
Temporal Dynamics of Anaplasma marginale Infections and the Composition of Anaplasma spp. in Calves in the Mnisi Communal Area, Mpumalanga, South Africa

 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Consideration
2.2. Study Area
2.3. Animals
2.4. Study Design and Sample Collection
2.5. Genomic DNA Extraction and Quantitative Real-Time PCR (qPCR) Assay
2.6. Amplification, Cloning and Sequencing of the A. marginale Msp1α Gene
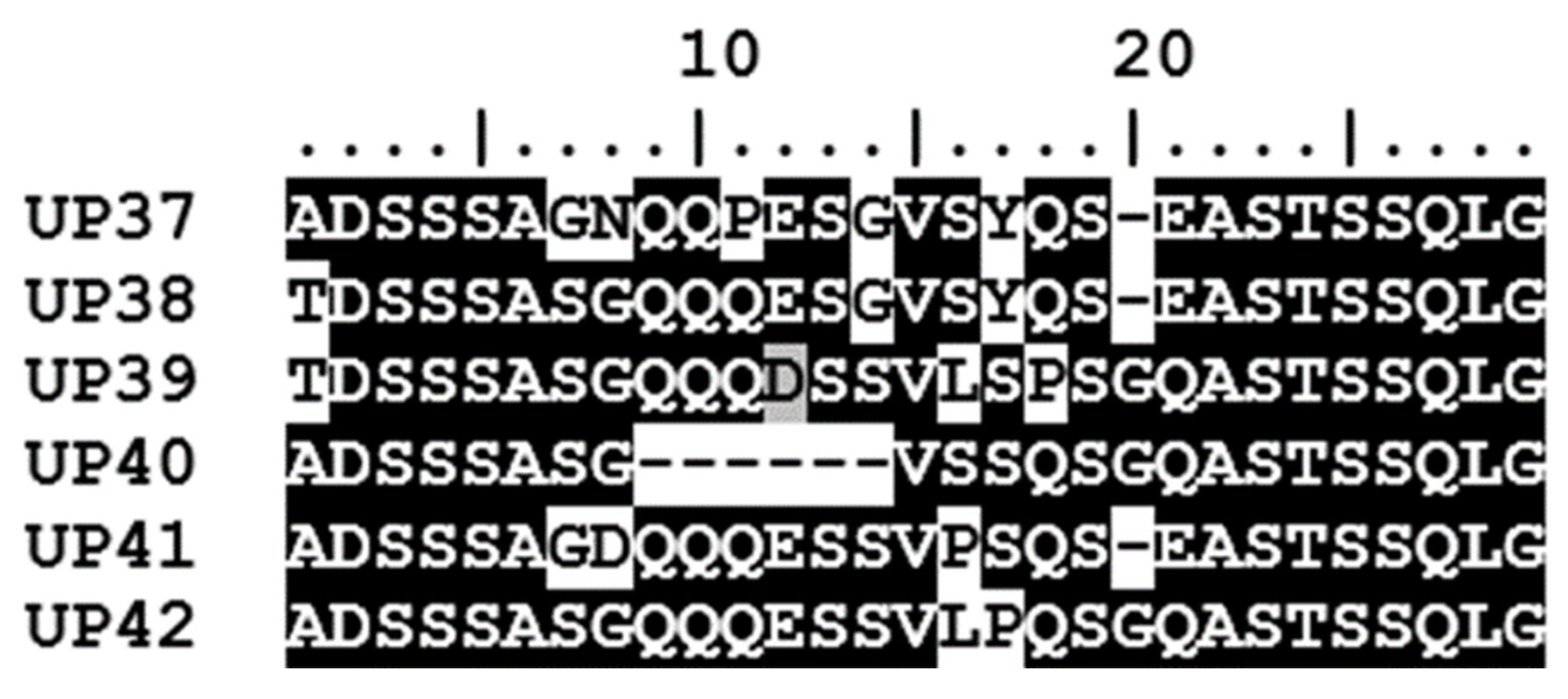
2.7. Characterization of A. marginale Msp1a Repeats and Msp1α Genotypes
2.8. 16S rRNA Gene Amplification and PacBio Sequencing
2.9. Analysis of Anaplasma 16S rRNA Sequences Identified by Microbiome Sequencing
2.10. Sequence and Phylogenetic Analysis
3. Results
3.1. Anaplasma Marginale Msp1α Genotype Analysis in the Calves for a Period of a Year
3.2. Occurrence of A. marginale Multi-Strain Infections in the Calves
3.3. The Composition of Anaplasma spp. in the Ten Calves
3.4. 16S rRNA Phylogenetic Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aubry, P.; Geale, D.W. A Review of Bovine Anaplasmosis. Transbound. Emerg. Dis. 2011, 58, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Kocan, K.M.; de la Fuente, J.; Guglielmone, A.A.; Meléndez, R.D. Antigens and Alternatives for Control of Anaplasma marginale Infection in Cattle. Clin. Microbiol. Rev. 2003, 16, 698–712. [Google Scholar] [CrossRef] [PubMed]
- Uilenberg, G. International Collaborative Research: Significance of Tick-Borne Hemoparasitic Diseases to World Animal Health. Vet. Parasitol. 1995, 57, 19–41. [Google Scholar] [CrossRef] [PubMed]
- De WaaL, D.T. Anaplasmosis Control and Diagnosis in South Africa. Annu. N. Y. Acad. Sci. 2000, 916, 474–483. [Google Scholar] [CrossRef]
- Makala, L.H.; Mangani, P.; Fujisaki, K.; Nagasawa, H. The Current Status of Major Tick-Borne Diseases in Zambia. Vet. Res. 2003, 34, 27–45. [Google Scholar] [CrossRef]
- Mtshali, M.S.; De Waal, D.T.; Mbati, P.A. A Sero-Epidemiological Survey of Blood Parasites in Cattle in the North-Eastern Free State, South Africa. Onderstepoort J. Vet. 2004, 71, 67–75. [Google Scholar] [CrossRef]
- Hove, P. Molecular Characterisation of Potential Vaccine Candidates from Anaplasma marginale Strains in South Africa. Ph.D. Dissertation, University of Pretoria, Pretoria, South Africa, 2018. [Google Scholar]
- Kocan, K.M.; De La Fuente, J.; Blouin, E.F.; Garcia-Garcia, J.C. Anaplasma marginale (Rickettsiales: Anaplasmataceae): Recent Advances in Defining Host–Pathogen Adaptations of a Tick-Borne Rickettsia. Parasitology 2004, 129, S285–S300. [Google Scholar] [CrossRef]
- Palmer, G.H. Sir Arnold Theiler and the Discovery of Anaplasmosis: A Centennial Perspective: Tick-Borne Diseases. Onderstepoort J. Vet. 2009, 76, 75–79. [Google Scholar] [CrossRef]
- Potgieter, F.T.; Stoltsz, W.H. Bovine Anaplasmosis. In Infectious Diseases of Livestock, Volume One, 2nd ed.; Oxford University Press: Cape Town, South Africa, 2004; pp. 594–616. [Google Scholar]
- Bock, R.; de Vos, A.; Kingston, T.; McLellan, D. Effect of Breed of Cattle on Innate Resistance to Infection with Babesia bovis, B. Bigemina and Anaplasma marginale. Aust. Vet. J. 1997, 75, 337–340. [Google Scholar] [CrossRef]
- Jonsson, N.N.; Bock, R.E.; Jorgensen, W.K.; Morton, J.M.; Stear, M.J. Is Endemic Stability of Tick-Borne Disease in Cattle a Useful Concept? Trends Parasitol. 2012, 28, 85–89. [Google Scholar] [CrossRef]
- de la Fuente, J.; Van Den Bussche, R.A.; Garcia-Garcia, J.C.; Rodríguez, S.D.; García, M.A.; Guglielmone, A.A.; Mangold, A.J.; Friche Passos, L.M.; Barbosa Ribeiro, M.F.; Blouin, E.F.; et al. Phylogeography of New World Isolates of Anaplasma marginale Based on Major Surface Protein Sequences. Vet. Microbiol. 2002, 88, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Kocan, K.M.; Blouin, E.F.; Barbet, A.F. Anaplasmosis Control: Past, Present, and Future. Ann. N. Y. Acad. Sci. 2000, 916, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.C.L.; de Magalhães, V.C.S.; de Oliveira, U.V.; Carvalho, F.S.; de Almeida, C.P.; Machado, R.Z.; Munhoz, A.D. Transplacental Transmission of Bovine Tick-Borne Pathogens: Frequency, Co-Infections and Fatal Neonatal Anaplasmosis in a Region of Enzootic Stability in the Northeast of Brazil. Ticks Tick Borne Dis. 2016, 7, 270–275. [Google Scholar] [CrossRef]
- Nyangiwe, N.; Harrison, A.; Horak, I.G. Displacement of Rhipicephalus decoloratus by Rhipicephalus microplus (Acari: Ixodidae) in the Eastern Cape Province, South Africa. Exp. Appl. Acarol. 2013, 61, 371–382. [Google Scholar] [CrossRef]
- Nyangiwe, N.; Horak, I.G.; Van der Mescht, L.; Matthee, S. Range Expansion of the Economically Important Asiatic Blue Tick, Rhipicephalus microplus, in South Africa. J. S. Afr. Vet. Assoc. 2017, 88, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Palmer, G.H.; Rurangirwa, F.R.; Kocan, K.M.; Brown, W.C. Molecular Basis for Vaccine Development against the Ehrlichial Pathogen Anaplasma marginale. Trends Parasitol. 1999, 15, 281–286. [Google Scholar] [CrossRef]
- de la Fuente, J.; Massung Robert, F.; Wong Susan, J.; Chu Frederick, K.; Lutz, H.; Meli, M.; von Loewenich, F.D.; Grzeszczuk, A.; Torina, A.; Caracappa, S.; et al. Sequence Analysis of the Msp4 Gene of Anaplasma phagocytophilum Strains. J. Clin. Microbiol. 2005, 43, 1309–1317. [Google Scholar] [CrossRef]
- de la Fuente, J.; Ruybal, P.; Mtshali, M.S.; Naranjo, V.; Shuqing, L.; Mangold, A.J.; Rodríguez, S.D.; Jiménez, R.; Vicente, J.; Moretta, R.; et al. Analysis of World Strains of Anaplasma marginale Using Major Surface Protein 1a Repeat Sequences. Vet. Microbiol. 2007, 119, 382–390. [Google Scholar] [CrossRef]
- Allred, D.R.; McGuire, T.C.; Palmer, G.H.; Leib, S.R.; Harkins, T.M.; McElwain, T.F.; Barbet, A.F. Molecular Basis for Surface Antigen Size Polymorphisms and Conservation of a Neutralization-Sensitive Epitope in Anaplasma marginale. Proc. Natl. Acad. Sci. USA 1990, 87, 3220–3224. [Google Scholar] [CrossRef]
- Catanese, H.N.; Brayton, K.A.; Gebremedhin, A.H. RepeatAnalyzer: A Tool for Analysing and Managing Short-Sequence Repeat Data. BMC Genom. 2016, 17, 422. [Google Scholar] [CrossRef]
- Mtshali, M.S.; De La Fuente, J.; Ruybal, P.; Kocan, K.M.; Vicente, J.; Mbati, P.A.; Shkap, V.; Blouin, E.F.; Mohale, N.E.; Moloi, T.P.; et al. Prevalence and Genetic Diversity of Anaplasma marginale Strains in Cattle in South Africa. Zoonoses Public Health 2007, 54, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Mutshembele, A.M.; Cabezas-Cruz, A.; Mtshali, M.S.; Thekisoe, O.M.M.; Galindo, R.C.; de la Fuente, J. Epidemiology and Evolution of the Genetic Variability of Anaplasma marginale in South Africa. Ticks Tick Borne Dis. 2014, 5, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Hove, P.; Chaisi, M.E.; Brayton, K.A.; Ganesan, H.; Catanese, H.N.; Mtshali, M.S.; Mutshembele, A.M.; Oosthuizen, M.C.; Collins, N.E. Co-Infections with Multiple Genotypes of Anaplasma marginale in Cattle Indicate Pathogen Diversity. Parasit. Vectors 2018, 11, 5. [Google Scholar] [CrossRef] [PubMed]
- de la Fuente, J.; Garcia-Garcia, J.C.; Blouin, E.F.; McEwen, B.R.; Clawson, D.; Kocan, K.M. Major Surface Protein 1a Effects Tick Infection and Transmission of Anaplasma marginale. Int. J. Parasitol. 2001, 31, 1705–1714. [Google Scholar] [CrossRef] [PubMed]
- Palmer, G.H.; Knowles, D.P.; Rodriguez, J.-L.; Gnad, D.P.; Hollis, L.C.; Marston, T.; Brayton, K.A. Stochastic Transmission of Multiple Genotypically Distinct Anaplasma marginale Strains in a Herd with High Prevalence of Anaplasma Infection. J. Clin. Microbiol. 2004, 42, 5381–5384. [Google Scholar] [CrossRef]
- Futse, J.E.; Brayton, K.A.; Dark, M.J.; Knowles, D.P.; Palmer, G.H. Superinfection as a Driver of Genomic Diversification in Antigenically Variant Pathogens. Proc. Natl. Acad. Sci. USA 2008, 105, 2123–2127. [Google Scholar] [CrossRef]
- Leverich, C.K.; Palmer, G.H.; Knowles, D.P.; Brayton, K.A. Tick-Borne Transmission of Two Genetically Distinct Anaplasma marginale Strains Following Superinfection of the Mammalian Reservoir Host. Infect. Immun. 2008, 76, 4066–4070. [Google Scholar] [CrossRef]
- Castañeda-Ortiz, E.J.; Ueti, M.W.; Camacho-Nuez, M.; Mosqueda, J.J.; Mousel, M.R.; Johnson, W.C.; Palmer, G.H. Association of Anaplasma marginale Strain Superinfection with Infection Prevalence within Tropical Regions. PLoS ONE 2015, 10, e0120748. [Google Scholar] [CrossRef]
- Koku, R.; Herndon, D.R.; Avillan, J.; Morrison, J.; Futse, J.E.; Palmer, G.H.; Brayton, K.A.; Noh, S.M.; Herbert, D.R. Both Coinfection and Superinfection Drive Complex Anaplasma marginale Strain Structure in a Natural Transmission Setting. Infect. Immun. 2021, 89, e00166-21. [Google Scholar] [CrossRef]
- Choopa, C.N. Diagnosis of Tick-Borne Diseases in Cattle in Bushbuckridge Mpumalanga South Africa and Identification of Theileria parva Carriers. Master’s Thesis, University of Pretoria, Pretoria, South Africa, 2015. [Google Scholar]
- Kolo, A.O.; Collins, N.E.; Brayton, K.A.; Chaisi, M.; Blumberg, L.; Frean, J.; Gall, C.A.; Wentzel, M.J.; Wills-Berriman, S.; Boni, L.D.; et al. Anaplasma phagocytophilum and Other Anaplasma spp. in Various Hosts in the Mnisi Community, Mpumalanga Province, South Africa. Microorganisms 2020, 8, 1812. [Google Scholar] [CrossRef]
- Chaisi, M.E.; Baxter, J.R.; Hove, P.; Choopa, C.N.; Oosthuizen, M.C.; Brayton, K.A.; Khumalo, Z.T.H.; Mutshembele, A.M.; Mtshali, M.S.; Collins, N.E. Comparison of Three Nucleic Acid-Based Tests for Detecting Anaplasma marginale and Anaplasma centrale in Cattle. Onderstepoort J. Vet. 2017, 84, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lew, A.E.; Bock, R.E.; Minchin, C.M.; Masaka, S. A msp1α Polymerase Chain Reaction Assay for Specific Detection and Differentiation of Anaplasma marginale Isolates. Vet. Microbiol. 2002, 86, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Lane, D. 16S/23S rRNA sequencing. In Nucleic Acid Sequencing Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley & Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Turner, S.; Pryer, K.M.; Miao, V.P.W.; Palmer, J.D. Investigating Deep Phylogenetic Relationships among Cyanobacteria and Plastids by Small Subunit rRNA Sequence Analysis. J. Eukaryot. Microbiol. 1999, 46, 327–338. [Google Scholar] [CrossRef]
- Cole, J.R.; Wang, Q.; Cardenas, E.; Fish, J.; Chai, B.; Farris, R.J.; Kulam-Syed-Mohideen, A.S.; McGarrell, D.M.; Marsh, T.; Garrity, G.M.; et al. The Ribosomal Database Project: Improved Alignments and New Tools for rRNA Analysis. Nucleic Acids Res. 2009, 37, D141–D145. [Google Scholar] [CrossRef]
- Gall, C.A.; Reif, K.E.; Scoles, G.A.; Mason, K.L.; Mousel, M.; Noh, S.M.; Brayton, K.A. The Bacterial Microbiome of Dermacentor andersoni Ticks Influences Pathogen Susceptibility. ISME J. 2016, 10, 1846–1855. [Google Scholar] [CrossRef] [PubMed]
- Caudill, M.T.; Brayton, K.A. The Use and Limitations of the 16S rRNA Sequence for Species Classification of Anaplasma Samples. Microorganisms 2022, 10, 605. [Google Scholar] [CrossRef]
- Makgabo, S.M.; Brayton, K.A.; Oosthuizen, M.C.; Collins, N.E. Unravelling the diversity of Anaplasma species circulating in selected African wildlife hosts by targeted 16S microbiome analysis. Curr. Res. Microb. Sci. (Submitted, Under Review).
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More Models, New Heuristics and Parallel Computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Guindon, S.; Gascuel, O. A Simple, Fast, and Accurate Algorithm to Estimate Large Phylogenies by Maximum Likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef]
- Anisimova, M.; Gascuel, O. Approximate Likelihood-Ratio Test for Branches: A Fast, Accurate, and Powerful Alternative. Syst. Biol. 2006, 55, 539–552. [Google Scholar] [CrossRef] [PubMed]
- Chevenet, F.; Brun, C.; Bañuls, A.-L.; Jacq, B.; Christen, R. TreeDyn: Towards Dynamic Graphics and Annotations for Analyses of Trees. BMC Bioinform. 2006, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Kieser, S.T.; Eriks, I.S.; Palmer, G.H. Cyclic Rickettsemia during Persistent Anaplasma marginale Infection of Cattle. Infect. Immun. 1990, 58, 1117–1119. [Google Scholar] [CrossRef]
- Brayton, K.A.; Kappmeyer, L.S.; Herndon, D.R.; Dark, M.J.; Tibbals, D.L.; Palmer, G.H.; McGuire, T.C.; Knowles Jr, D.P. Complete Genome Sequencing of Anaplasma marginale Reveals That the Surface Is Skewed to Two Superfamilies of Outer Membrane Proteins. Proc. Natl. Acad. Sci. USA 2005, 102, 844–849. [Google Scholar] [CrossRef]
- Guo, W.-P.; Tian, J.-H.; Lin, X.-D.; Ni, X.-B.; Chen, X.-P.; Liao, Y.; Yang, S.-Y.; Dumler, J.S.; Holmes, E.C.; Zhang, Y.-Z. Extensive Genetic Diversity of Rickettsiales Bacteria in Multiple Mosquito Species. Sci. Rep. 2016, 6, 38770. [Google Scholar] [CrossRef]
- Allsopp, M.T.E.P.; Visser, E.S.; du Plessis, J.L.; Vogel, S.W.; Allsopp, B.A. Different Organisms Associated with Heartwater as Shown by Analysis of 16S Ribosomal RNA Gene Sequences. Vet. Parasitol. 1997, 71, 283–300. [Google Scholar] [CrossRef]
- Roy, B.; Krücken, J.; Ahmed, J.; Majumder, S.; Baumann, M.; Clausen, P.; Nijhof, A. Molecular Identification of Tick-borne Pathogens Infecting Cattle in Mymensingh District of Bangladesh Reveals Emerging Species of Anaplasma and Babesia. Transbound. Emerg. Dis. 2018, 65, e231–e242. [Google Scholar] [CrossRef]
- Bastos, A.D.S.; Mohammed, O.B.; Bennett, N.C.; Petevinos, C.; Alagaili, A.N. Molecular Detection of Novel Anaplasmataceae Closely Related to Anaplasma platys and Ehrlichia canis in the Dromedary Camel (Camelus dromedarius). Vet. Microbiol. 2015, 179, 310–314. [Google Scholar] [CrossRef]
- Potgieter, F.T. Epizootiology and Control of Anaplasmosis in South Africa. J. S. Afr. Vet. Assoc. 1979, 50, 367–372. [Google Scholar] [CrossRef]
- Henrichs, B.; Oosthuizen, M.C.; Troskie, M.; Gorsich, E.; Gondhalekar, C.; Beechler, B.R.; Ezenwa, V.O.; Jolles, A.E. Within Guild Co-Infections Influence Parasite Community Membership: A Longitudinal Study in African Buffalo. J. Anim. Ecol. 2016, 85, 1025–1034. [Google Scholar] [CrossRef] [PubMed]
- Khumalo, Z.T.; Catanese, H.N.; Liesching, N.; Hove, P.; Collins, N.E.; Chaisi, M.E.; Gebremedhin, A.H.; Oosthuizen, M.C.; Brayton, K.A. Characterization of Anaplasma marginale Subsp. centrale Strains by Use of Msp1aS Genotyping Reveals a Wildlife Reservoir. J. Clin. Microbiol. 2016, 54, 2503–2512. [Google Scholar] [CrossRef] [PubMed]
- Sisson, D.; Hufschmid, J.; Jolles, A.; Beechler, B.; Jabbar, A. Molecular Characterisation of Anaplasma Species from African Buffalo (Syncerus caffer) in Kruger National Park, South Africa. Ticks Tick Borne Dis. 2017, 8, 400–406. [Google Scholar] [CrossRef]
- Mason, C.A.; Norval, R.A.I. The Transfer of Boophilus microplus (Acarina: Ixodidae) from Infested to Uninfested Cattle under Field Conditions. Vet. Parasitol. 1981, 8, 185–188. [Google Scholar] [CrossRef]
- Aguirre, D.H.; Gaido, A.B.; Vinabal, A.E.; Echaide, S.T.D.; Guglielmone, A.A. Transmission of Anaplasma marginale with Adult Boophilus microplus Ticks Fed as Nymphs on Calves with Different Levels of Rickettsaemia. Parasite 1994, 1, 405–407. [Google Scholar] [CrossRef]
- Scoles, G.A.; Broce, A.B.; Lysyk, T.J.; Palmer, G.H. Relative Efficiency of Biological Transmission of Anaplasma marginale (Rickettsiales: Anaplasmataceae) by Dermacentor andersoni (Acari: Ixodidae) Compared with Mechanical Transmission by Stomoxys calcitrans (Diptera: Muscidae). J. Med. Entomol. 2005, 42, 668–675. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, N. Control of Cattle Ticks (Boophilus microplus) on Queensland Dairy Farms. Aust. Vet. J. 1997, 75, 802–807. [Google Scholar] [CrossRef]
- Sungirai, M.; Moyo, D.Z.; De Clercq, P.; Madder, M. Communal Farmers’ Perceptions of Tick-Borne Diseases Affecting Cattle and Investigation of Tick Control Methods Practiced in Zimbabwe. Ticks Tick Borne Dis. 2016, 7, 1–9. [Google Scholar] [CrossRef]
- Woolhouse, M.E.; Thumbi, S.M.; Jennings, A.; Chase-Topping, M.; Callaby, R.; Kiara, H.; Oosthuizen, M.C.; Mbole-Kariuki, M.N.; Conradie, I.; Handel, I.G. Co-Infections Determine Patterns of Mortality in a Population Exposed to Parasite Infection. Sci. Adv. 2015, 1, e1400026. [Google Scholar] [CrossRef]
- Löhr, K. Game Anaplasmosis: The Isolation of Anaplasma Organisms from Antelope. Zeitschr. Tropenmed. Parasitol. 1973, 24, 192–197. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Calf No.a | No. of Genotypes | Size (bp) | No. of Msp1a Repeats | Genotype | Number Allocated to Genotype | Genotype #, If Previously Detected in Study |
|---|---|---|---|---|---|---|
| 1 (EG1) | 13 | 949 | 5 | 171-2;UP3 b 172-2;UP4 61 172-2;UP4 172-2;UP4 | 1 | |
| 700 | 2 | τ 10 | 2 | |||
| 697 | 2 | 171-2;UP3 172-2;UP4 | 3 | |||
| 866 | 3 | 171-2;UP3 172-2;UP4 61 | 4 | |||
| 836 | 6 | UP37 c UP31 UP31 UP31 UP31 UP31 | 5 | |||
| 781 | 2 | τ UP31 | 6 | |||
| 787 | 3 | τ 10 22-2 | 7 | |||
| 610 | 1 | UP38 c | 8 | |||
| 893 | 5 | 61 172-2;UP4 61 172-2;UP4 172-2;UP4 | 9 | |||
| 781 | 3 | 61 172-2;UP4 169-2 | 10 | |||
| 781 | 3 | 61 172-2;UP4 172-2;UP4 | 11 | |||
| 781 | 3 | 171-2;UP3 172-2;UP4 172-2;UP4 | 12 | |||
| 697 | 2 | 172-2;UP4 172-2;UP4 | 13 | |||
| 2 (EG2) | 4 | 784 | 3 | UP39 c 10 UP31 | 14 | 5, 7 |
| 781 | 3 | 179-2 169-2 172-2;UP4 | 15 | |||
| 3 (EG3) | 4 | 787 | 3 | 84 172-2;UP4 172-2;UP4 | 16 d | |
| 959 | 5 | 34 3 36 36 38 | 17 d | |||
| 958 | 5 | 13 27 36 3 38 | 18 | |||
| 700 | 2 | 13 27 | 19 | |||
| 4 (EG4) | 12 | 1040 | 6 | 34 36 36 3 36 38 | 20 | 17 d |
| 1037 | 6 | UP40 c β β β β F | 21 | |||
| 959 | 5 | 34 36 36 27 18 | 22 | |||
| 954 | 5 | MZ2 3 UP41 c 36 38 | 23 | |||
| 880 | 4 | 3 β 36 3 | 24 | |||
| 962 | 5 | 42 43 43 25 31 | 25 | |||
| 1131 | 7 | 34 3 UP1 43 43 25 31 | 26 | |||
| 1026 | 6 | UP40 c β β β Is9;78 31 | 27 | |||
| 965 | 5 | 84 172-2;UP4 172-2;UP4 172-2;UP4 172-2;UP4 | 28 | |||
| 870 | 4 | τ 22-2 13 18 | 34 b | |||
| 705 | 2 | 34 3 | 41 | |||
| 5 (EG5) | 9 | 689 | 2 | UP40 c β | 29 | 17 d, 21, 41 |
| 1001 | 5 | UP40 c β β β F | 30 | |||
| 875 | 4 | 42 43 25 31 | 31 | |||
| 790 | 3 | 42 UP42 c 27 | 32 | |||
| 791 | 3 | H M 27 | 33 c | |||
| 602 | 1 | UP40 c | 42 | |||
| 6 (UT1) | 4 | 919 | 5 | UP5 UP6 25 31 31 | 35 | 17 d, 33 d, 34 d |
| 7 (UT2) | 9 | 1075 | 7 | UP5 UP6 25 31 UP6 27 18 | 36 | 16 d, 33 d, 34 d, 35 |
| 787 | 3 | 84 61 31 | 37 | |||
| 863 | 4 | UP5 UP6 25 31 | 38 | |||
| 1202 | 8 | UP5 UP6 25 31 UP6 25 31 31 | 39 | |||
| 955 | 4 | 84 Is9;78 31 31 | 40 |
| Calf No. | A. platys-like | A. marginale | A. boleense |
|---|---|---|---|
| 1 (EG1) | 67.3 | 32.3 | 0.4 |
| 2 (EG2) | 98.7 | 1.3 | 0 |
| 3 (EG3) | 56.0 | 43.9 | 0.1 |
| 4 (EG4) | 0 | 100 | 0 |
| 5 (EG5) | 68.5 | 31.3 | 0.2 |
| 6 (UT1) | 7.0 | 93.0 | 0 |
| 7 (UT2) | 95.3 | 4.7 | 0 |
| 8 (DI1) | 0 | 0 | 0 |
| 9 (DI2) | 0 | 0 | 0 |
| 10 (DI3) | 100 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Makgabo, S.M.; Brayton, K.A.; Biggs, L.; Oosthuizen, M.C.; Collins, N.E. Temporal Dynamics of Anaplasma marginale Infections and the Composition of Anaplasma spp. in Calves in the Mnisi Communal Area, Mpumalanga, South Africa. Microorganisms 2023, 11, 465. https://doi.org/10.3390/microorganisms11020465
Makgabo SM, Brayton KA, Biggs L, Oosthuizen MC, Collins NE. Temporal Dynamics of Anaplasma marginale Infections and the Composition of Anaplasma spp. in Calves in the Mnisi Communal Area, Mpumalanga, South Africa. Microorganisms. 2023; 11(2):465. https://doi.org/10.3390/microorganisms11020465
Chicago/Turabian StyleMakgabo, S. Marcus, Kelly A. Brayton, Louise Biggs, Marinda C. Oosthuizen, and Nicola E. Collins. 2023. "Temporal Dynamics of Anaplasma marginale Infections and the Composition of Anaplasma spp. in Calves in the Mnisi Communal Area, Mpumalanga, South Africa" Microorganisms 11, no. 2: 465. https://doi.org/10.3390/microorganisms11020465
APA StyleMakgabo, S. M., Brayton, K. A., Biggs, L., Oosthuizen, M. C., & Collins, N. E. (2023). Temporal Dynamics of Anaplasma marginale Infections and the Composition of Anaplasma spp. in Calves in the Mnisi Communal Area, Mpumalanga, South Africa. Microorganisms, 11(2), 465. https://doi.org/10.3390/microorganisms11020465