408 | c.383delG | p.Gly128fs | BGV76_RS05580 | Transcript_gene1114 | |
| 1,076,406 | CG | C | 408 | c.19delG | p.Ala7fs | BGV76_RS05585 | Transcript_gene1115 | |
| 1,094,586 | CG | C | 259 | c.75delC | p.Cys25fs | BGV76_RS05690 | Transcript_gene1136 | |
| 1,094,590 | CA | C | 264 | c.71delT | p.Val24fs | BGV76_RS05690 | Transcript_gene1136 | |
| 1,212,247 | AACAAAGAAAT | A | 63 | c.242_251delACAAAGAAAT | p.Asn81fs | Gene_gene1265 | gene1265 | |
| 1,212,259 | CTATT | C | 50 | c.254_257delTATT | p.Leu85fs | Gene_gene1265 | gene1265 | |
| 1,224,178 | C | CT | 423 | c.1164dupA | p.Ala389fs | Gene_gene1275 | gene1275 | |
| 1,245,304 | TA | T | 387 | c.204delT | p.Phe68fs | BGV76_RS06470 | Transcript_gene1292 | |
| 1,252,008 | A | AG | 351 | c.377dupG | p.Val127fs | BGV76_RS06515 | Transcript_gene1301 | |
| 1,408,360 | CG | C | 400 | c.1306delC | p.Arg436fs | Gene_gene1448 | gene1448 | |
| 1,472,109 | CT | C | 425 | c.55delA | p.Ser19fs | Gene_gene1508 | gene1508 | |
| 1,494,271 | AT | A | 581 | c.355delA | p.Ile119fs | Gene_gene1538 | gene1538 | |
| 1,496,486 | T | TTA | 673 | c.220_221insAT | p.Ser74fs | Gene_gene1541 | gene1541 | |
| 1,567,158 | C | CG | 445 | c.968dupG | p.Arg324fs | Gene_gene1609 | gene1609 | |
| 1,570,436 | C | CG | 448 | c.230dupG | p.Ser78fs | Gene_gene1612 | gene1612 | |
| 1,578,725 | C | CG | 438 | c.749dupG | p.His251fs | Gene_gene1621 | gene1621 | |
| 1,589,275 | C | CG | 457 | c.108dupC | p.Asp37fs | BGV76_RS08160 | Transcript_gene1630 | |
| 1,598,314 | A | AG | 428 | c.1433dupG | p.Gly479fs | Gene_gene1636 | gene1636 | |
| 1,617,562 | CA | C | 530 | c.327delA | p.Lys109fs | BGV76_RS08260 | Transcript_gene1650 | |
| 1,662,103 | G | GAA | 41 | c.37_38insAA | p.Gly13fs | BGV76_RS08455 | Transcript_gene1689 | |
| 1,662,104 | GGC | G | 49 | c.39_40delGC | p.Gly13fs | BGV76_RS08455 | Transcript_gene1689 | |
| 1,664,328 | ATATTTCC | A | 62 | c.537_543delTATTTCC | p.Asp179fs | BGV76_RS08465 | Transcript_gene1691 | |
| 1,664,342 | G | GCC | 75 | c.550_551insCC | p.Glu184fs | BGV76_RS08465 | Transcript_gene1691 | |
| 1,664,344 | A | AACGAG | 84 | c.553_554insCGAGA | p.Lys185fs | BGV76_RS08465 | Transcript_gene1691 | |
| 1,673,647 | G | GC | 435 | c.284dupG | p.Cys95fs | Gene_gene1701 | gene1701 | |
| 1,704,505 | AC | A | 464 | c.29delG | p.Arg10fs | Gene_gene1732 | gene1732 | |
| 1,729,123 | CT | C | 455 | c.559delA | p.Arg187fs | BGV76_RS08790 | Transcript_gene1756 | |
| 1,729,237 | AATTGAAATGACAATTCCGGCCAGTAATCCTTTTTAACGAAAGCGTACTGGTAGAGGATGTCTAAGCCGTTATGGGTGATGGCAAG | A | 497 | c.361_445delCTTGCCATCACCCATAACGGCTTAGACATCCTCTACCAGTACGCTTTCGTTAAAAAGGATTACTGGCCGGAATTGTCATTTCAAT | p.Leu121fs | BGV76_RS08790 | Transcript_gene1756 | |
| 1,730,794 | A | AC | 413 | c.104dupC | p.Ter36fs | Gene_gene1758 | gene1758 | |
| 1,731,126 | CT | C | 415 | c.188delA | p.Lys63fs | BGV76_RS08805 | Transcript_gene1759 | |
| 1,925,766 | T | TC | 385 | c.1575dupG | p.Ile526fs | Gene_gene1960 | gene1960 | |
| 1,957,489 | G | GGC | 467 | c.195_196dupGC | p.His66fs | BGV76_RS09960 | Transcript_gene1990 | |
| 1,957,495 | CCT | C | 466 | c.203_204delCT | p.Ser68fs | BGV76_RS09960 | Transcript_gene1990 | |
| 1,977,748 | T | TG | 414 | c.1034dupG | p.Phe346fs | Gene_gene2007 | gene2007 | |
| Stop_gained | 348,140 | C | T | 339 | c.73C>T | p.Gln25* | BGV76_RS01850 | Transcript_gene368 |
| 1,076,573 | C | T | 358 | c.184C>T | p.Gln62* | BGV76_RS05585 | Transcript_gene1115 | |
| 1,094,442 | G | A | 401 | c.220C>T | p.Arg74* | BGV76_RS05690 | Transcript_gene1136 | |
| 1,094,621 | C | T | 270 | c.41G>A | p.Trp14* | BGV76_RS05690 | Transcript_gene1136 | |
| 1,094,634 | C | A | 251 | c.28G>T | p.Glu10* | BGV76_RS05690 | Transcript_gene1136 | |
| 1,175,482 | G | A | 409 | c.976C>T | p.Gln326* | BGV76_RS06175 | Transcript_gene1233 | |
| 1,191,164 | C | T | 1001 | c.70C>T | p.Gln24* | BGV76_RS06240 | Transcript_gene1246 | |
| 1,192,075 | C | T | 892 | c.502C>T | p.Arg168* | BGV76_RS06245 | Transcript_gene1247 | |
| 1,293,932 | C | T | 1589 | c.847C>T | p.Gln283* | BGV76_RS06730 | Transcript_gene1344 | |
| 1,401,457 | C | T | 439 | c.52C>T | p.Gln18* | BGV76_RS07210 | Transcript_gene1440 | |
| 1,412,900 | G | A | 320 | c.847C>T | p.Gln283* | BGV76_RS07270 | Transcript_gene1452 | |
| 1,469,046 | C | T | 488 | c.202C>T | p.Gln68* | BGV76_RS07540 | Transcript_gene1506 | |
| 1,472,072 | A | C | 433 | c.93T>G | p.Tyr31* | Gene_gene1508 | gene1508 | |
| 1,599,733 | C | A | 434 | c.2846C>A | p.Ser949* | Gene_gene1636 | gene1636 | |
| 1,661,887 | T | G | 152 | c.588T>G | p.Tyr196* | BGV76_RS08450 | Transcript_gene1688 | |
| 1,663,750 | A | T | 254 | c.883A>T | p.Arg295* | BGV76_RS08460 | Transcript_gene1690 | |
| 1,673,835 | G | A | 439 | c.97C>T | p.Gln33* | Gene_gene1701 | gene1701 | |
| 1,705,464 | G | T | 411 | c.4G>T | p.Glu2* | Gene_gene1734 | gene1734 | |
| 1,729,568 | G | A | 447 | c.115C>T | p.Arg39* | BGV76_RS08790 | Transcript_gene1756 | |
| Frameshift_variant & stop_gained | 328,669 | G | GGCTCTACACTAAATCTTGTTGATGGATTACCATCGTGGTGATTCGTTGACAGGAT | 294 | c.596_597insATCCTGTCAACGAATCACCACGATGGTAATCCATCAACAAGATTTAGTGTAGAGC | p.His199fs | BGV76_RS01765 | Transcript_gene351 |
| 619,738 | G | GATACTAAGCTT | 50 | c.5_6insATACTAAGCTT | p.Ser2fs | Gene_gene646 | gene646 | |
| 664,789 | T | TCCTAAATTGCAAGATTAAGTGAGCCACCCGGCCACGGGAG | 93 | c.208_209insCCTAAATTGCAAGATTAAGTGAGCCACCCGGCCACGGGAG | p.Leu70fs | BGV76_RS03485 | Transcript_gene695 | |
| 1,021,085 | A | AATCTACTGCTTAATCTTAGAACGTAAACTTC | 57 | c.1643_1644insGAAGTTTACGTTCTAAGATTAAGCAGTAGAT | p.Ter549fs | BGV76_RS05335 | Transcript_gene1065 | |
| 1,175,869 | C | CTGAAGTGAACCCCCGAGATTGGACAACAATCTCGGGGGTTTTATTATGA | 207 | c.588_589insTCATAATAAAACCCCCGAGATTGTTGTCCAATCTCGGGGGTTCACTTCA | p.Ala197fs | BGV76_RS06175 | Transcript_gene1233 | |
| 1,311,946 | T | TGGTTATGTCCGTATAATTGGTGTAAATTCTAAATAGGACTTTGTGAA | 419 | c.99_100insTTCACAAAGTCCTATTTAGAATTTACACCAATTATACGGACATAACC | p.Lys34fs | BGV76_RS06805 | Transcript_gene1359 | |
| 1,412,781 | G | GTGGTCAACTGAGCGTATTCTGCTGGGTATTCTTGCCCGTACTTTTC | 127 | c.965_966insGAAAAGTACGGGCAAGAATACCCAGCAGAATACGCTCAGTTGACCA | p.Phe322fs | BGV76_RS07270 | Transcript_gene1452 | |
| 1,704,983 | T | TCAAAGGGCAAGAAGTCCTGTTGCTGGTAGGTGGCCAGGGGCTG | 199 | c.346_347insCAGCCCCTGGCCACCTACCAGCAACAGGACTTCTTGCCCTTTG | p.Gln116fs | BGV76_RS08675 | Transcript_gene1733 | |
| Stop_lost& splice_region_variant | 2,72,047 | A | G | 16 | c.470A>G | p.Ter157Trpext*? | BGV76_RS01460 | Transcript_gene290 |
| 1,192,261 | T | C | 935 | c.688T>C | p.Ter230Glnext*? | BGV76_RS06245 | Transcript_gene1247 | |
| 1,705,777 | A | T | 398 | c.317A>T | p.Ter106Leuext*? | Gene_gene1734 | gene1734 | |
| 1,948,678 | A | C | 385 | c.399A>C | p.Ter133Tyrext*? | BGV76_RS09915 | Transcript_gene1981 | |
| Frameshift_variant & stop_lost | 1,476,796 | CATAA | C | 399 | c.192_195delATAA | p.Lys64fs | Gene_gene1517 | gene1517 |
| 1,832,671 | ATTTGCGGCAGCTTGTTCAAACTGGCGGTTCAAGGTATTCTGTGCTTCATAAGTTTCCTACTCCTGCACACGTTTTCTC | A | 959 | c.-1_74delGAGAAAACGTGTGCAGGAGTAGGAAACTTATGAAGCACAGAATACCTTGAACCGCCAGTTTGAACAAGCTGCCGCAAA | p.Lys1fs | Gene_gene1865 | gene1865 | |
| Start_lost& inframe_deletion | 669,110 | CCTTTAAGCGCATCAGA | C | 396 | c.-1_12delTCTGATGCGCTTAAAG | p.Met1_Lys4del | BGV76_RS03520 | Transcript_gene702 |
| Stop_gained & disruptive_inframe_insertion | 1,286,056 | C | CGGTTATGTCCGTATAATTGGTGTAAATTCTAAATAGGACTTTGTGAAA | 269 | c.125_126insGGTTATGTCCGTATAATTGGTGTAAATTCTAAATAGGACTTTGTGAAA | p.Thr27_Ala42dup | BGV76_RS06700 | Transcript_gene1338 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ann, S.; Choi, Y.; Yoon, Y. Comparative Genomic Analysis and Physiological Properties of Limosilactobacillus fermentum SMFM2017-NK2 with Ability to Inflammatory Bowel Disease. Microorganisms 2023, 11, 547. https://doi.org/10.3390/microorganisms11030547
Ann S, Choi Y, Yoon Y. Comparative Genomic Analysis and Physiological Properties of Limosilactobacillus fermentum SMFM2017-NK2 with Ability to Inflammatory Bowel Disease. Microorganisms. 2023; 11(3):547. https://doi.org/10.3390/microorganisms11030547
Chicago/Turabian StyleAnn, Sumin, Yukyung Choi, and Yohan Yoon. 2023. "Comparative Genomic Analysis and Physiological Properties of Limosilactobacillus fermentum SMFM2017-NK2 with Ability to Inflammatory Bowel Disease" Microorganisms 11, no. 3: 547. https://doi.org/10.3390/microorganisms11030547
APA StyleAnn, S., Choi, Y., & Yoon, Y. (2023). Comparative Genomic Analysis and Physiological Properties of Limosilactobacillus fermentum SMFM2017-NK2 with Ability to Inflammatory Bowel Disease. Microorganisms, 11(3), 547. https://doi.org/10.3390/microorganisms11030547