
Effect of Tea Saponins on Rumen Microbiota and Rumen Function in Qinchuan Beef Cattle

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals, Experimental Design and Diets
2.3. Sample Collection and Processing
2.4. DNA Extraction and Illumina Sequencing
2.5. Bioinformatics Analysis
2.6. Statistical Analysis
2.7. Data Accession Number
3. Results
3.1. Impact of Tea Saponins Addition on the Diversity of Rumen Fermentation Parameters
3.2. Impact of Tea Saponins Addition on the Diversity of Cattle Ruminal Microbiota Diversity Analysis
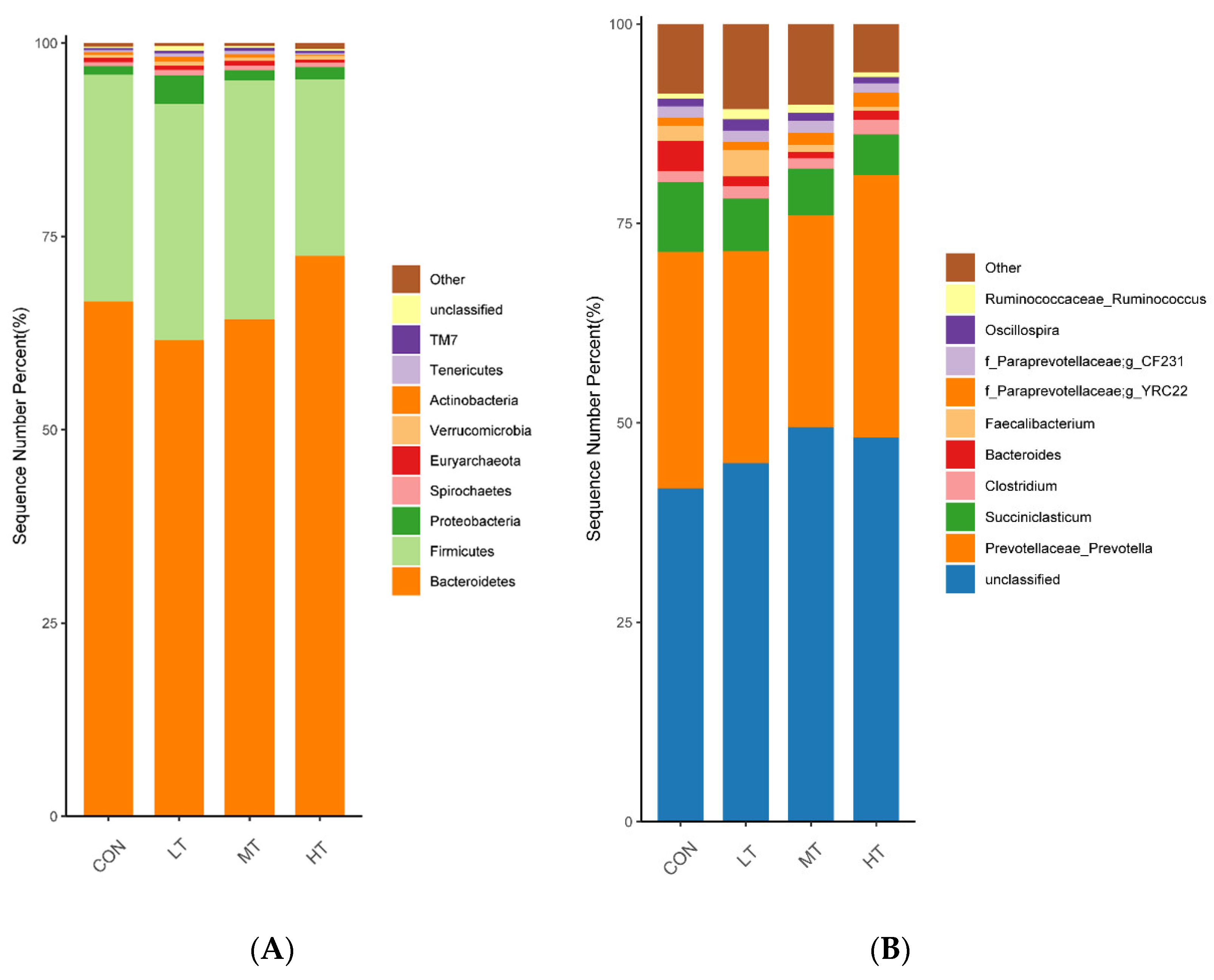
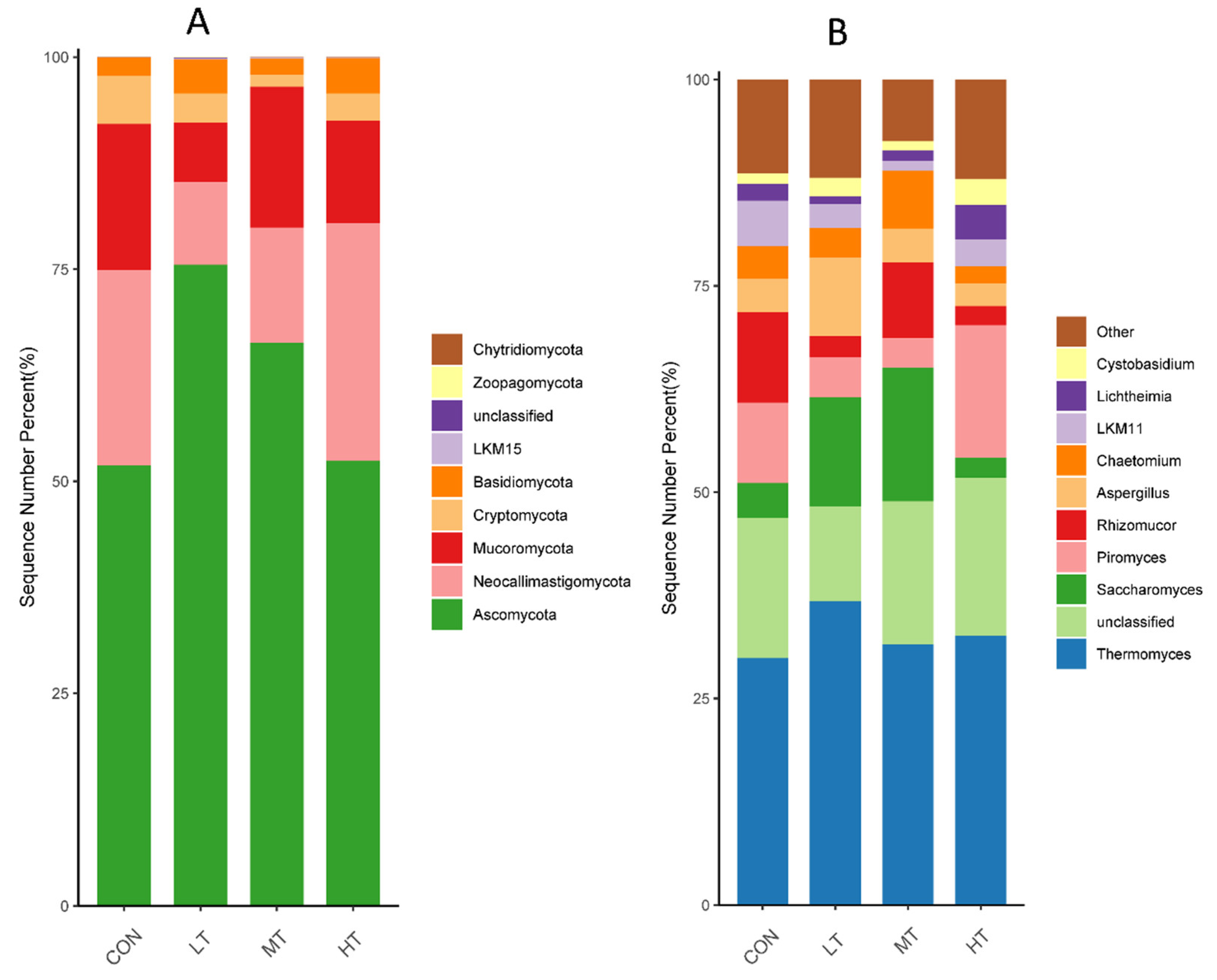
3.3. Analysis of Microbial Composition and Community Structure
3.4. Microbial Community Differences between Four Groups
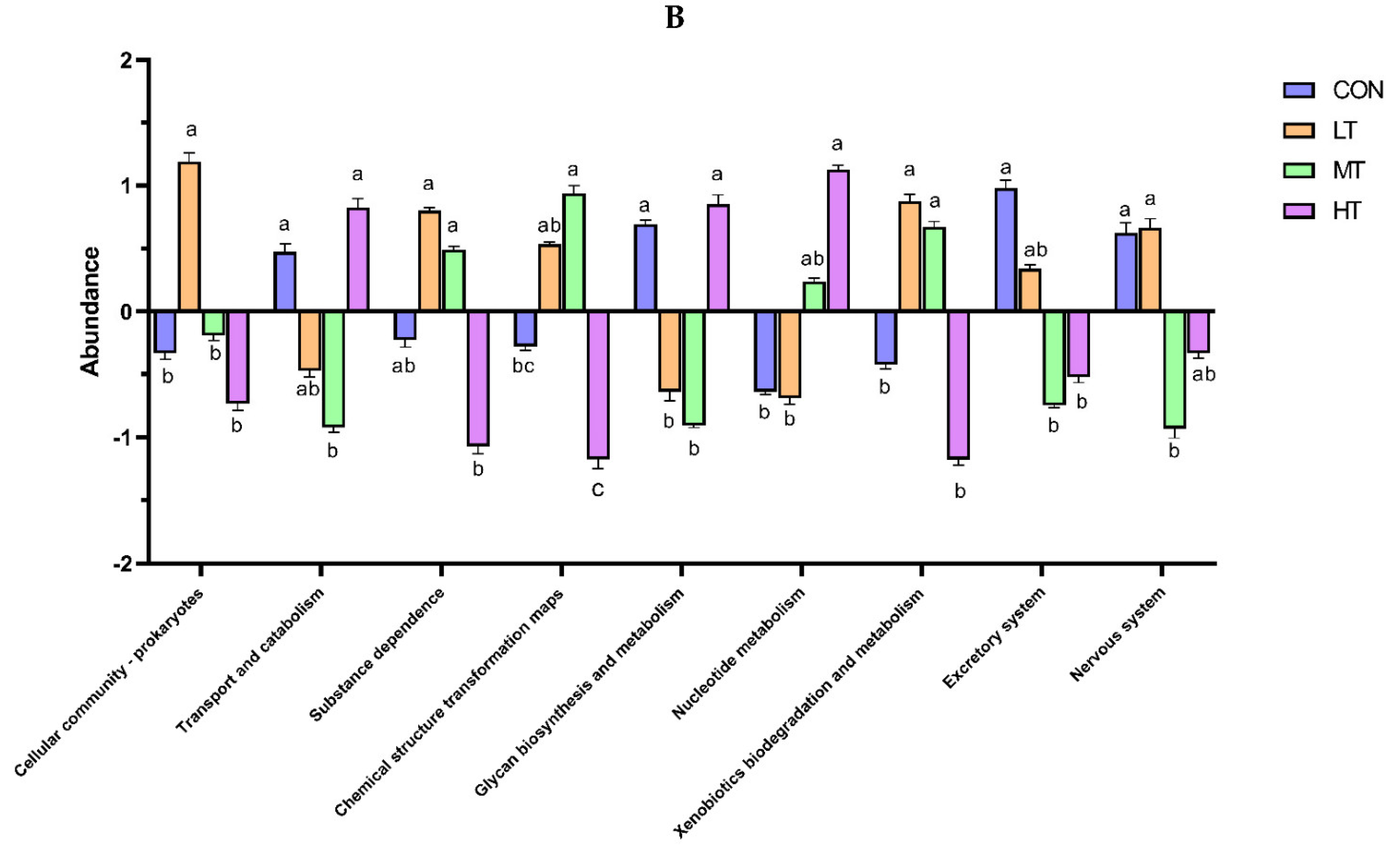
3.5. Functions Estimation
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mingmongkolchai, S.; Panbangred, W. BacillusBacillus probiotics: An alternative to antibiotics for livestock production. J. Appl. Microbiol. 2018, 124, 1334–1346. [Google Scholar] [CrossRef] [PubMed]
- Kuralkar, P.S.; Kuralkar, S.V. Role of Herbal products in Animal Production-An updated review. J. Ethnopharmacol. 2021, 278, 114246. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.K.; Saxena, J. A new perspective on the use of plant secondary metabolites to inhibit methanogenesis in the rumen. Phytochemistry 2010, 71, 1198–1222. [Google Scholar] [CrossRef]
- Wei, M.; Ren, L.; Zhou, Z.; Meng, Q. Effect of Addition of Three Plant Extracts on Gas Production, Ruminal Fermentation, Methane Production and Ruminal Digestibility Based on an in vitro Technique. J. Anim. Vet. Adv. 2012, 64, 1820–1823. [Google Scholar]
- Kozłowska, M.; Cieślak, A.; Jóźwik, A.; El-Sherbiny, M.; Stochmal, A.; Oleszek, W.; Kowalczyk, M.; Filipiak, W.; Szumacher-Strabel, M. The effect of total and individual alfalfa saponins on rumen methane production. J. Sci. Food Agric. 2019, 100, 1922–1930. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Zong, J.; Sun, Y.; Zhang, L.; Ho, C.-T.; Wan, X.; Hou, R. Triterpenoid saponins from the genus Camellia: Structures, biological activities, and molecular simulation for structure–activity relationship. Food Funct. 2018, 9, 3069–3091. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ma, T.; Chen, D.; Zhang, N.; Si, B.; Deng, K.; Tu, Y.; Diao, Q. Effects of tea saponin supplementation on nutrient digestibility, methanogenesis, and ruminal microbial flora in dorper crossbred ewe. Animals 2019, 9, 29. [Google Scholar] [CrossRef]
- Jadhav, R.V.; Kannan, A.; Bhar, R.; Sharma, O.P.; Gulati, A.; Rajkumar, K.; Mal, G.; Singh, B.; Verma, M.R. Effect of tea (Camellia sinensis) seed saponins on in vitro rumen fermentation, methane production and true digestibility at different forage to concentrate ratios. Taylor Fr. 2018, 46, 118–124. [Google Scholar] [CrossRef]
- Weimer, P.J. Redundancy, resilience, and host specificity of the ruminal microbiota: Implications for engineering improved ruminal fermentations. Front. Microbiol. 2015, 6, 296. [Google Scholar] [CrossRef]
- Wang, J.K.; Ye, J.A.; Liu, J.X. Effects of tea saponins on rumen microbiota, rumen fermentation, methane production and growth performance--a review. Trop. Anim. Health Prod. 2012, 44, 697–706. [Google Scholar] [CrossRef]
- Guo, Y.Q.; Liu, J.; Lu, Y.; Zhu, W.Y.; Denman, S.E.; Mcsweeney, C.S. Effect of Tea Saponin on Methanogenesis, Microbial Community Structure and Expression of McrA Gene, in Cultures of Rumen Micro-Organisms. Lett. Appl. Microbiol. 2010, 47, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; He, Z.; Beauchemin, K.A.; Tang, S.; Zhou, C.; Han, X.; Wang, M.; Kang, J.; Odongo, N.E.; Tan, Z. Comparison of two live B acillus species as feed additives for improving in vitro fermentation of cereal straws. Anim. Sci. J. 2016, 87, 27–36. [Google Scholar] [CrossRef]
- Wang, Y.; Qian, P.Y. Conservative fragments in bacterial 16S rRNA genes and primer design for 16S ribosomal DNA amplicons in metagenomic studies. PLoS ONE 2009, 4, e7401. [Google Scholar] [CrossRef] [PubMed]
- Cheung, M.K.; Au, C.H.; Chu, K.H.; Kwan, H.S.; Wong, C.K. Composition and genetic diversity of picoeukaryotes in subtropical coastal waters as revealed by 454 pyrosequencing. ISME J. 2010, 4, 1053–1059. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- Rohart, F.; Gautier, B.; Singh, A.; Lê Cao, K.-A. mixOmics: An R package for ’omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Shah, A.M.; Ma, J.; Wang, Z.; Zou, H.; Hu, R.; Peng, Q. Betaine supplementation improves the production performance, rumen fermentation, and antioxidant profile of dairy cows in heat stress. Animals 2020, 10, 634. [Google Scholar] [CrossRef]
- Poungchompu, O.; Wanapat, M.; Wachirapakorn, C.; Wanapat, S.; Cherdthong, A. Manipulation of ruminal fermentation and methane production by dietary saponins and tannins from mangosteen peel and soapberry fruit. Arch. Anim. Nutr. 2009, 63, 389–400. [Google Scholar] [CrossRef]
- Zhao, C.; Wang, L.; Ke, S.; Chen, X.; Kenéz, .; Xu, W.; Wang, D.; Zhang, F.; Li, Y.; Cui, Z.; et al. Yak rumen microbiome elevates fiber degradation ability and alters rumen fermentation pattern to increase feed efficiency. Anim. Nutr. 2022, 11, 201–214. [Google Scholar] [CrossRef] [PubMed]
- Ben Shabat, S.K.; Sasson, G.; Doron-Faigenboim, A.; Durman, T.; Yaacoby, S.; Berg Miller, M.E.; White, B.A.; Shterzer, N.; Mizrahi, I. Specific microbiome-dependent mechanisms underlie the energy harvest efficiency of ruminants. ISME J. 2016, 10, 2958–2972. [Google Scholar] [CrossRef] [PubMed]
- Abecia, L.; Waddams, K.E.; Martínez-Fernandez, G.; Martín-García, A.I.; Ramos-Morales, E.; Newbold, C.J.; Yáñez-Ruiz, D.R. An antimethanogenic nutritional intervention in early life of ruminants modifies ruminal colonization by Archaea. Archaea 2014, 2014, 1–12. [Google Scholar] [CrossRef]
- Liu, K.; Zhang, Y.; Yu, Z.; Xu, Q.; Zheng, N.; Zhao, S.; Huang, G.; Wang, J. Ruminal microbiota–host interaction and its effect on nutrient metabolism. Anim. Nutr. 2021, 7, 49–55. [Google Scholar] [CrossRef]
- Singh, K.M.; Ahir, V.B.; Tripathi, A.; Ramani, U.V.; Sajnani, M.; Koringa, P.G.; Jakhesara, S.; Pandya, P.R.; Rank, D.N.; Murty, D.S.; et al. Metagenomic analysis of Surti buffalo (Bubalus bubalis) rumen: A preliminary study. Mol. Biol. Rep. 2011, 39, 4841–4848. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, M.N.V.; Jewell, K.A.; Freitas, F.S.; Benjamin, L.A.; Tótola, M.R.; Borges, A.C.; Moraes, C.A.; Suen, G. Characterizing the microbiota across the gastrointestinal tract of a Brazilian Nelore steer. Vet. Microbiol. 2013, 164, 307–314. [Google Scholar] [CrossRef]
- Shen, H.; Chen, Z.; Shen, Z.; Lu, Z. Maintaining stability of the rumen ecosystem is associated with changes of microbial composition and epithelial TLR signaling. Microbiologyopen 2017, 6, e00436. [Google Scholar] [CrossRef]
- Wu, S.; Cui, Z.; Chen, X.; Zheng, L.; Ren, H.; Wang, D.; Yao, J. Diet-ruminal microbiome-host crosstalk contributes to differential effects of calf starter and alfalfa hay on rumen epithelial development and pancreatic α-amylase activity in yak calves. J. Dairy Sci. 2021, 104, 4326–4340. [Google Scholar] [CrossRef]
- Söllinger, A.; Tveit, A.T.; Poulsen, M.; Noel, S.J.; Bengtsson, M.; Bernhardt, J.; Hellwing, A.L.F.; Lund, P.; Riedel, K.; Schleper, C.; et al. Holistic assessment of rumen microbiome dynamics through quantitative metatranscriptomics reveals multifunctional redundancy during key steps of anaerobic feed degradation. Msystems 2018, 3, e00038-18. [Google Scholar] [CrossRef]
- Binda, C.; Lopetuso, L.R.; Rizzatti, G.; Gibiino, G.; Cennamo, V.; Gasbarrini, A. Actinobacteria: A relevant minority for the maintenance of gut homeostasis. Dig. Liver Dis. 2018, 50, 421–428. [Google Scholar] [CrossRef]
- Purchiaroni, F.; Tortora, A.; Gabrielli, M.; Bertucci, F.; Gigante, G.; Ianiro, G.; Ojetti, V.; Scarpellini, E.; Gasbarrini, A. The role of intestinal microbiota and the immune system. Eur. Rev. Med Pharmacol. Sci. 2013, 17, 323–333. [Google Scholar] [PubMed]
- Ryan, S.M.; Fitzgerald, G.F.; van Sinderen, D. Screening for and identification of starch-, amylopectin-, and pullulan-degrading activities in bifidobacterial strains. Appl. Environ. Microbiol. 2006, 72, 5289–5296. [Google Scholar] [CrossRef]
- Pokusaeva, K.; Fitzgerald, G.F.; van Sinderen, D. Carbohydrate metabolism in Bifidobacteria. Genes Nutr. 2011, 6, 285–306. [Google Scholar] [CrossRef]
- Rezaeian, M.; Beakes, G.W.; Parker, D.S. Distribution and estimation of anaerobic zoosporic fungi along the digestive tracts of sheep. Mycol. Res. 2004, 108, 1227–1233. [Google Scholar] [CrossRef] [PubMed]
- Kameshwar, A.K.S.; Qin, W. Genome wide analysis reveals the extrinsic cellulolytic and biohydrogen generating abilities of neocallimastigomycota fungi. J. Genom. 2018, 6, 74–87. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, P.; Liu, X.; Zhang, C.; Lu, Q.; Xi, D.; Yang, R.; Wang, S.; Bai, W.; Yang, Z.; et al. The Composition of Fungal Communities in the Rumen of Gayals (Bos frontalis), Yaks (Bos grunniens), and Yunnan and Tibetan Yellow Cattle (Bos taurs). Pol. J. Microbiol. 2019, 68, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Beimforde, C.; Feldberg, K.; Nylinder, S.; Rikkinen, J.; Tuovila, H.; Dörfelt, H.; Gube, M.; Jackson, D.J.; Reitner, J.; Seyfullah, L.; et al. Estimating the Phanerozoic history of the Ascomycota lineages: Combining fossil and molecular data. Mol. Phylogenetics Evol. 2014, 78, 386–398. [Google Scholar] [CrossRef] [PubMed]
- Belanche, A.; Doreau, M.; Edwards, J.E.; Moorby, J.; Pinloche, E.; Newbold, C.J. Shifts in the rumen microbiota due to the type of carbohydrate and level of protein ingested by dairy cattle are associated with changes in rumen fermentation. J. Nutr. 2012, 142, 1684–1692. [Google Scholar] [CrossRef]
- Mohamed, H.; El-Shanawany, A.-R.; Shah, A.M.; Nazir, Y.; Naz, T.; Ullah, S.; Mustafa, K.; Song, Y. Comparative analysis of different isolated oleaginous Mucoromycota fungi for their γ-linolenic acid and carotenoid production. BioMed Res. Int. 2020, 2020, 1–13. [Google Scholar] [CrossRef]
- Dadheech, T.; Jakhesara, S.; Chauhan, P.S.; Pandit, R.; Hinsu, A.; Kunjadiya, A.; Rank, D.; Joshi, C. Draft genome analysis of lignocellulolytic enzymes producing Aspergillus terreus with structural insight of β-glucosidases through molecular docking approach. Int. J. Biol. Macromol. 2018, 125, 181–190. [Google Scholar] [CrossRef]
- Garcia-Mazcorro, J.; Ishaq, S.; Rodriguez-Herrera, M.; Garcia-Hernandez, C.; Kawas, J.; Nagaraja, T. Are there indigenous Saccharomyces in the digestive tract of livestock animal species? Implications for health, nutrition and productivity traits. Animal 2020, 14, 22–30. [Google Scholar] [CrossRef]
- Wei, Y.-Q.; Yang, H.-J.; Long, R.-J.; Wang, Z.-Y.; Cao, B.-B.; Ren, Q.-C.; Wu, T.-T. Characterization of natural co-cultures of Piromyces with Methanobrevibacter ruminantium from yaks grazing on the Qinghai-Tibetan Plateau: A microbial consortium with high potential in plant biomass degradation. AMB Express 2017, 7, 160–172. [Google Scholar] [CrossRef]
- Stein, D.R.; Allen, D.T.; Perry, E.B.; Bruner, J.C.; Gates, K.W.; Rehberger, T.G.; Mertz, K.; Jones, D.; Spicer, L.J. Effects of Feeding Propionibacteria to Dairy Cows on Milk Yield, Milk Components, and Reproduction. J. Dairy Sci. 2006, 89, 111–125. [Google Scholar] [CrossRef]
- Vyas, D.; Uwizeye, A.; Mohammed, R.; Yang, W.Z.; Walker, N.D.; Beauchemin, K.A. The effects of active dried and killed dried yeast on subacute ruminal acidosis, ruminal fermentation, and nutrient digestibility in beef heifers. J. Anim. Sci. 2014, 92, 724–732. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Bai, H.; Zheng, L.; Jiang, H.; Cui, H.; Cao, Y.; Yao, J. Bioactive polysaccharides and oligosaccharides as possible feed additives to manipulate rumen fermentation in rusitec fermenters. Int. J. Biol. Macromol. 2018, 109, 1088–1094. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.K.; Saxena, J. The effect and mode of action of saponins on the microbial populations and fermentation in the rumen and ruminant production. Nutr. Res. Rev. 2009, 22, 204–219. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.; Stiverson, J.; Yu, Z. Effects of quillaja and yucca saponins on communities and select populations of rumen bacteria and archaea, and fermentation in vitro. J. Appl. Microbiol. 2012, 113, 1329–1340. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Wu, X.; He, J. Study on the antifungal activity and mechanism of tea saponin from Camellia oleifera cake. Eur. Food Res. Technol. 2022, 248, 783–795. [Google Scholar] [CrossRef]
- Yan, X.; Yan, B.; Ren, Q.; Dou, J.; Wang, W.; Zhang, J.; Zhou, J.; Long, R.; Ding, L.; Han, J.; et al. Effect of slow-release urea on the composition of ruminal bacteria and fungi communities in yak. Anim. Feed. Sci. Technol. 2018, 244, 18–27. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, H.; Guo, X. Comparative analysis of rumen fermentation parameters and bacterial proles during adaption to different fattening stages in beef cattle fed TMR with various forage silage. Anim. Feed Sci. Technol. 2021, 278, 115006. [Google Scholar] [CrossRef]
- Zhu, N.; Wang, J.; Yu, L.; Zhang, Q.; Chen, K.; Liu, B. Modulation of Growth Performance and Intestinal Microbiota in Chickens Fed Plant Extracts or Virginiamycin. Front. Microbiol. 2019, 10, 1333. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Content, g/kg DM |
|---|---|
| Ingredient | |
| Corn silage | 300 |
| Wheat straw | 100 |
| Corn | 330 |
| Soybean meal | 174 |
| Wheat bran | 66 |
| Salt | 2.4 |
| Sodium bicarbonate | 3.6 |
| Premix 1 | 24 |
| Nutritional level | |
| CP | 161.9 |
| NDF | 376.9 |
| ADF | 243.7 |
| Ca | 7.5 |
| P | 4.0 |
| ME/(MJ/kg) | 10.23 |
| Items | Treatment | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| CON | LT | MT | HT | Treat | Linear | Quadratic | ||
| pH | 6.31 | 6.27 | 6.26 | 6.25 | 0.04 | 0.13 | 0.07 | 0.37 |
| Ammonia N, mg/100 mL | 15.94 | 13.24 | 14.12 | 13.40 | 0.73 | 0.15 | 0.37 | 0.41 |
| TVFAs, mmol/L | 57.50 b | 60.17 a | 58.28 b | 57.31 b | 1.01 | 0.03 | 0.87 | 0.68 |
| VFAs profile, %/100 mol | ||||||||
| Acetate | 64.99 | 64.54 | 65.19 | 64.61 | 0.65 | 0.37 | 0.31 | 0.19 |
| Propionate | 19.68 b | 20.81 a | 20.18 ab | 19.63 b | 0.39 | <0.01 | 0.49 | 0.05 |
| Butyrate | 11.31 | 11.35 | 11.50 | 11.69 | 0.02 | 0.87 | 0.04 | 0.88 |
| Iso-butyrate | 1.56 | 1.31 | 1.41 | 1.44 | 0.24 | 0.40 | 0.62 | 0.28 |
| Valerate, | 0.88 | 0.86 | 0.82 | 0.85 | 0.04 | 0.34 | 0.40 | 0.95 |
| Iso-valerate | 1.55 | 1.19 | 1.42 | 1.76 | 0.36 | 0.07 | 0.10 | 0.07 |
| A:P | 3.30 a | 3.11 b | 3.18 ab | 3.29 a | 0.13 | 0.02 | 0.86 | 0.01 |
| Items | CON | LT | MT | HT | SEM | p-Value |
|---|---|---|---|---|---|---|
| chao1 index | 1423.88 a | 1329.05 ab | 1291.31 b | 1353.30 ab | 46.23 | 0.03 |
| faith_pd index | 75.70 a | 74.64 a | 66.25 b | 73.83 ab | 5.32 | 0.02 |
| Shannon index | 9.00 | 8.90 | 8.36 | 8.77 | 0.39 | 0.09 |
| simpson index | 0.99 | 0.99 | 0.97 | 0.98 | 0.02 | 0.59 |
| Items | CON | LT | MT | HT | SEM | p-Value |
|---|---|---|---|---|---|---|
| chao1 index | 79.50 | 86.25 | 75.50 | 70.00 | 5.60 | 0.84 |
| faith_pd index | 7.42 | 7.27 | 7.19 | 5.87 | 0.33 | 0.15 |
| Shannon index | 3.85 | 3.71 | 3.39 | 3.47 | 0.09 | 0.39 |
| simpson index | 0.85 | 0.82 | 0.81 | 0.80 | 0.04 | 0.23 |
| Items | CON | LT | MT | HT | SEM | p-Value |
|---|---|---|---|---|---|---|
| Phylum level | ||||||
| Bacteroidetes | 68.02 | 66.20 | 67.89 | 69.12 | 2.80 | 0.48 |
| Firmicutes | 28.33 | 28.19 | 27.53 | 25.51 | 1.07 | 0.82 |
| Proteobacteria | 0.66 b | 1.14 ab | 0.91 b | 1.64 a | 0.13 | <0.01 |
| Spirochaetes | 0.55 | 0.92 | 0.79 | 0.66 | 0.14 | 0.45 |
| Euryarchaeota | 0.53 | 0.75 | 0.73 | 0.27 | 0.17 | 0.49 |
| Verrucomicrobia | 0.43 | 0.51 | 0.51 | 0.31 | 0.07 | 0.36 |
| Actinobacteria | 0.33 b | 0.54 a | 0.23 b | 0.11 b | 0.06 | 0.03 |
| Tenericutes | 0.26 | 0.48 | 0.44 | 0.23 | 0.05 | 0.12 |
| TM7 | 0.28 | 0.27 | 0.35 | 0.30 | 0.03 | 0.74 |
| unclassified | 0.23 | 0.67 | 0.14 | 0.31 | 0.25 | 0.06 |
| Genus level | ||||||
| unclassified | 41.79 | 45.95 | 49.45 | 48.82 | 3.63 | 0.12 |
| Prevotellaceae_Prevotella | 29.64 | 26.59 | 26.58 | 32.04 | 1.19 | 0.40 |
| Succiniclasticum | 8.78 a | 6.58 ab | 5.84 b | 4.39 b | 0.56 | 0.01 |
| Clostridium | 1.35 | 1.54 | 1.30 | 1.85 | 0.16 | 0.35 |
| Bacteroides | 3.80 a | 1.24 b | 0.79 b | 1.13 b | 0.45 | 0.02 |
| Faecalibacterium | 1.85 | 2.29 | 1.07 | 0.51 | 0.32 | 0.62 |
| f_Paraprevotellaceae;g_YRC22 | 1.06 | 1.04 | 1.50 | 1.77 | 0.16 | 0.15 |
| f_Paraprevotellaceae;g-CF231 | 1.41 | 1.40 | 1.53 | 1.33 | 0.17 | 0.83 |
| Oscillospira | 0.96 | 1.44 | 1.01 | 0.78 | 0.15 | 0.29 |
| Ruminococcaceae_Ruminococcus | 0.63 | 1.26 | 1.03 | 0.59 | 0.18 | 0.24 |
| Items | CON | LT | MT | HT | SEM | p-Value |
|---|---|---|---|---|---|---|
| Phylum level | ||||||
| Ascomycota | 51.82 b | 75.52 a | 66.31 ab | 52.40 b | 3.99 | 0.03 |
| Neocallimastigomycota | 23.06 | 9.75 | 13.57 | 28.04 | 4.95 | 0.53 |
| Mucoromycota | 17.22 | 7.04 | 16.62 | 12.05 | 2.35 | 0.36 |
| Cryptomycota | 5.69 | 3.42 | 1.42 | 3.24 | 0.97 | 0.30 |
| Basidiomycota | 2.14 | 4.03 | 1.88 | 4.17 | 0.50 | 0.21 |
| LKM15 | 0.01 | 0.11 | 0.11 | 0.02 | 0.02 | 0.28 |
| unclassified | 0.02 | 0.08 | 0.04 | 0.02 | 0.01 | 0.36 |
| Zoopagomycota | 0.02 | 0.04 | 0.02 | 0.05 | 0.01 | 0.12 |
| Chytridiomycota | 0.02 | 0.01 | 0.03 | 0.01 | 0.01 | 0.94 |
| Genus level | ||||||
| Thermomyces | 29.87 | 36.78 | 31.54 | 32.60 | 2.34 | 0.83 |
| unclassified | 17.01 | 11.47 | 17.35 | 19.13 | 1.64 | 0.39 |
| Saccharomyces | 4.21 b | 13.27 a | 16.15 a | 2.43 b | 2.35 | 0.03 |
| Piromyces | 9.73 | 4.82 | 3.61 | 16.07 | 2.58 | 0.07 |
| Rhizomucor | 11.03 | 2.56 | 9.16 | 2.31 | 1.71 | 0.21 |
| Aspergillus | 4.02 b | 9.53 a | 4.12 b | 2.75 b | 1.02 | 0.04 |
| Chaetomium | 3.94 ab | 3.63 ab | 6.99 a | 2.11 b | 0.79 | 0.03 |
| LKM11 | 5.53 | 2.89 | 1.22 | 3.21 | 0.95 | 0.28 |
| Lichtheimia | 2.04 | 0.90 | 1.24 | 4.19 | 0.97 | 0.60 |
| Cystobasidium | 1.24 | 2.25 | 1.14 | 3.19 | 0.52 | 0.76 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qu, X.; Raza, S.H.A.; Zhao, Y.; Deng, J.; Ma, J.; Wang, J.; Alkhorayef, N.; Alkhalil, S.S.; Pant, S.D.; Lei, H.; et al. Effect of Tea Saponins on Rumen Microbiota and Rumen Function in Qinchuan Beef Cattle. Microorganisms 2023, 11, 374. https://doi.org/10.3390/microorganisms11020374
Qu X, Raza SHA, Zhao Y, Deng J, Ma J, Wang J, Alkhorayef N, Alkhalil SS, Pant SD, Lei H, et al. Effect of Tea Saponins on Rumen Microbiota and Rumen Function in Qinchuan Beef Cattle. Microorganisms. 2023; 11(2):374. https://doi.org/10.3390/microorganisms11020374
Chicago/Turabian StyleQu, Xiaopeng, Sayed Haidar Abbas Raza, Yanqing Zhao, Jiahan Deng, Jing Ma, Juze Wang, Nada Alkhorayef, Samia S. Alkhalil, Sameer D. Pant, Hongtao Lei, and et al. 2023. "Effect of Tea Saponins on Rumen Microbiota and Rumen Function in Qinchuan Beef Cattle" Microorganisms 11, no. 2: 374. https://doi.org/10.3390/microorganisms11020374
APA StyleQu, X., Raza, S. H. A., Zhao, Y., Deng, J., Ma, J., Wang, J., Alkhorayef, N., Alkhalil, S. S., Pant, S. D., Lei, H., & Zan, L. (2023). Effect of Tea Saponins on Rumen Microbiota and Rumen Function in Qinchuan Beef Cattle. Microorganisms, 11(2), 374. https://doi.org/10.3390/microorganisms11020374