

Rapid and Easy-Read Porcine Circovirus Type 4 Detection with CRISPR–Cas13a-Based Lateral Flow Strip

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plasmids and Viruses
2.2. RPA Primer Design and crisprRNA (crRNA) Preparation
2.3. Nucleic Acid Preparation
2.4. Evaluating and Optimizing RPA Reactions
2.5. Cas13a Nucleic Acid Detection
2.6. PCV4 Analytical Sensitivity and Specificity by the RPA-Cas13a-LFD
2.7. Evaluating RPA-Cas13a-LFD Tolerance to Serum Inhibitors
2.8. Assay Validation Using Clinical Samples
2.9. Ethics Statement
3. Results
3.1. RPA Product Detection
3.2. Establishing the CRISPR–Cas13a-LFD
3.3. CRISPR–Cas13a-LFD Analytical Sensitivity
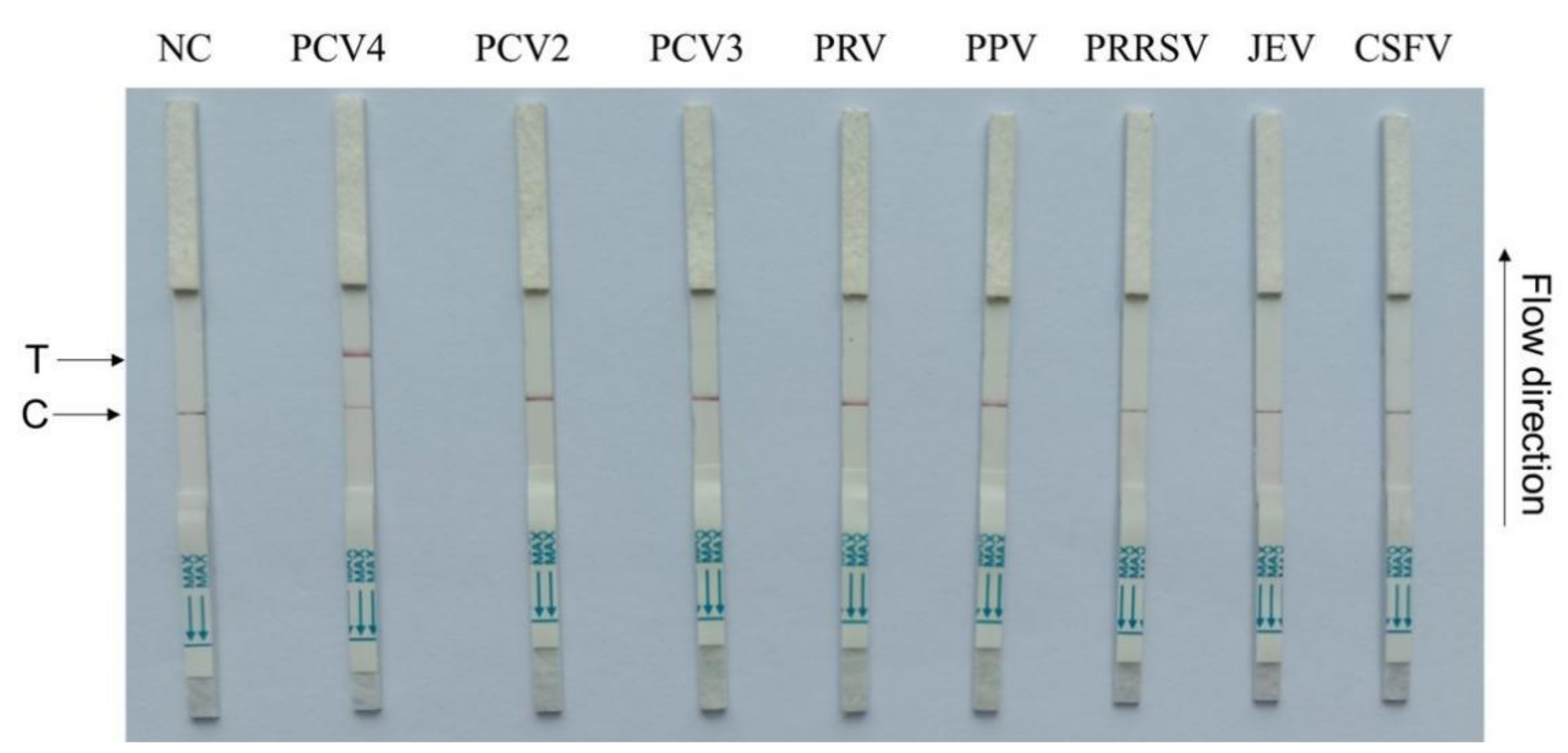
3.4. CRISPR–Cas13a-LFD Analytical Specificity
3.5. Evaluating RPA-Cas13a-LFD Tolerance to Serum Inhibitors
3.6. Clinical Sample Detection Using CRISPR–Cas13a-LFD
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Finsterbusch, T.; Mankertz, A. Porcine circoviruses—Small but powerful. Virus Res. 2009, 143, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Palinski, R.; Pineyro, P.; Shang, P.; Yuan, F.; Guo, R.; Fang, Y.; Byers, E.; Hause, B.M. A Novel Porcine Circovirus Distantly Related to Known Circoviruses Is Associated with Porcine Dermatitis and Nephropathy Syndrome and Reproductive Failure. J. Virol. 2017, 91, e01879-16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Bai, C.; Ge, K.; Li, Y.; Gao, W.; Jiang, S.; Wang, Y. Establishment of an SYBR Green-based real-time PCR assay for porcine circovirus type 4 detection. J. Virol. Methods 2020, 285, 113963. [Google Scholar] [CrossRef]
- Correa-Fiz, F.; Franzo, G.; Llorens, A.; Huerta, E.; Sibila, M.; Kekarainen, T.; Segales, J. Porcine circovirus 2 (PCV2) population study in experimentally infected pigs developing PCV2-systemic disease or a subclinical infection. Sci. Rep. 2020, 10, 17747. [Google Scholar] [CrossRef] [PubMed]
- Allan, G.M.; Ellis, J.A. Porcine circoviruses: A review. J. Vet. Diagn. Invest. 2000, 12, 3–14. [Google Scholar] [CrossRef]
- Allan, G.; Krakowka, S.; Ellis, J.; Charreyre, C. Discovery and evolving history of two genetically related but phenotypically different viruses, porcine circoviruses 1 and 2. Virus Res. 2012, 164, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Karuppannan, A.K.; Castro, A.; Xiao, C.T. Porcine circoviruses: Current status, knowledge gaps and challenges. Virus Res. 2020, 286, 198044. [Google Scholar] [CrossRef]
- Wang, D.; Mai, J.; Lei, B.; Zhang, Y.; Yang, Y.; Wang, N. Structure, Antigenic Properties, and Highly Efficient Assembly of PCV4 Capsid Protein. Front. Vet. Sci. 2021, 8, 695466. [Google Scholar] [CrossRef]
- Sun, W.; Du, Q.; Han, Z.; Bi, J.; Lan, T.; Wang, W.; Zheng, M. Detection and genetic characterization of porcine circovirus 4 (PCV4) in Guangxi, China. Gene 2021, 773, 145384. [Google Scholar] [CrossRef]
- Li, X.; Chen, S.; Niu, G.; Zhang, X.; Ji, W.; Ren, Y.; Zhang, L.; Ren, L. Porcine Circovirus Type 4 Strains Circulating in China Are Relatively Stable and Have Higher Homology with Mink Circovirus than Other Porcine Circovirus Types. Int. J. Mol. Sci. 2022, 23, 3288. [Google Scholar] [CrossRef]
- Ge, M.; Hu, W.Q.; Ning, K.M.; Li, S.Y.; Xiao, C.T. The seroprevalence of the newly identified porcine circovirus type 4 in China investigated by an enzymed-linked immunosorbent assay. Transbound. Emerg. Dis. 2021, 68, 2910–2914. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.G.; Do, H.Q.; Huynh, T.M.; Park, Y.H.; Park, B.K.; Chung, H.C. Molecular-based detection, genetic characterization and phylogenetic analysis of porcine circovirus 4 from Korean domestic swine farms. Transbound. Emerg. Dis. 2022, 69, 538–548. [Google Scholar] [CrossRef]
- Franzo, G.; Ruiz, A.; Grassi, L.; Sibila, M.; Drigo, M.; Segales, J. Lack of Porcine circovirus 4 Genome Detection in Pig Samples from Italy and Spain. Pathogens 2020, 9, 433. [Google Scholar] [CrossRef]
- Veenema, T.G.; Toke, J. Early detection and surveillance for biopreparedness and emerging infectious diseases. Online J. Issues Nurs. 2006, 11, 3. [Google Scholar] [CrossRef]
- Hattermann, K.; Maerz, A.; Slanina, H.; Schmitt, C.; Mankertz, A. Assessing the risk potential of porcine circoviruses for xenotransplantation: Consensus primer-PCR-based search for a human circovirus. Xenotransplantation 2004, 11, 547–550. [Google Scholar] [CrossRef]
- Ouardani, M.; Wilson, L.; Jette, R.; Montpetit, C.; Dea, S. Multiplex PCR for detection and typing of porcine circoviruses. J. Clin. Microbiol. 1999, 37, 3917–3924. [Google Scholar] [CrossRef]
- Yang, K.; Jiao, Z.; Zhou, D.; Guo, R.; Duan, Z.; Tian, Y. Development of a multiplex PCR to detect and discriminate porcine circoviruses in clinical specimens. BMC Infect. Dis. 2019, 19, 778. [Google Scholar] [CrossRef] [PubMed]
- Chang, G.N.; Hwang, J.F.; Chen, J.T.; Tsen, H.Y.; Wang, J.J. Fast diagnosis and quantification for porcine circovirus type 2 (PCV-2) using real-time polymerase chain reaction. J. Microbiol. Immunol. Infect. 2010, 43, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Song, Y.; Deng, X.; Vanegas, J.A.; You, Z.; Zhang, Y.; Weng, Z.; Avery, L.; Dieckhaus, K.D.; Peddi, A.; et al. Engineered LwaCas13a with enhanced collateral activity for nucleic acid detection. Nat. Chem. Biol. 2023, 19, 45–54. [Google Scholar] [CrossRef]
- Fozouni, P.; Son, S.; Diaz de Leon Derby, M.; Knott, G.J.; Gray, C.N.; D’Ambrosio, M.V.; Zhao, C.; Switz, N.A.; Kumar, G.R.; Stephens, S.I.; et al. Amplification-free detection of SARS-CoV-2 with CRISPR-Cas13a and mobile phone microscopy. Cell 2021, 184, 323–333 e329. [Google Scholar] [CrossRef]
- Myhrvold, C.; Freije, C.A.; Gootenberg, J.S.; Abudayyeh, O.O.; Metsky, H.C.; Durbin, A.F.; Kellner, M.J.; Tan, A.L.; Paul, L.M.; Parham, L.A.; et al. Field-deployable viral diagnostics using CRISPR-Cas13. Science 2018, 360, 444–448. [Google Scholar] [CrossRef] [PubMed]
- Gootenberg, J.S.; Abudayyeh, O.O.; Lee, J.W.; Essletzbichler, P.; Dy, A.J.; Joung, J.; Verdine, V.; Donghia, N.; Daringer, N.M.; Freije, C.A.; et al. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science 2017, 356, 438–442. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Li, F.; Chen, Q.; Wu, J.; Duan, J.; Lei, X.; Zhang, Y.; Zhao, D.; Bu, Z.; Yin, H. Rapid detection of African swine fever virus using Cas12a-based portable paper diagnostics. Cell Discov. 2020, 6, 18. [Google Scholar] [CrossRef] [PubMed]
- Patchsung, M.; Jantarug, K.; Pattama, A.; Aphicho, K.; Suraritdechachai, S.; Meesawat, P.; Sappakhaw, K.; Leelahakorn, N.; Ruenkam, T.; Wongsatit, T.; et al. Clinical validation of a Cas13-based assay for the detection of SARS-CoV-2 RNA. Nat. Biomed. Eng. 2020, 4, 1140–1149. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Xu, L.; Liu, Q.; Cao, Y.; Yang, K.; Song, X.; Shao, Y.; Tu, J.; Qi, K. Enzymatic recombinase amplification coupled with CRISPR-Cas12a for ultrasensitive, rapid, and specific Porcine circovirus 3 detection. Mol. Cell Probes. 2021, 59, 101763. [Google Scholar] [CrossRef] [PubMed]
- Ai, J.W.; Zhou, X.; Xu, T.; Yang, M.; Chen, Y.; He, G.Q.; Pan, N.; Cai, Y.; Li, Y.; Wang, X.; et al. CRISPR-based rapid and ultra-sensitive diagnostic test for Mycobacterium tuberculosis. Emerg. Microbes. Infect. 2019, 8, 1361–1369. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Yin, L.; Dong, Y.; Peng, L.; Liu, G.; Man, S.; Ma, L. CRISPR-Cas13a based bacterial detection platform: Sensing pathogen Staphylococcus aureus in food samples. Anal. Chim. Acta 2020, 1127, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, H.; Liu, C.; Peng, L.; Khan, H.; Cui, L.; Huang, R.; Wu, C.; Shen, S.; Wang, S.; et al. CRISPR-Cas13a Nanomachine Based Simple Technology for Avian Influenza A (H7N9) Virus On-Site Detection. J. Biomed. Nanotechnol. 2019, 15, 790–798. [Google Scholar] [CrossRef]
- Niu, G.; Zhang, X.; Ji, W.; Chen, S.; Li, X.; Yang, L.; Zhang, L.; Ouyang, H.; Li, C.; Ren, L. Porcine circovirus 4 rescued from an infectious clone is replicable and pathogenic in vivo. Transbound. Emerg. Dis. 2022, 69, e1632–e1641. [Google Scholar] [CrossRef]
- Chen, N.; Xiao, Y.; Li, X.; Li, S.; Xie, N.; Yan, X.; Li, X.; Zhu, J. Development and application of a quadruplex real-time PCR assay for differential detection of porcine circoviruses (PCV1 to PCV4) in Jiangsu province of China from 2016 to 2020. Transbound. Emerg. Dis. 2021, 68, 1615–1624. [Google Scholar] [CrossRef]
- Zhang, H.H.; Hu, W.Q.; Li, J.Y.; Liu, T.N.; Zhou, J.Y.; Opriessnig, T.; Xiao, C.T. Novel circovirus species identified in farmed pigs designated as Porcine circovirus 4, Hunan province, China. Transbound. Emerg. Dis. 2020, 67, 1057–1061. [Google Scholar] [CrossRef]
- Xu, T.; Chen, X.M.; Fu, Y.; Ai, Y.; Wang, D.M.; Wei, Z.Y.; Li, X.S.; Zheng, L.L.; Chen, H.Y. Cross-species transmission of an emerging porcine circovirus (PCV4): First molecular detection and retrospective investigation in dairy cows. Vet. Microbiol. 2022, 273, 109528. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Hou, C.; Wang, Z.; Meng, P.; Chen, H.; Cao, H. First complete genomic sequence analysis of porcine circovirus type 4 (PCV4) in wild boars. Vet. Microbiol 2022, 273, 109547. [Google Scholar] [CrossRef]
- Wei, N.; Zheng, B.; Niu, J.; Chen, T.; Ye, J.; Si, Y.; Cao, S. Rapid Detection of Genotype II African Swine Fever Virus Using CRISPR Cas13a-Based Lateral Flow Strip. Viruses 2022, 14, 179. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.Y.; Xu, T.; Zhang, L.H.; Cui, J.T.; Zhang, Y.H.; Li, X.S.; Zheng, L.L.; Chen, H.Y. Simultaneous detection and differentiation of porcine circovirus 3 and 4 using a SYBR Green capital I, Ukrainian-based duplex quantitative PCR assay. J. Virol. Methods 2021, 293, 114152. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, Y.; Li, C.; Yang, K.; Li, Z.; Shang, W.; Song, X.; Shao, Y.; Qi, K.; Tu, J. Rapid detection of porcine circovirus type 4 via multienzyme isothermal rapid amplification. Front. Vet. Sci. 2022, 9, 949172. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′-3′) | |
|---|---|---|
| Primer 1 | Cap-RPA-F1 Cap-RPA-R1 | TGCTGTGGTTTGCCAGGACATCATAAGTTT TCCCATTTGCATATTACCGGATCAGAAAGG |
| Primer 2 | Cap-RPA-F2 Cap-RPA-R2 | GACATCATAAGTTTGGTTTTTCCCTTCCCCC TTTACAGCCTCCCATTTGCATATTACCGGAT |
| Primer 3 | Cap-RPA-F3 Cap-RPA-R3 | TCATAAGTTTGGTTTTTCCCTTCCCCCACATAG CACGCCCTCTTGGAACGTTGGACATTACGATTT |
| Primer 4 | Cap-RPA-F4 Cap-RPA-R4 | CTGCTGCTGTGGTTTGCCAGGACATCATAAGTTT AGCCTCCCATTTGCATATTACCGGATCAGAAAGG |
| Primer 5 | Cap-RPA-F5 Cap-RPA-R5 | TTTTTCCCTTCCCCCACATAGTCTCCATCCAGTT GCCCTCTTGGAACGTTGGACATTACGATTTCAAA |
| Name | Sequence (5′-3′) |
|---|---|
| crRNA1 | AACTGGATGGAGACTATGTGGGGGAAGG |
| crRNA2 | GAAAGGTCAAAGTCGAATTTCTGCCACT |
| crRNA3 | TTACAGCCTCCCATTTGCATATTACCGG |
| crRNA4 | AACTGGATGGAGACTATGTGGGGGAAGG |
| crRNA5 | TTACAGCCTCCCATTTGCATATTACCGG |
| FAM-N6-BIO probe | /56-FAM/mArArUrGrGrCmAmArArUrGrGrCmA/3 Bio/ |
| Primer Name | Peak Fragment Size (bp) | Theoretical Fragment Size (bp) | Product Concentration (ng/µL) |
|---|---|---|---|
| PCV4 Primer1 | 168 | 162 | 1.44 |
| PCV4 Primer2 | 161 | 155 | 1.56 |
| PCV4 Primer3 | 244 | 231 | 1.09 |
| PCV4 Primer4 | 172 | 170 | 1.7 |
| PCV4 Primer5 | 214 | 216 | 3.37 |
| Assay | Number of Samples | |
|---|---|---|
| Positive | Negative | |
| RT-PCR | 10 | 5 |
| Cas13a lateral flow detection | 10 | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Zhu, X.; Yin, D.; Cai, C.; Liu, H.; Yang, Y.; Guo, Z.; Yin, L.; Shen, X.; Dai, Y.; et al. Rapid and Easy-Read Porcine Circovirus Type 4 Detection with CRISPR–Cas13a-Based Lateral Flow Strip. Microorganisms 2023, 11, 354. https://doi.org/10.3390/microorganisms11020354
Wang J, Zhu X, Yin D, Cai C, Liu H, Yang Y, Guo Z, Yin L, Shen X, Dai Y, et al. Rapid and Easy-Read Porcine Circovirus Type 4 Detection with CRISPR–Cas13a-Based Lateral Flow Strip. Microorganisms. 2023; 11(2):354. https://doi.org/10.3390/microorganisms11020354
Chicago/Turabian StyleWang, Jieru, Xiaojie Zhu, Dongdong Yin, Chang Cai, Hailong Liu, Yuqing Yang, Zishi Guo, Lei Yin, Xuehuai Shen, Yin Dai, and et al. 2023. "Rapid and Easy-Read Porcine Circovirus Type 4 Detection with CRISPR–Cas13a-Based Lateral Flow Strip" Microorganisms 11, no. 2: 354. https://doi.org/10.3390/microorganisms11020354
APA StyleWang, J., Zhu, X., Yin, D., Cai, C., Liu, H., Yang, Y., Guo, Z., Yin, L., Shen, X., Dai, Y., & Pan, X. (2023). Rapid and Easy-Read Porcine Circovirus Type 4 Detection with CRISPR–Cas13a-Based Lateral Flow Strip. Microorganisms, 11(2), 354. https://doi.org/10.3390/microorganisms11020354