
Microorganisms in Fish Feeds, Technological Innovations, and Key Strategies for Sustainable Aquaculture

Abstract
1. Significance of Aquaculture and Aquafeed
1.1. Use of Fishmeal and Fish Oil in Aquafeed
1.2. Use of Terrestrial Crops and Oils in Aquafeeds and Environmental Challenges
1.2.1. Nutritional Disadvantages of Terrestrial Crop Protein
1.2.2. Nutritional Disadvantages of Terrestrial Crop Oil
1.2.3. Fillet Quality for Human Consumption
2. Novel Aquafeed Ingredients
2.1. Microalgae Protein and Oil

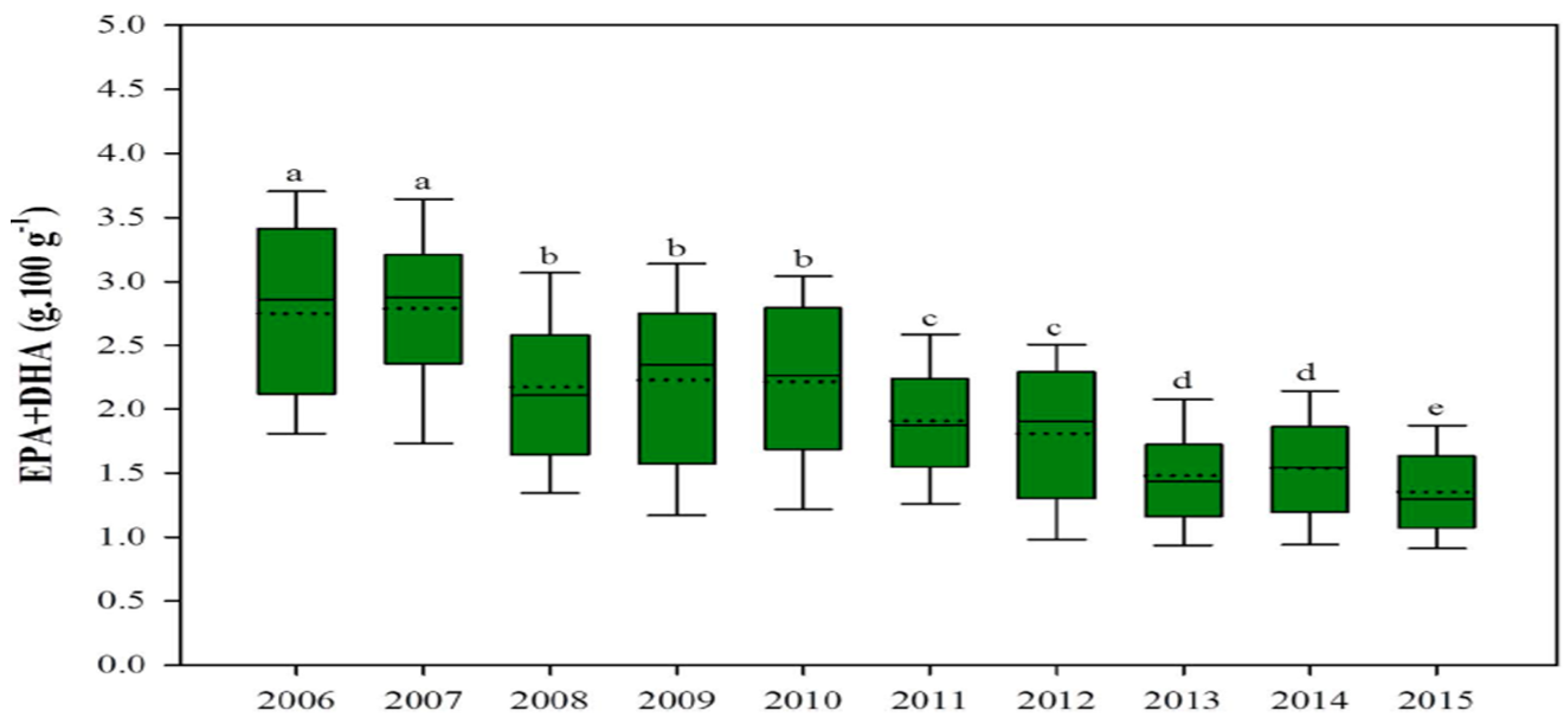{kind=link}
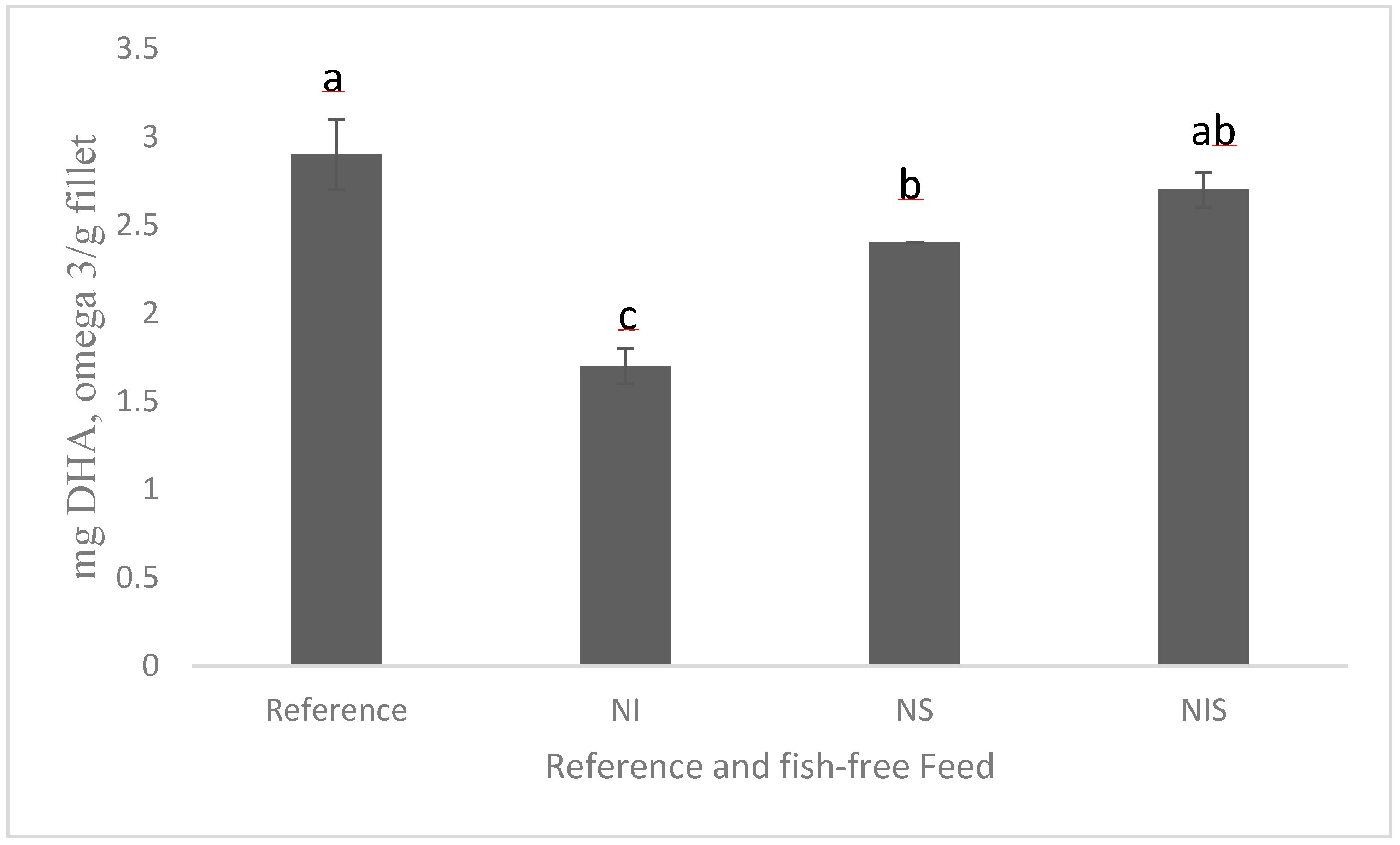{kind=link}
| Microalgae | Fish Species | FCR | Inclusion % of Diet | Reference |
|---|---|---|---|---|
| Schizochytrium sp. | Atlantic salmon | 1.40 and 1.42 | 5.5 and 11 | [105] |
| Schizochytrium sp. | Tilapia | 1.0, 1.0, 0.9 and 0.9 | 4, 8, 12.5, and 16.1 | [97] |
| Schizochytrium sp. | Tilapia | 1.57, 1.60 and 1.4 | 6.20 | [29] |
| Schizochytrium sp. | Rainbow trout | 1.0 and 0.98 | 9 and 6.5 | [41] |
| Schizochytrium sp. | Pacific white shrimp | Not reported | 0.6, 1.2, 1.8, 2.3 and 3.5 | [106] |
| Schizochytrium sp. | Atlantic salmon | 0.72 and 0.73 | 2.5 and 5.0 | [107] |
| Schizochytrium sp. | Rainbow trout | 0.98 | 2.50 | [100] |
| Isochrysis sp | Rainbow trout | 1.0 | 2.40 | [100] |
| Isochrysis sp | European sea bass | 1.69 and 1.76 | 7 and 14 | [108] |
2.2. Yeast, Fungi, and Bacteria
2.3. Genetically Modified Oil
2.4. Insect Meal
2.5. Fish Processing Byproduct
3. Key Strategies to Measure Nutritional and Environmental Sustainability
3.1. Ingredients Digestibility
3.2. Feed Conversion Ratio (FCR)
3.3. Life Cycle Assessment (LCA) for Ecological Impact Measures
4. Novel Technology for Improving Ingredients Quality
4.1. Extrusion Processing
4.2. Use of Exogenous Enzymes in Aquafeed
4.3. Use of Additives in Aquafeed to Improve Palatability
5. Challenges with Adopting and Opportunities to Adopt the New Aquafeeds
6. Conclusions and Future Steps
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2016: Contributing to Food Security and Nutrition for All; FAO: Rome, Italy, 2016; ISBN 978-92-5-109185-2. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture 2018: Meeting the Sustainable Development Goals; FAO: Rome, Italy, 2018; ISBN 978-92-5-130562-1. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture 2020: Sustainability in Action; FAO: Rome, Italy, 2020; ISBN 978-92-5-132692-3. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture 2022; FAO: Rome, Italy, 2022. [Google Scholar]
- Rockström, J.; Williams, J.; Daily, G.; Noble, A.; Matthews, N.; Gordon, L.; Wetterstrand, H.; DeClerck, F.; Shah, M.; Steduto, P.; et al. Sustainable Intensification of Agriculture for Human Prosperity and Global Sustainability. Ambio 2017, 46, 4–17. [Google Scholar] [CrossRef] [PubMed]
- Willett, W.; Rockström, J.; Loken, B.; Springmann, M.; Lang, T.; Vermeulen, S.; Garnett, T.; Tilman, D.; DeClerck, F.; Wood, A.; et al. Food in the Anthropocene: The EAT–Lancet Commission on Healthy Diets from Sustainable Food Systems. Lancet 2019, 393, 447–492. [Google Scholar] [CrossRef] [PubMed]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food Security: The Challenge of Feeding 9 Billion People. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Sarker, P.K.; Bureau, D.P.; Hua, K.; Drew, M.D.; Forster, I.; Were, K.; Hicks, B.; Vandenberg, G.W. Sustainability Issues Related to Feeding Salmonids: A Canadian Perspective. Rev. Aquac. 2013, 5, 199–219. [Google Scholar] [CrossRef]
- Fry, J.P.; Mailloux, N.A.; Love, D.C.; Milli, M.C.; Cao, L. Feed Conversion Efficiency in Aquaculture: Do We Measure It Correctly? Environ. Res. Lett. 2018, 13, 024017. [Google Scholar] [CrossRef]
- Froehlich, H.E.; Jacobsen, N.S.; Essington, T.E.; Clavelle, T.; Halpern, B.S. Avoiding the Ecological Limits of Forage Fish for Fed Aquaculture. Nat. Sustain. 2018, 1, 298–303. [Google Scholar] [CrossRef]
- Hua, K.; Cobcroft, J.M.; Cole, A.; Condon, K.; Jerry, D.R.; Mangott, A.; Praeger, C.; Vucko, M.J.; Zeng, C.; Zenger, K.; et al. The Future of Aquatic Protein: Implications for Protein Sources in Aquaculture Diets. One Earth 2019, 1, 316–329. [Google Scholar] [CrossRef]
- Tacon, A.G.J. Trends in Global Aquaculture and Aquafeed Production: 2000–2017. Rev. Fish. Sci. Aquac. 2020, 28, 43–56. [Google Scholar] [CrossRef]
- Browdy, C.L.; Hargreaves, J.A. Overcoming Technical Barriers to the Sustainable Development of Competitive Marine Aquaculture in the United States; NOAA Technical Memo NMFS F/SPO100; U.S. Department of Commerce: Silver Spring, MD, USA, 2009; 114p.
- McKuin, B.L.; Kapuscinski, A.R.; Sarker, P.K.; Cheek, N.; Colwell, A.; Schoffstall, B.; Greenwood, C. Comparative Life Cycle Assessment of Heterotrophic Microalgae Schizochytrium and Fish Oil in Sustainable Aquaculture Feeds. Elem. Sci. Anthr. 2022, 10, 00098. [Google Scholar] [CrossRef]
- Hilborn, R.; Banobi, J.; Hall, S.J.; Pucylowski, T.; Walsworth, T.E. The Environmental Cost of Animal Source Foods. Front. Ecol. Environ. 2018, 16, 329–335. [Google Scholar] [CrossRef]
- Parodi, A.; Leip, A.; De Boer, I.J.M.; Slegers, P.M.; Ziegler, F.; Temme, E.H.M.; Herrero, M.; Tuomisto, H.; Valin, H.; Van Middelaar, C.E.; et al. The Potential of Future Foods for Sustainable and Healthy Diets. Nat. Sustain. 2018, 1, 782–789. [Google Scholar] [CrossRef]
- MacLeod, M.J.; Hasan, M.R.; Robb, D.H.F.; Mamun-Ur-Rashid, M. Quantifying Greenhouse Gas Emissions from Global Aquaculture. Sci. Rep. 2020, 10, 11679. [Google Scholar] [CrossRef]
- Ellingsen, H.; Aanondsen, S.A. Environmental Impacts of Wild Caught Cod and Farmed Salmon—A Comparison with Chicken (7 Pp). Int. J. Life Cycle Assess. 2006, 11, 60–65. [Google Scholar] [CrossRef]
- Pelletier, N.; Tyedmers, P. Feeding Farmed Salmon: Is Organic Better? Aquaculture 2007, 272, 399–416. [Google Scholar] [CrossRef]
- Iribarren, D.; Vázquez-Rowe, I.; Hospido, A.; Moreira, M.T.; Feijoo, G. Estimation of the Carbon Footprint of the Galician Fishing Activity (NW Spain). Sci. Total Environ. 2010, 408, 5284–5294. [Google Scholar] [CrossRef]
- Parker, R. Implications of High Animal By-Product Feed Inputs in Life Cycle Assessments of Farmed Atlantic Salmon. Int. J. Life Cycle Assess. 2018, 23, 982–994. [Google Scholar] [CrossRef]
- Jones, A.R.; Alleway, H.K.; McAfee, D.; Reis-Santos, P.; Theuerkauf, S.J.; Jones, R.C. Climate-Friendly Seafood: The Potential for Emissions Reduction and Carbon Capture in Marine Aquaculture. BioScience 2022, 72, 123–143. [Google Scholar] [CrossRef]
- Asche, F.; Cojocaru, A.L.; Roth, B. The Development of Large Scale Aquaculture Production: A Comparison of the Supply Chains for Chicken and Salmon. Aquaculture 2018, 493, 446–455. [Google Scholar] [CrossRef]
- Han, D.; Shan, X.; Zhang, W.; Chen, Y.; Wang, Q.; Li, Z.; Zhang, G.; Xu, P.; Li, J.; Xie, S.; et al. A Revisit to Fishmeal Usage and Associated Consequences in Chinese Aquaculture. Rev. Aquac. 2018, 10, 493–507. [Google Scholar] [CrossRef]
- Little, D.C.; Young, J.A.; Zhang, W.; Newton, R.W.; Al Mamun, A.; Murray, F.J. Sustainable Intensification of Aquaculture Value Chains between Asia and Europe: A Framework for Understanding Impacts and Challenges. Aquaculture 2018, 493, 338–354. [Google Scholar] [CrossRef]
- Xu, C.; Su, G.; Zhao, K.; Xu, X.; Li, Z.; Hu, Q.; Xue, Y.; Xu, J. Current Status of Greenhouse Gas Emissions from Aquaculture in China. Water Biol. Secur. 2022, 1, 100041. [Google Scholar] [CrossRef]
- Ballester-Moltó, M.; Sanchez-Jerez, P.; Cerezo-Valverde, J.; Aguado-Giménez, F. Particulate Waste Outflow from Fish-Farming Cages. How Much Is Uneaten Feed? Mar. Pollut. Bull. 2017, 119, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Maiolo, S.; Parisi, G.; Biondi, N.; Lunelli, F.; Tibaldi, E.; Pastres, R. Fishmeal Partial Substitution within Aquafeed Formulations: Life Cycle Assessment of Four Alternative Protein Sources. Int. J. Life Cycle Assess. 2020, 25, 1455–1471. [Google Scholar] [CrossRef]
- Sarker, P.K.; Kapuscinski, A.R.; McKuin, B.; Fitzgerald, D.S.; Nash, H.M.; Greenwood, C. Microalgae-Blend Tilapia Feed Eliminates Fishmeal and Fish Oil, Improves Growth, and Is Cost Viable. Sci. Rep. 2020, 10, 19328. [Google Scholar] [CrossRef]
- Arru, B.; Furesi, R.; Gasco, L.; Madau, F.; Pulina, P. The Introduction of Insect Meal into Fish Diet: The First Economic Analysis on European Sea Bass Farming. Sustainability 2019, 11, 1697. [Google Scholar] [CrossRef]
- Ekmekci, H.; Gül, M. Economic Structure and Problems of Trout Enterprises: A Case of Fethiye. Turk. J. Agric. Food Sci. Technol. 2017, 5, 33–42. [Google Scholar] [CrossRef]
- Global Market Insights Inc. Aquafeed Market Size Worth USD 185 Billion by 2023: Global Market Insights Inc. Available online: http://www.globenewswire.com/news-release/2016/07/06/853958/0/en/Aquafeed-Market-size-worth-USD-185-Billion-by-2023-Global-Market-Insights-Inc.html (accessed on 23 September 2020).
- Cottrell, R.S.; Blanchard, J.L.; Halpern, B.S.; Metian, M.; Froehlich, H.E. Global Adoption of Novel Aquaculture Feeds Could Substantially Reduce Forage Fish Demand by 2030. Nat. Food 2020, 1, 301–308. [Google Scholar] [CrossRef]
- Shepherd, C.J.; Jackson, A.J. Global Fishmeal and Fish-Oil Supply: Inputs, Outputs and Markets. J. Fish Biol. 2013, 83, 1046–1066. [Google Scholar] [CrossRef]
- Tacon, A.G.J.; Hasan, M.R.; Metian, M. Demand and Supply of Feed Ingredients for Farmed Fish and Crustaceans; Food and Agriculture Organization of the United Nations: Rome, Italy, 2011; ISBN 978-92-5-106933-2. [Google Scholar]
- Hasan, M.R.; Soto, D. Improving Feed Conversion Ratio and Its Impact on Reducing Greenhouse Gas Emissions in Aquaculture; Food and Agriculture Organization of the United Nations: Rome, Italy, 2017. [Google Scholar]
- Jones, S.W.; Karpol, A.; Friedman, S.; Maru, B.T.; Tracy, B.P. Recent Advances in Single Cell Protein Use as a Feed Ingredient in Aquaculture. Curr. Opin. Biotechnol. 2020, 61, 189–197. [Google Scholar] [CrossRef]
- Klinger, D.H.; Naylor, R. Searching for Solutions in Aquaculture: Charting a Sustainable Course. Annu. Rev. Environ. Resour. 2012, 37, 247–276. [Google Scholar] [CrossRef]
- Chauton, M.S.; Reitan, K.I.; Norsker, N.H.; Tveterås, R.; Kleivdal, H.T. A Techno-Economic Analysis of Industrial Production of Marine Microalgae as a Source of EPA and DHA-Rich Raw Material for Aquafeed: Research Challenges and Possibilities. Aquaculture 2015, 436, 95–103. [Google Scholar] [CrossRef]
- Hardy, R.W. Utilization of Plant Proteins in Fish Diets: Effects of Global Demand and Supplies of Fishmeal. Aquac. Res. 2010, 41, 770–776. [Google Scholar] [CrossRef]
- Bélanger-Lamonde, A.; Sarker, P.K.; Ayotte, P.; Bailey, J.L.; Bureau, D.P.; Chouinard, P.Y.; Dewailly, É.; Leblanc, A.; Weber, J.-P.; Vandenberg, G.W. Algal and Vegetable Oils as Sustainable Fish Oil Substitutes in Rainbow Trout Diets: An Approach to Reduce Contaminant Exposure. J. Food Qual. 2018, 2018, 7949782. [Google Scholar] [CrossRef]
- Boissy, J.; Aubin, J.; Drissi, A.; van der Werf, H.M.G.; Bell, G.J.; Kaushik, S.J. Environmental Impacts of Plant-Based Salmonid Diets at Feed and Farm Scales. Aquaculture 2011, 321, 61–70. [Google Scholar] [CrossRef]
- Troell, M.; Naylor, R.L.; Metian, M.; Beveridge, M.; Tyedmers, P.H.; Folke, C.; Arrow, K.J.; Barrett, S.; Crépin, A.-S.; Ehrlich, P.R.; et al. Does Aquaculture Add Resilience to the Global Food System? Proc. Natl. Acad. Sci. USA 2014, 111, 13257–13263. [Google Scholar] [CrossRef]
- Garcia, M.A.; Altieri, M.A. Transgenic Crops: Implications for Biodiversity and Sustainable Agriculture. Bull. Sci. Technol. Soc. 2005, 25, 335–353. [Google Scholar] [CrossRef]
- Hoc, B.; Tomson, T.; Malumba, P.; Blecker, C.; Jijakli, M.H.; Purcaro, G.; Francis, F.; Caparros Megido, R. Production of Rainbow Trout (Oncorhynchus mykiss) Using Black Soldier Fly (Hermetia Illucens) Prepupae-Based Formulations with Differentiated Fatty Acid Profiles. Sci. Total Environ. 2021, 794, 148647. [Google Scholar] [CrossRef]
- Stamer, A. Insect Proteins—A New Source for Animal Feed: The Use of Insect Larvae to Recycle Food Waste in High-quality Protein for Livestock and Aquaculture Feeds Is Held Back Largely Owing to Regulatory Hurdles. EMBO Rep. 2015, 16, 676–680. [Google Scholar] [CrossRef]
- Boerema, A.; Peeters, A.; Swolfs, S.; Vandevenne, F.; Jacobs, S.; Staes, J.; Meire, P. Soybean Trade: Balancing Environmental and Socio-Economic Impacts of an Intercontinental Market. PLoS ONE 2016, 11, e0155222. [Google Scholar] [CrossRef]
- Mendes dos Reis, J.G.; Sanches Amorim, P.; Sarsfield Pereira Cabral, J.A.; Toloi, R.C. The Impact of Logistics Performance on Argentina, Brazil, and the US Soybean Exports from 2012 to 2018: A Gravity Model Approach. Agriculture 2020, 10, 338. [Google Scholar] [CrossRef]
- Li, P.; Mai, K.; Trushenski, J.; Wu, G. New Developments in Fish Amino Acid Nutrition: Towards Functional and Environmentally Oriented Aquafeeds. Amino Acids 2009, 37, 43–53. [Google Scholar] [CrossRef] [PubMed]
- He, J.-Y.; Tian, L.-X.; Lemme, A.; Gao, W.; Yang, H.-J.; Niu, J.; Liang, G.-Y.; Chen, P.-F.; Liu, Y.-J. Methionine and Lysine Requirements for Maintenance and Efficiency of Utilization for Growth of Two Sizes of Tilapia (Oreochromis niloticus). Aquac. Nutr. 2013, 19, 629–640. [Google Scholar] [CrossRef]
- Sarker, P.K.; Hosokawa, H. Effect of Phytase on Growth and Phosphorus Utilization in Japanese Flounder (Paralichthys olivaceus). Int. J. Recirc. Aquac. 2009, 10, 25–41. [Google Scholar] [CrossRef]
- Sarker, P.K.; Fournier, J.; Boucher, E.; Proulx, E.; de la Noüe, J.; Vandenberg, G.W. Effects of Low Phosphorus Ingredient Combinations on Weight Gain, Apparent Digestibility Coefficients, Non-Fecal Phosphorus Excretion, Phosphorus Retention and Loading of Large Rainbow Trout (Oncorhynchus mykiss). Anim. Feed. Sci. Technol. 2011, 168, 241–249. [Google Scholar] [CrossRef]
- Pelletier, N.; Tyedmers, P. Life Cycle Assessment of Frozen Tilapia Fillets From Indonesian Lake-Based and Pond-Based Intensive Aquaculture Systems. J. Ind. Ecol. 2010, 14, 467–481. [Google Scholar] [CrossRef]
- Wang, F.; Yang, Y.-H.; Han, Z.-Z.; Dong, H.-W.; Yang, C.-H.; Zou, Z.-Y. Effects of Phytase Pretreatment of Soybean Meal and Phytase-Sprayed in Diets on Growth, Apparent Digestibility Coefficient and Nutrient Excretion of Rainbow Trout (Oncorhynchus mykiss Walbaum). Aquac. Int. 2008, 17, 143–157. [Google Scholar] [CrossRef]
- National Research Council (U.S.). Committee on Animal Nutrition. In Nutrient Requirements of Fish; National Academy Press: Washington, DC, USA, 1993; ISBN 978-0-309-04891-0. [Google Scholar]
- Jobling, M. National Research Council (NRC): Nutrient Requirements of Fish and Shrimp. Aquac. Int. 2012, 20, 601–602. [Google Scholar] [CrossRef]
- Kumar, V.; Sinha, A.K.; Makkar, H.P.S.; Boeck, G.D.; Becker, K. Phytate and Phytase in Fish Nutrition. J. Anim. Physiol. Anim. Nutr. 2012, 96, 335–364. [Google Scholar] [CrossRef]
- Storebakken, T.; Shearer, K.D.; Roem, A.J. Availability of Protein, Phosphorus and Other Elements in Fish Meal, Soy-Protein Concentrate and Phytase-Treated Soy-Protein-Concentrate-Based Diets to Atlantic Salmon, Salmo salar. Aquaculture 1998, 161, 365–379. [Google Scholar] [CrossRef]
- Baruah, K.; Sahu, N.P.; Pal, A.K.; Debnath, D. Dietary Phytase: An Ideal Approach for a Cost Effective and Low-Polluting Aquafeed. NAGA WorldFish Cent. Q. 2004, 27, 15–19. [Google Scholar]
- Naylor, R.L.; Hardy, R.W.; Bureau, D.P.; Chiu, A.; Elliott, M.; Farrell, A.P.; Forster, I.; Gatlin, D.M.; Goldburg, R.J.; Hua, K.; et al. Feeding Aquaculture in an Era of Finite Resources. Proc. Natl. Acad. Sci. USA 2009, 106, 15103–15110. [Google Scholar] [CrossRef] [PubMed]
- Sajjadi, M.; Carter, C.G. Dietary Phytase Supplementation and the Utilisation of Phosphorus by Atlantic Salmon (Salmo salar L.) Fed a Canola-Meal-Based Diet. Aquaculture 2004, 240, 417–431. [Google Scholar] [CrossRef]
- Dalsgaard, J.; Ekmann, K.S.; Pedersen, P.B.; Verlhac, V. Effect of Supplemented Fungal Phytase on Performance and Phosphorus Availability by Phosphorus-Depleted Juvenile Rainbow Trout (Oncorhynchus mykiss), and on the Magnitude and Composition of Phosphorus Waste Output. Aquaculture 2009, 286, 105–112. [Google Scholar] [CrossRef]
- Barrows, F.T.; Bellis, D.; Krogdahl, Å.; Silverstein, J.T.; Herman, E.M.; Sealey, W.M.; Rust, M.B.; Gatlin, D.M., III. Report of the Plant Products in Aquafeed Strategic Planning Workshop: An Integrated, Interdisciplinary Research Roadmap for Increasing Utilization of Plant Feedstuffs in Diets for Carnivorous Fish. Rev. Fish. Sci. 2008, 16, 449–455. [Google Scholar] [CrossRef]
- Cho, C.Y.; Bureau, D.P. Reduction of Waste Output from Salmonid Aquaculture through Feeds and Feeding. Progress. Fish-Cult. 1997, 59, 155–160. [Google Scholar] [CrossRef]
- National Research Council; Division on Earth and Life Studies; Board on Agriculture and Natural Resources; Committee on the Nutrient Requirements of Fish and Shrimp. Nutrient Requirements of Fish and Shrimp; National Academies Press: Washington, DC, USA, 2011; ISBN 978-0-309-47322-4. [Google Scholar]
- Drew, M.D.; Borgeson, T.L.; Thiessen, D.L. A Review of Processing of Feed Ingredients to Enhance Diet Digestibility in Finfish. Anim. Feed. Sci. Technol. 2007, 138, 118–136. [Google Scholar] [CrossRef]
- Hixson, S.M.; Parrish, C.C.; Anderson, D.M. Full Substitution of Fish Oil with Camelina (Camelina sativa) Oil, with Partial Substitution of Fish Meal with Camelina Meal, in Diets for Farmed Atlantic Salmon (Salmo salar) and Its Effect on Tissue Lipids and Sensory Quality. Food Chem. 2014, 157, 51–61. [Google Scholar] [CrossRef]
- Torstensen, B.E.; Bell, J.G.; Rosenlund, G.; Henderson, R.J.; Graff, I.E.; Tocher, D.R.; Lie, Ø.; Sargent, J.R. Tailoring of Atlantic Salmon (Salmo salar L.) Flesh Lipid Composition and Sensory Quality by Replacing Fish Oil with a Vegetable Oil Blend. J. Agric. Food Chem. 2005, 53, 10166–10178. [Google Scholar] [CrossRef]
- Turchini, G.M.; Ng, W.-K.; Tocher, D.R.; Ng, W.-K.; Tocher, D.R. Fish Oil Replacement and Alternative Lipid Sources in Aquaculture Feeds; CRC Press: Boca Raton, FL, USA, 2010; ISBN 978-0-429-15103-3. [Google Scholar]
- Bell, J.G.; McEvoy, J.; Tocher, D.R.; McGhee, F.; Campbell, P.J.; Sargent, J.R. Replacement of Fish Oil with Rapeseed Oil in Diets of Atlantic Salmon (Salmo salar) Affects Tissue Lipid Compositions and Hepatocyte Fatty Acid Metabolism. J. Nutr. 2001, 131, 1535–1543. [Google Scholar] [CrossRef]
- Drew, M.D.; Ogunkoya, A.E.; Janz, D.M.; Van Kessel, A.G. Dietary Influence of Replacing Fish Meal and Oil with Canola Protein Concentrate and Vegetable Oils on Growth Performance, Fatty Acid Composition and Organochlorine Residues in Rainbow Trout (Oncorhynchus mykiss). Aquaculture 2007, 267, 260–268. [Google Scholar] [CrossRef]
- Grisdale-Helland, B.; Ruyter, B.; Rosenlund, G.; Obach, A.; Helland, S.J.; Sandberg, M.G.; Standal, H.; Røsjø, C. Influence of High Contents of Dietary Soybean Oil on Growth, Feed Utilization, Tissue Fatty Acid Composition, Heart Histology and Standard Oxygen Consumption of Atlantic Salmon (Salmo salar) Raised at Two Temperatures. Aquaculture 2002, 207, 311–329. [Google Scholar] [CrossRef]
- Bell, J.G.; Waagbø, R. Safe and Nutritious Aquaculture Produce: Benefits and Risks of Alternative Sustainable Aquafeeds. In Aquaculture in the Ecosystem; Holmer, M., Black, K., Duarte, C.M., Marbà, N., Karakassis, I., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 185–225. ISBN 978-1-4020-6810-2. [Google Scholar]
- Turchini, G.M.; Torstensen, B.E.; Ng, W.-K. Fish Oil Replacement in Finfish Nutrition. Rev. Aquac. 2009, 1, 10–57. [Google Scholar] [CrossRef]
- Olsen, Y. Resources for Fish Feed in Future Mariculture. Aquac. Environ. Interact. 2011, 1, 187–200. [Google Scholar] [CrossRef]
- Berntssen, M.H.G.; Maage, A.; Julshamn, K.; Oeye, B.E.; Lundebye, A.-K. Carry-over of Dietary Organochlorine Pesticides, PCDD/Fs, PCBs, and Brominated Flame Retardants to Atlantic Salmon (Salmo salar L.) Fillets. Chemosphere 2011, 83, 95–103. [Google Scholar] [CrossRef]
- Sprague, M.; Dick, J.R.; Tocher, D.R. Impact of Sustainable Feeds on Omega-3 Long-Chain Fatty Acid Levels in Farmed Atlantic Salmon, 2006–2015. Sci. Rep. 2016, 6, 21892. [Google Scholar] [CrossRef]
- Gebauer, S.K.; Psota, T.L.; Harris, W.S.; Kris-Etherton, P.M. N−3 Fatty Acid Dietary Recommendations and Food Sources to Achieve Essentiality and Cardiovascular Benefits. Am. J. Clin. Nutr. 2006, 83, 1526S–1535S. [Google Scholar] [CrossRef]
- Lucas, M.; Asselin, G.; Plourde, M.; Cunnane, S.C.; Dewailly, É.; Dodin, S. N-3 Fatty Acid Intake from Marine Food Products among Quebecers: Comparison to Worldwide Recommendations. Public Health Nutr. 2010, 13, 63–70. [Google Scholar] [CrossRef]
- Nakamura, M.T.; Nara, T.Y. Structure, Function, and Dietary Regulation of Δ6, Δ5, and Δ9 Desaturases. Annu. Rev. Nutr. 2004, 24, 345–376. [Google Scholar] [CrossRef]
- Farrell, A.P.; Friesen, E.N.; Higgs, D.A.; Ikonomou, M.G. Toward Improved Public Confidence in Farmed Fish Quality: A Canadian Perspective on the Consequences of Diet Selection. J. World Aquac. Soc. 2010, 41, 207–224. [Google Scholar] [CrossRef]
- Moya-Falcón, C.; Thomassen, M.S.; Jakobsen, J.V.; Ruyter, B. Effects of Dietary Supplementation of Rapeseed Oil on Metabolism of [1-14C]18∶1n−9, [1-14C]20∶3n−6, and [1-14C]20∶4n−3 in Atlantic Salmon Heaptocytes. Lipids 2005, 40, 709–717. [Google Scholar] [CrossRef]
- Moya-Falcón, C.; Hvattum, E.; Tran, T.N.; Thomassen, M.S.; Skorve, J.; Ruyter, B. Phospholipid Molecular Species, β-Oxidation, Desaturation and Elongation of Fatty Acids in Atlantic Salmon Hepatocytes: Effects of Temperature and 3-Thia Fatty Acids. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2006, 145, 68–80. [Google Scholar] [CrossRef]
- Codabaccus, M.B.; Bridle, A.R.; Nichols, P.D.; Carter, C.G. Effect of Feeding Atlantic Salmon (Salmo salar L.) a Diet Enriched with Stearidonic Acid from Parr to Smolt on Growth and n-3 Long-Chain PUFA Biosynthesis. Br. J. Nutr. 2011, 105, 1772–1782. [Google Scholar] [CrossRef]
- Ng, W.-K.; Low, S.-Y. Evaluation of Spent Bleaching Clay from Palm Oil Refining as an Ingredient for Diets of Red Hybrid Tilapia, Oreochromis Sp. J. Appl. Aquac. 2005, 17, 87–97. [Google Scholar] [CrossRef]
- Ng, W.-K.; Lim, P.-K.; Sidek, H. The Influence of a Dietary Lipid Source on Growth, Muscle Fatty Acid Composition and Erythrocyte Osmotic Fragility of Hybrid Tilapia. Fish Physiol. Biochem. 2001, 25, 301–310. [Google Scholar] [CrossRef]
- Karapanagiotidis, I.T.; Bell, M.V.; Little, D.C.; Yakupitiyage, A. Replacement of Dietary Fish Oils by Alpha-Linolenic Acid-Rich Oils Lowers Omega 3 Content in Tilapia Flesh. Lipids 2007, 42, 547–559. [Google Scholar] [CrossRef]
- Weaver, K.L.; Ivester, P.; Chilton, J.A.; Wilson, M.D.; Pandey, P.; Chilton, F.H. The Content of Favorable and Unfavorable Polyunsaturated Fatty Acids Found in Commonly Eaten Fish. J. Am. Diet. Assoc. 2008, 108, 1178–1185. [Google Scholar] [CrossRef]
- Rosenthal, E. Another Side of Tilapia, the Perfect Factory Fish. The New York Times, 3 May 2011. [Google Scholar]
- Karapanagiotidis, I.T.; Bell, M.V.; Little, D.C.; Yakupitiyage, A.; Rakshit, S.K. Polyunsaturated Fatty Acid Content of Wild and Farmed Tilapias in Thailand: Effect of Aquaculture Practices and Implications for Human Nutrition. J. Agric. Food Chem. 2006, 54, 4304–4310. [Google Scholar] [CrossRef]
- Teoh, C.-Y.; Turchini, G.M.; Ng, W.-K. Erratum to “Genetically Improved Farmed Nile Tilapia and Red Hybrid Tilapia Showed Differences in Fatty Acid Metabolism When Fed Diets with Added Fish Oil or a Vegetable Oil Blend”. Aquaculture 2011, 316, 144–154. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Evolutionary Aspects of Omega-3 Fatty Acids in the Food Supply. Prostaglandins Leukot. Essent. Fat. Acids 1999, 60, 421–429. [Google Scholar] [CrossRef]
- Burghardt, P.R.; Kemmerer, E.S.; Buck, B.J.; Osetek, A.J.; Yan, C.; Koch, L.G.; Britton, S.L.; Evans, S.J. Dietary N-3:N-6 Fatty Acid Ratios Differentially Influence Hormonal Signature in a Rodent Model of Metabolic Syndrome Relative to Healthy Controls. Nutr. Metab. 2010, 7, 53. [Google Scholar] [CrossRef]
- Strobel, C.; Jahreis, G.; Kuhnt, K. Survey of N-3 and n-6 Polyunsaturated Fatty Acids in Fish and Fish Products. Lipids Health Dis. 2012, 11, 144. [Google Scholar] [CrossRef] [PubMed]
- Justi, K.C.; Hayashi, C.; Visentainer, J.V.; de Souza, N.E.; Matsushita, M. The Influence of Feed Supply Time on the Fatty Acid Profile of Nile Tilapia (Oreochromis Niloticus) Fed on a Diet Enriched with n-3 Fatty Acids. Food Chem. 2003, 80, 489–493. [Google Scholar] [CrossRef]
- Young, K. Omega-6 (n-6) and Omega-3 (n-3) Fatty Acids in Tilapia and Human Health: A Review. Int. J. Food Sci. Nutr. 2009, 60, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Sarker, P.K.; Kapuscinski, A.R.; Lanois, A.J.; Livesey, E.D.; Bernhard, K.P.; Coley, M.L. Towards Sustainable Aquafeeds: Complete Substitution of Fish Oil with Marine Microalga Schizochytrium Sp. Improves Growth and Fatty Acid Deposition in Juvenile Nile Tilapia (Oreochromis niloticus). PLoS ONE 2016, 11, e0156684. [Google Scholar] [CrossRef] [PubMed]
- Beal, C.M.; Gerber, L.N.; Thongrod, S.; Phromkunthong, W.; Kiron, V.; Granados, J.; Archibald, I.; Greene, C.H.; Huntley, M.E. Marine Microalgae Commercial Production Improves Sustainability of Global Fisheries and Aquaculture. Sci. Rep. 2018, 8, 15064. [Google Scholar] [CrossRef]
- Shah, M.R.; Lutzu, G.A.; Alam, A.; Sarker, P.; Kabir Chowdhury, M.A.; Parsaeimehr, A.; Liang, Y.; Daroch, M. Microalgae in Aquafeeds for a Sustainable Aquaculture Industry. J. Appl. Phycol. 2018, 30, 197–213. [Google Scholar] [CrossRef]
- Sarker, P.K.; Kapuscinski, A.R.; Vandenberg, G.W.; Proulx, E.; Sitek, A.J. Towards Sustainable and Ocean-Friendly Aquafeeds: Evaluating a Fish-Free Feed for Rainbow Trout (Oncorhynchus mykiss) Using Three Marine Microalgae Species. Elem. Sci. Anthr. 2020, 8, 5. [Google Scholar] [CrossRef]
- Kiron, V.; Sørensen, M.; Huntley, M.; Vasanth, G.K.; Gong, Y.; Dahle, D.; Palihawadana, A.M. Defatted Biomass of the Microalga, Desmodesmus Sp., Can Replace Fishmeal in the Feeds for Atlantic Salmon. Front. Mar. Sci. 2016, 3, 67. [Google Scholar] [CrossRef]
- Sirakov, I.; Velichkova, K.; Nikolov, G. The Effect of Algae Meal (Spirulina) on the Growth Performance and Carcass Parameters of Rainbow Trout (Oncorhynchus mykiss). J. BioSci. Biotechnol. 2012, 151–156. [Google Scholar]
- Sarker, P.K.; Kapuscinski, A.R.; Bae, A.Y.; Donaldson, E.; Sitek, A.J.; Fitzgerald, D.S.; Edelson, O.F. Towards Sustainable Aquafeeds: Evaluating Substitution of Fishmeal with Lipid-Extracted Microalgal Co-Product (Nannochloropsis oculata) in Diets of Juvenile Nile Tilapia (Oreochromis niloticus). PLoS ONE 2018, 13, e0201315. [Google Scholar] [CrossRef]
- Sørensen, M.; Gong, Y.; Bjarnason, F.; Vasanth, G.K.; Dahle, D.; Huntley, M.; Kiron, V. Nannochloropsis Oceania-Derived Defatted Meal as an Alternative to Fishmeal in Atlantic Salmon Feeds. PLoS ONE 2017, 12, e0179907. [Google Scholar] [CrossRef]
- Sprague, M.; Walton, J.; Campbell, P.J.; Strachan, F.; Dick, J.R.; Bell, J.G. Replacement of Fish Oil with a DHA-Rich Algal Meal Derived from Schizochytrium Sp. on the Fatty Acid and Persistent Organic Pollutant Levels in Diets and Flesh of Atlantic Salmon (Salmo salar, L.) Post-Smolts. Food Chem. 2015, 185, 413–421. [Google Scholar] [CrossRef]
- Wang, Y.; Li, M.; Filer, K.; Xue, Y.; Ai, Q.; Mai, K. Replacement of Fish Oil with a DHA-Rich Schizochytrium Meal on Growth Performance, Activities of Digestive Enzyme and Fatty Acid Profile of Pacific White Shrimp (Litopenaeus vannamei) Larvae. Aquac. Nutr. 2017, 23, 1113–1120. [Google Scholar] [CrossRef]
- Kousoulaki, K.; Mørkøre, T.; Nengas, I.; Berge, R.K.; Sweetman, J. Microalgae and Organic Minerals Enhance Lipid Retention Efficiency and Fillet Quality in Atlantic Salmon (Salmo salar L.). Aquaculture 2016, 451, 47–57. [Google Scholar] [CrossRef]
- Tibaldi, E.; Chini Zittelli, G.; Parisi, G.; Bruno, M.; Giorgi, G.; Tulli, F.; Venturini, S.; Tredici, M.R.; Poli, B.M. Growth Performance and Quality Traits of European Sea Bass (D. Labrax) Fed Diets Including Increasing Levels of Freeze-Dried Isochrysis Sp. (T-ISO) Biomass as a Source of Protein and n-3 Long Chain PUFA in Partial Substitution of Fish Derivatives. Aquaculture 2015, 440, 60–68. [Google Scholar] [CrossRef]
- Tocher, D.R.; Betancor, M.B.; Sprague, M.; Olsen, R.E.; Napier, J.A. Omega-3 Long-Chain Polyunsaturated Fatty Acids, EPA and DHA: Bridging the Gap between Supply and Demand. Nutrients 2019, 11, 89. [Google Scholar] [CrossRef]
- Øverland, M.; Karlsson, A.; Mydland, L.T.; Romarheim, O.H.; Skrede, A. Evaluation of Candida Utilis, Kluyveromyces Marxianus and Saccharomyces Cerevisiae Yeasts as Protein Sources in Diets for Atlantic Salmon (Salmo salar). Aquaculture 2013, 402–403, 1–7. [Google Scholar] [CrossRef]
- Glencross, B.D.; Huyben, D.; Schrama, J.W. The Application of Single-Cell Ingredients in Aquaculture Feeds—A Review. Fishes 2020, 5, 22. [Google Scholar] [CrossRef]
- Øvrum Hansen, J.; Hofossæter, M.; Sahlmann, C.; Ånestad, R.; Reveco-Urzua, F.E.; Press, C.M.; Mydland, L.T.; Øverland, M. Effect of Candida Utilis on Growth and Intestinal Health of Atlantic Salmon (Salmo salar) Parr. Aquaculture 2019, 511, 734239. [Google Scholar] [CrossRef]
- Ritala, A.; Häkkinen, S.T.; Toivari, M.; Wiebe, M.G. Single Cell Protein-State-of-the-Art, Industrial Landscape and Patents 2001-2016. Front. Microbiol. 2017, 8, 2009. [Google Scholar] [CrossRef]
- Suman, G.; Nupur, M.; Anuradha, S.; Pradeep, B. Single Cell Protein Production: A Review. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 251–262. [Google Scholar]
- Aas, T.S.; Grisdale-Helland, B.; Terjesen, B.F.; Helland, S.J. Improved Growth and Nutrient Utilisation in Atlantic Salmon (Salmo salar) Fed Diets Containing a Bacterial Protein Meal. Aquaculture 2006, 259, 365–376. [Google Scholar] [CrossRef]
- Tlusty, M.; Rhyne, A.; Szczebak, J.T.; Bourque, B.; Bowen, J.L.; Burr, G.; Marx, C.J.; Feinberg, L. A Transdisciplinary Approach to the Initial Validation of a Single Cell Protein as an Alternative Protein Source for Use in Aquafeeds. PeerJ 2017, 5, e3170. [Google Scholar] [CrossRef] [PubMed]
- Chumpol, S.; Kantachote, D.; Nitoda, T.; Kanzaki, H. Administration of Purple Nonsulfur Bacteria as Single Cell Protein by Mixing with Shrimp Feed to Enhance Growth, Immune Response and Survival in White Shrimp (Litopenaeus vannamei) Cultivation. Aquaculture 2018, 489, 85–95. [Google Scholar] [CrossRef]
- Hamidoghli, A.; Yun, H.; Won, S.; Kim, S.; Farris, N.W.; Bai, S.C. Evaluation of a Single-Cell Protein as a Dietary Fish Meal Substitute for Whiteleg Shrimp Litopenaeus vannamei. Fish Sci. 2019, 85, 147–155. [Google Scholar] [CrossRef]
- Glencross, B.; Irvin, S.; Arnold, S.; Blyth, D.; Bourne, N.; Preston, N. Effective Use of Microbial Biomass Products to Facilitate the Complete Replacement of Fishery Resources in Diets for the Black Tiger Shrimp, Penaeus Monodon. Aquaculture 2014, 431, 12–19. [Google Scholar] [CrossRef]
- Arnold, S.; Smullen, R.; Briggs, M.; West, M.; Glencross, B. The Combined Effect of Feed Frequency and Ration Size of Diets with and without Microbial Biomass on the Growth and Feed Conversion of Juvenile Penaeus Monodon. Aquac. Nutr. 2016, 22, 1340–1347. [Google Scholar] [CrossRef]
- Sharma, P.; Gaur, V.K.; Kim, S.-H.; Pandey, A. Microbial Strategies for Bio-Transforming Food Waste into Resources. Bioresour. Technol. 2020, 299, 122580. [Google Scholar] [CrossRef]
- Olmos Soto, J.; Paniagua-Michel, J.D.J.; Lopez, L.; Ochoa, L. Functional Feeds in Aquaculture. In Springer Handbook of Marine Biotechnology; Kim, S.-K., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 1303–1319. ISBN 978-3-642-53971-8. [Google Scholar]
- Ruiz-Lopez, N.; Haslam, R.P.; Napier, J.A.; Sayanova, O. Successful High-Level Accumulation of Fish Oil Omega-3 Long-Chain Polyunsaturated Fatty Acids in a Transgenic Oilseed Crop. Plant J. 2014, 77, 198–208. [Google Scholar] [CrossRef]
- Thomassen, M.S.; Rein, D.; Berge, G.M.; Østbye, T.-K.; Ruyter, B. High Dietary EPA Does Not Inhibit ∆5 and ∆6 Desaturases in Atlantic Salmon (Salmo salar L.) Fed Rapeseed Oil Diets. Aquaculture 2012, 360–361, 78–85. [Google Scholar] [CrossRef]
- Lock, E.-J.; Biancarosa, I.; Gasco, L. Insects as Raw Materials in Compound Feed for Aquaculture. In Edible Insects in Sustainable Food Systems; Halloran, A., Flore, R., Vantomme, P., Roos, N., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 263–276. ISBN 978-3-319-74011-9. [Google Scholar]
- Lock, E.R.; Arsiwalla, T.; Waagbø, R. Insect Larvae Meal as an Alternative Source of Nutrients in the Diet of Atlantic Salmon (Salmo salar) Postsmolt. Aquac. Nutr. 2016, 22, 1202–1213. [Google Scholar] [CrossRef]
- Biancarosa, I.; Sele, V.; Belghit, I.; Ørnsrud, R.; Lock, E.-J.; Amlund, H. Replacing Fish Meal with Insect Meal in the Diet of Atlantic Salmon (Salmo salar) Does Not Impact the Amount of Contaminants in the Feed and It Lowers Accumulation of Arsenic in the Fillet. Food Addit. Contam. Part A 2019, 36, 1191–1205. [Google Scholar] [CrossRef]
- Belghit, I.; Liland, N.S.; Waagbø, R.; Biancarosa, I.; Pelusio, N.; Li, Y.; Krogdahl, Å.; Lock, E.-J. Potential of In-sect-Based Diets for Atlantic Salmon (Salmo salar). Aquaculture 2018, 491, 72–81. [Google Scholar] [CrossRef]
- Belghit, I.; Liland, N.S.; Gjesdal, P.; Biancarosa, I.; Menchetti, E.; Li, Y.; Waagbø, R.; Krogdahl, Å.; Lock, E.-J. Black Soldier Fly Larvae Meal Can Replace Fish Meal in Diets of Sea-Water Phase Atlantic Salmon (Salmo salar). Aquaculture 2019, 503, 609–619. [Google Scholar] [CrossRef]
- Salomone, R.; Saija, G.; Mondello, G.; Giannetto, A.; Fasulo, S.; Savastano, D. Environmental Impact of Food Waste Bioconversion by Insects: Application of Life Cycle Assessment to Process Using Hermetia Illucens. J. Clean. Prod. 2017, 140, 890–905. [Google Scholar] [CrossRef]
- Renna, M.; Schiavone, A.; Gai, F.; Dabbou, S.; Lussiana, C.; Malfatto, V.; Prearo, M.; Capucchio, M.T.; Biasato, I.; Biasibetti, E.; et al. Evaluation of the Suitability of a Partially Defatted Black Soldier Fly (Hermetia Illucens L.) Larvae Meal as Ingredient for Rainbow Trout (Oncorhynchus mykiss Walbaum) Diets. J. Anim. Sci. Biotechnol. 2017, 8, 57. [Google Scholar] [CrossRef]
- Jackson and Newton Project to Model the Use of Fisheries By-Products in The Production of Marine Ingredients, with Special Reference to the Omega 3 Fatty Acids Epa and Dha. Available online: https://www.iffo.com/system/files/downloads/Report%20IoA%20IFFO%20project%20Final_0.pdf (accessed on 25 September 2020).
- Henriksson, P.J.G.; Belton, B.; Jahan, K.M.; Rico, A. Measuring the Potential for Sustainable Intensification of Aquaculture in Bangladesh Using Life Cycle Assessment. Proc. Natl. Acad. Sci. USA 2018, 115, 2958–2963. [Google Scholar] [CrossRef]
- Campos, I.; Pinheiro Valente, L.M.; Matos, E.; Marques, P.; Freire, F. Life-Cycle Assessment of Animal Feed Ingredients: Poultry Fat, Poultry by-Product Meal and Hydrolyzed Feather Meal. J. Clean. Prod. 2020, 252, 119845. [Google Scholar] [CrossRef]
- Sandhu, H.; Scialabba, N.E.-H.; Warner, C.; Behzadnejad, F.; Keohane, K.; Houston, R.; Fujiwara, D. Evaluating the Holistic Costs and Benefits of Corn Production Systems in Minnesota, US. Sci. Rep. 2020, 10, 3922. [Google Scholar] [CrossRef]
- Sillman, J.; Nygren, L.; Kahiluoto, H.; Ruuskanen, V.; Tamminen, A.; Bajamundi, C.; Nappa, M.; Wuokko, M.; Lindh, T.; Vainikka, P.; et al. Bacterial Protein for Food and Feed Generated via Renewable Energy and Direct Air Capture of CO2: Can It Reduce Land and Water Use? Glob. Food Secur. 2019, 22, 25–32. [Google Scholar] [CrossRef]
- Ketzer, F.; Skarka, J.; Rösch, C. Critical Review of Microalgae LCA Studies for Bioenergy Production. Bioenergy Res. 2018, 11, 95–105. [Google Scholar] [CrossRef]
- Tibbetts, S.M.; Yasumaru, F.; Lemos, D. In Vitro Prediction of Digestible Protein Content of Marine Microalgae (Nannochloropsis granulata) Meals for Pacific White Shrimp (Litopenaeus vannamei) and Rainbow Trout (Oncorhynchus mykiss). Algal Res. 2017, 21, 76–80. [Google Scholar] [CrossRef]
- Teuling, E.; Wierenga, P.A.; Agboola, J.O.; Gruppen, H.; Schrama, J.W. Cell Wall Disruption Increases Bioavailability of Nannochloropsis Gaditana Nutrients for Juvenile Nile Tilapia (Oreochromis niloticus). Aquaculture 2019, 499, 269–282. [Google Scholar] [CrossRef]
- Lee, A.K.; Lewis, D.M.; Ashman, P.J. Disruption of Microalgal Cells for the Extraction of Lipids for Biofuels: Processes and Specific Energy Requirements. Biomass Bioenergy 2012, 46, 89–101. [Google Scholar] [CrossRef]
- Safi, C.; Charton, M.; Ursu, A.V.; Laroche, C.; Zebib, B.; Pontalier, P.-Y.; Vaca-Garcia, C. Release of Hydro-Soluble Microalgal Proteins Using Mechanical and Chemical Treatments. Algal Res. 2014, 3, 55–60. [Google Scholar] [CrossRef]
- Avilés-Gaxiola, S.; Chuck-Hernández, C.; Saldívar, S.O.S. Inactivation Methods of Trypsin Inhibitor in Legumes: A Review. J. Food Sci. 2018, 83, 17–29. [Google Scholar] [CrossRef]
- Draganovic, V.; van der Goot, A.J.; Boom, R.; Jonkers, J. Assessment of the Effects of Fish Meal, Wheat Gluten, Soy Protein Concentrate and Feed Moisture on Extruder System Parameters and the Technical Quality of Fish Feed. Anim. Feed. Sci. Technol. 2011, 165, 238–250. [Google Scholar] [CrossRef]
- Barrows, F.T.; Frost, J.B. Evaluation of the Nutritional Quality of Co-Products from the Nut Industry, Algae and an Invertebrate Meal for Rainbow Trout, Oncorhynchus mykiss. Aquaculture 2014, 434, 315–324. [Google Scholar] [CrossRef]
- Simon, C.J.; Blyth, D.; Ahmad Fatan, N.; Suri, S. Microbial Biomass (NovacqTM) Stimulates Feeding and Improves the Growth Performance on Extruded Low to Zero-Fishmeal Diets in Tilapia (GIFT Strain). Aquaculture 2019, 501, 319–324. [Google Scholar] [CrossRef]
- Cowieson, A.J.; Ravindran, V. Effect of Exogenous Enzymes in Maize-Based Diets Varying in Nutrient Density for Young Broilers: Growth Performance and Digestibility of Energy, Minerals and Amino Acids. Br. Poult. Sci. 2008, 49, 37–44. [Google Scholar] [CrossRef]
- Sinha, A.K.; Kumar, V.; Makkar, H.P.S.; De Boeck, G.; Becker, K. Non-Starch Polysaccharides and Their Role in Fish Nutrition—A Review. Food Chem. 2011, 127, 1409–1426. [Google Scholar] [CrossRef]
- Dalsgaard, J.; Verlhac, V.; Hjermitslev, N.H.; Ekmann, K.S.; Fischer, M.; Klausen, M.; Pedersen, P.B. Effects of Exogenous Enzymes on Apparent Nutrient Digestibility in Rainbow Trout (Oncorhynchus mykiss) Fed Diets with High Inclusion of Plant-Based Protein. Anim. Feed. Sci. Technol. 2012, 171, 181–191. [Google Scholar] [CrossRef]
- Dalsgaard, J.; Knudsen, K.E.B.; Verlhac, V.; Ekmann, K.S.; Pedersen, P.B. Supplementing Enzymes to Extruded, Soybean-Based Diet Improves Breakdown of Non-Starch Polysaccharides in Rainbow Trout (Oncorhynchus mykiss). Aquac. Nutr. 2016, 22, 419–426. [Google Scholar] [CrossRef]
- Qi, G.; Ai, Q.; Mai, K.; Xu, W.; Liufu, Z.; Yun, B.; Zhou, H. Effects of Dietary Taurine Supplementation to a Casein-Based Diet on Growth Performance and Taurine Distribution in Two Sizes of Juvenile Turbot (Scophthalmus maximus L.). Aquaculture 2012, 358–359, 122–128. [Google Scholar] [CrossRef]
- Watson, A.M.; Barrows, F.T.; Place, A.R. Effects of Graded Taurine Levels on Juvenile Cobia. N. Am. J. Aquac. 2014, 76, 190–200. [Google Scholar] [CrossRef]
- El-Sayed, A.-F.M. Is Dietary Taurine Supplementation Beneficial for Farmed Fish and Shrimp? A Comprehensive Review. Rev. Aquac. 2014, 6, 241–255. [Google Scholar] [CrossRef]
- Poston, H.A. Effect of Body Size on Growth, Survival, and Chemical Composition of Atlantic Salmon Fed Soy Lecithin and Choline. Progress. Fish-Cult. 1990, 52, 226–230. [Google Scholar] [CrossRef]
- Poston, H.A. Response of Rainbow Trout to Soy Lecithin, Choline, and Autoclaved Isolated Soy Protein. Progress. Fish-Cult. 1991, 53, 85–90. [Google Scholar] [CrossRef]
- Trushenski, J.T.; Kanczuzewski, K.L. Traditional and Modified Soy Oils as Substitutes for Fish Oil in Feeds for Hybrid Striped Bass. N. Am. J. Aquac. 2013, 75, 295–304. [Google Scholar] [CrossRef]
- Engle, C. The Value of Alternative Feed Ingredients. J. World Aquac. Soc. 2017, 48, 377–380. [Google Scholar] [CrossRef]
- Lopes da Silva, T.; Moniz, P.; Silva, C.; Reis, A. The Dark Side of Microalgae Biotechnology: A Heterotrophic Biorefinery Platform Directed to ω-3 Rich Lipid Production. Microorganisms 2019, 7, 670. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarker, P.K. Microorganisms in Fish Feeds, Technological Innovations, and Key Strategies for Sustainable Aquaculture. Microorganisms 2023, 11, 439. https://doi.org/10.3390/microorganisms11020439
Sarker PK. Microorganisms in Fish Feeds, Technological Innovations, and Key Strategies for Sustainable Aquaculture. Microorganisms. 2023; 11(2):439. https://doi.org/10.3390/microorganisms11020439
Chicago/Turabian StyleSarker, Pallab K. 2023. "Microorganisms in Fish Feeds, Technological Innovations, and Key Strategies for Sustainable Aquaculture" Microorganisms 11, no. 2: 439. https://doi.org/10.3390/microorganisms11020439
APA StyleSarker, P. K. (2023). Microorganisms in Fish Feeds, Technological Innovations, and Key Strategies for Sustainable Aquaculture. Microorganisms, 11(2), 439. https://doi.org/10.3390/microorganisms11020439