
Harnessing Novel Soil Bacteria for Beneficial Interactions with Soybean

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
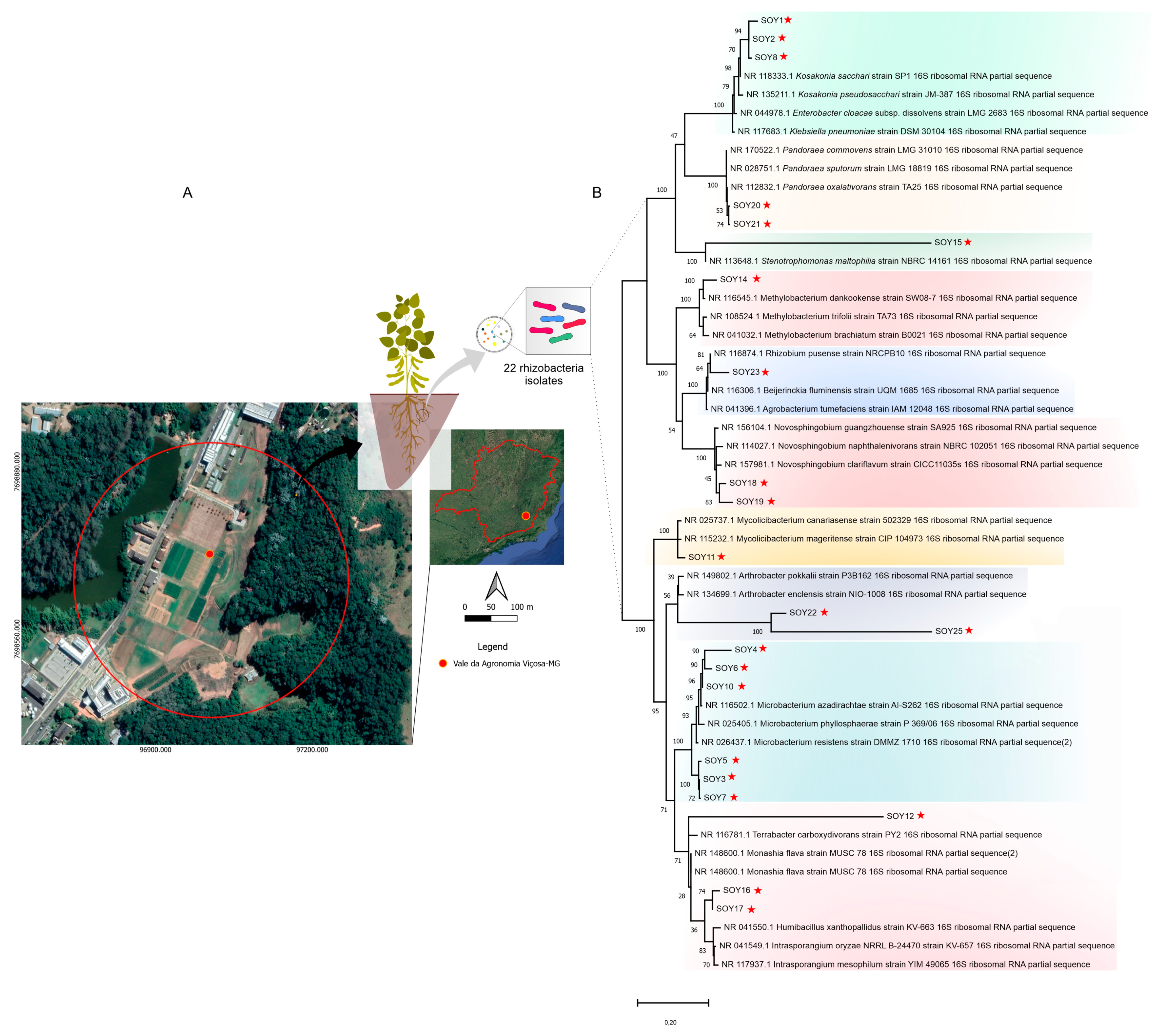
2.1. Soil Sampling, Preparation of Media, and Isolation Procedures
2.2. DNA Fingerprinting
2.3. DNA Extraction and 16S rRNA Sequencing, Processing, and Analysis
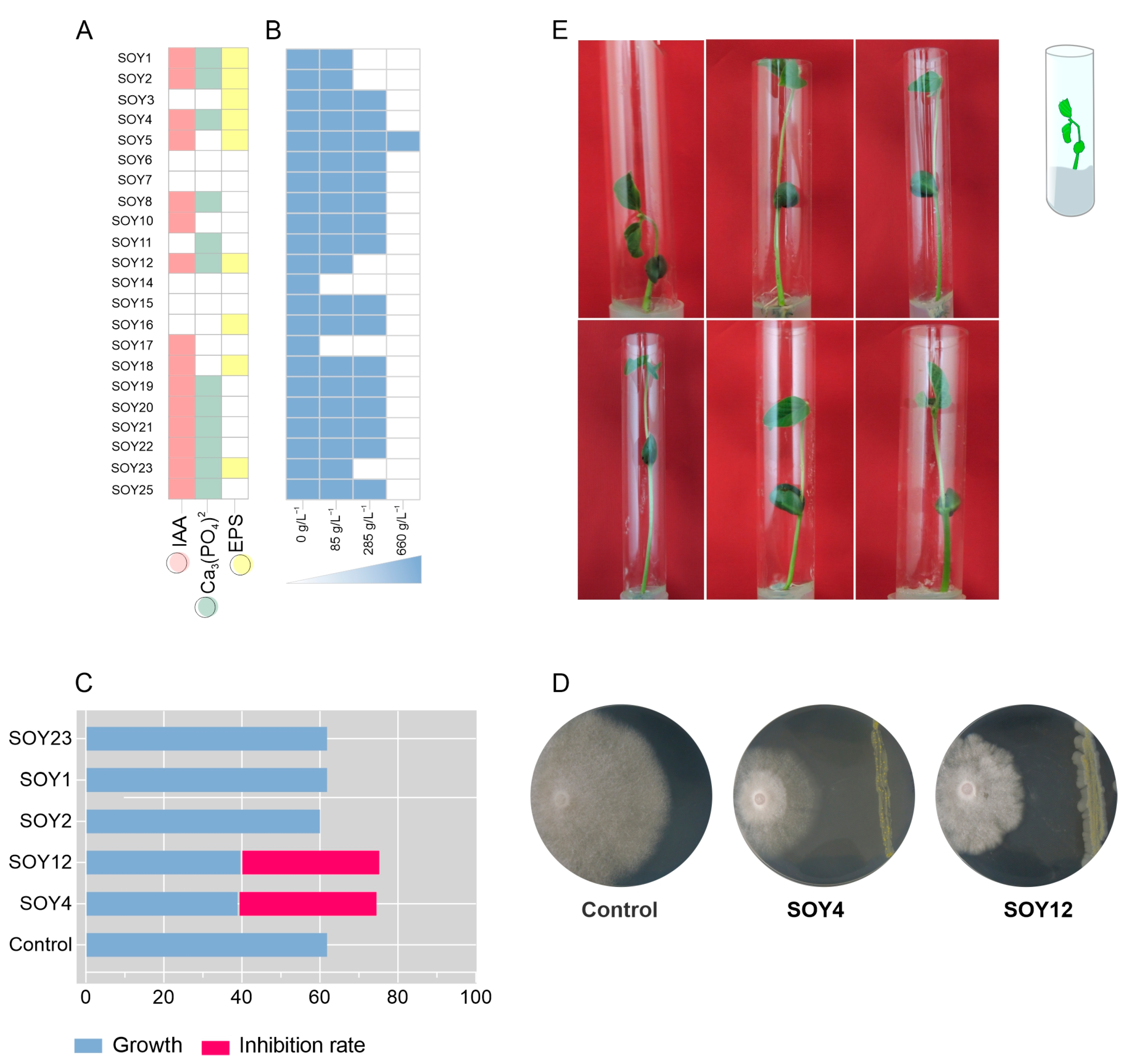
2.4. Screening of Bacterial Isolates for In Vitro Plant Growth-Promotion Traits and Biocontrol
2.4.1. Bacterial Growth under Reduced Water Availability
2.4.2. Exopolysaccharide Production
2.4.3. Indole Acetic Acid (IAA) Production
2.4.4. Phosphate Solubilization Assay
2.4.5. Siderophore Production
2.4.6. Biocontrol Test
2.5. Plant Growth Promotion in Greenhouse Experiment
2.6. DNA Extraction and Whole-Genome Sequencing
2.7. Genome Assembly
2.8. Data Retrieving from a Public Database and Bioinformatics Analysis
3. Results
3.1. The Selection of Distinct Rhizobacteria with the Potential for Novel Taxonomic Species
3.2. The Novel Bacterial Species Exhibited Plant Growth-Promoting Traits
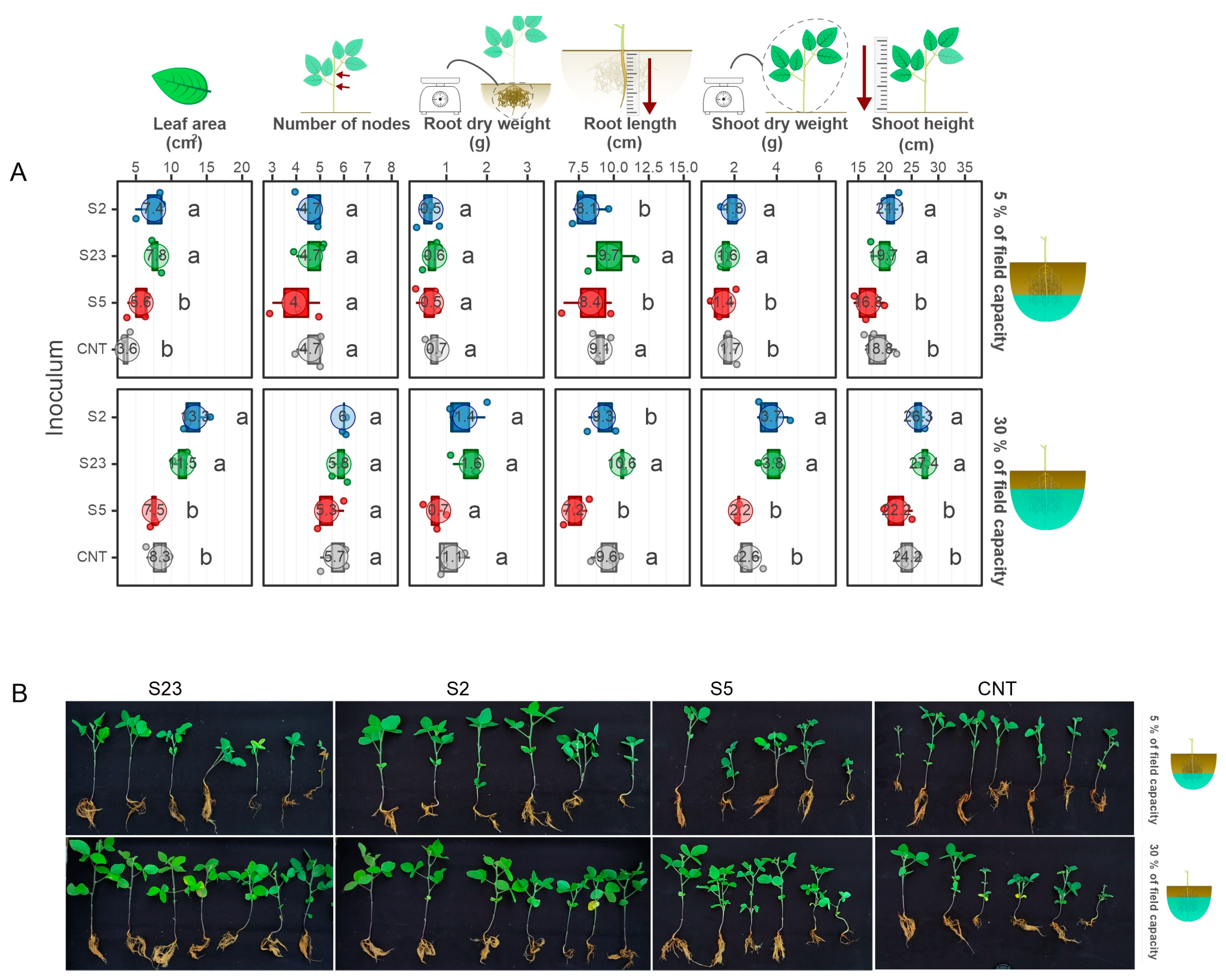
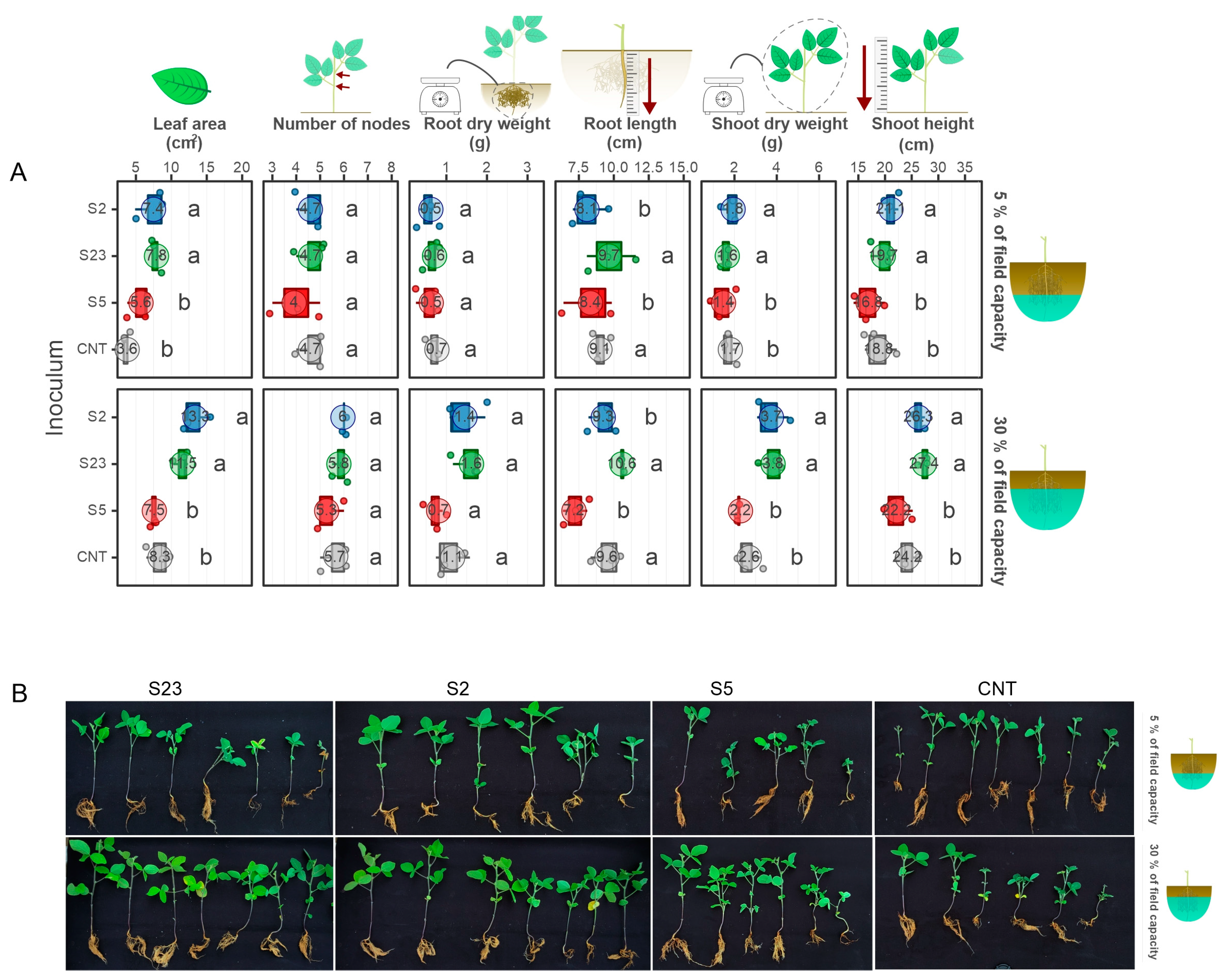
3.3. Two Isolates Showed the Ability to Enhance Soybean Growth under Drought in Greenhouse Conditions
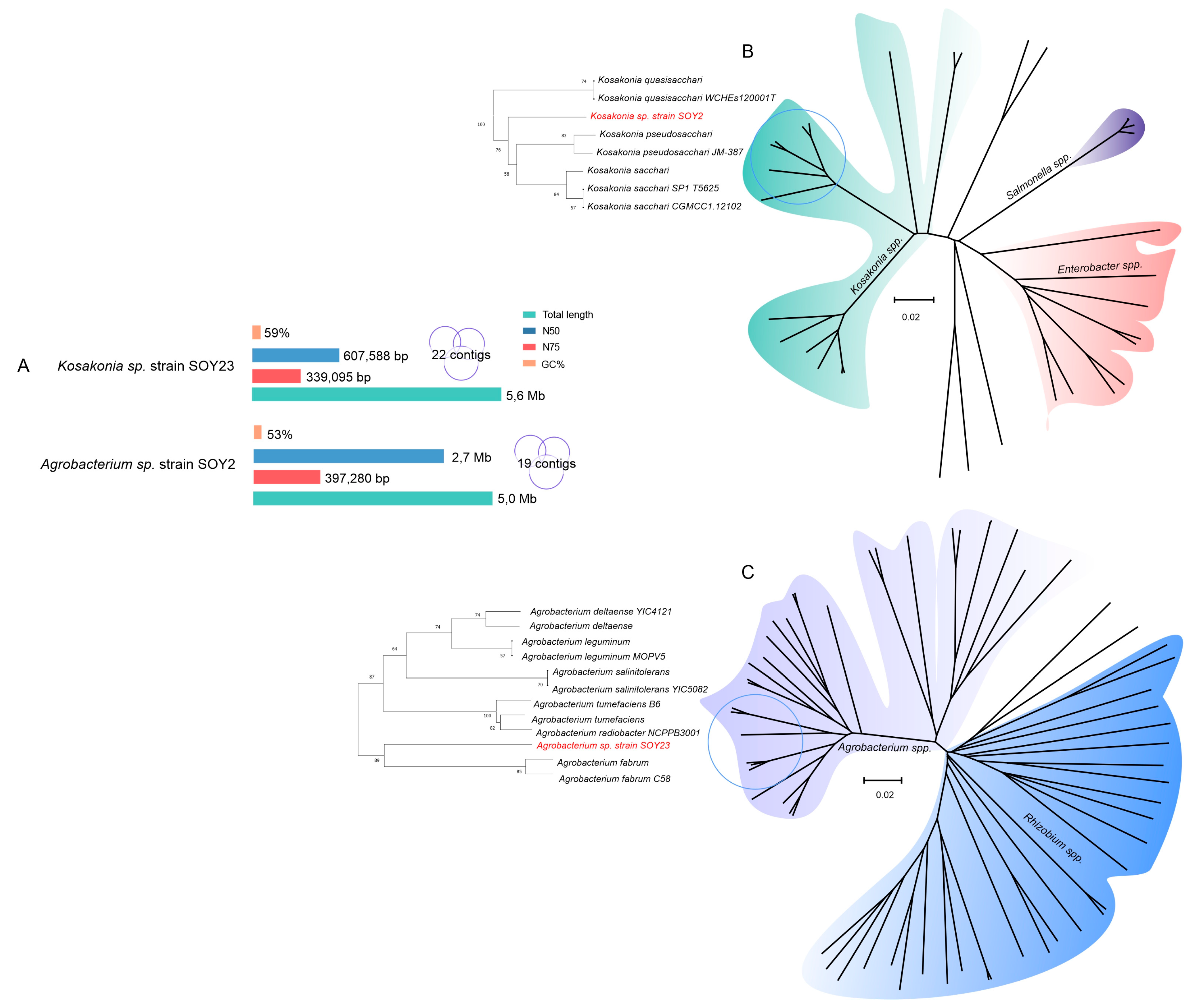
3.4. The Genomic Sequences of Kosakonia and Agrobacterium
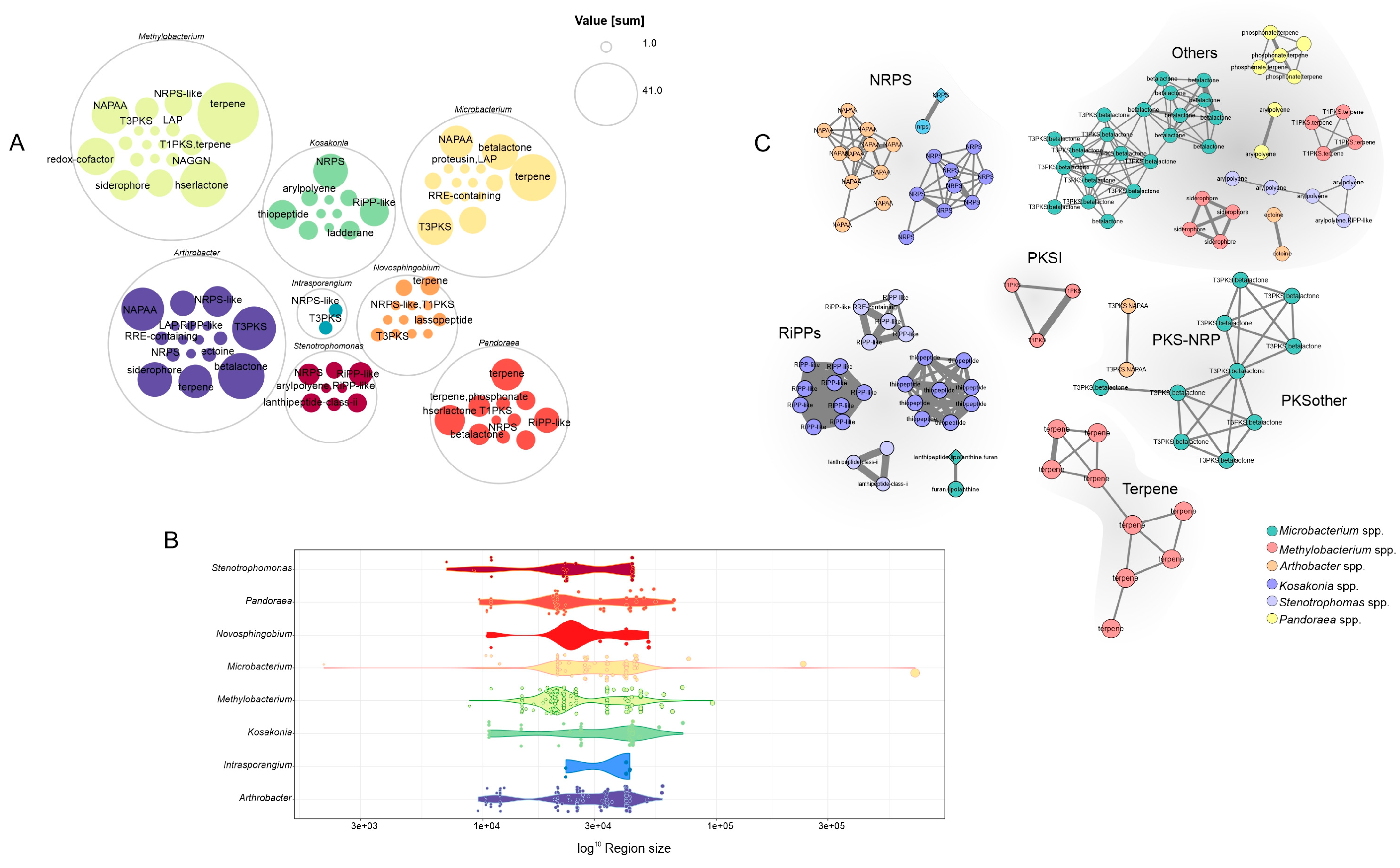
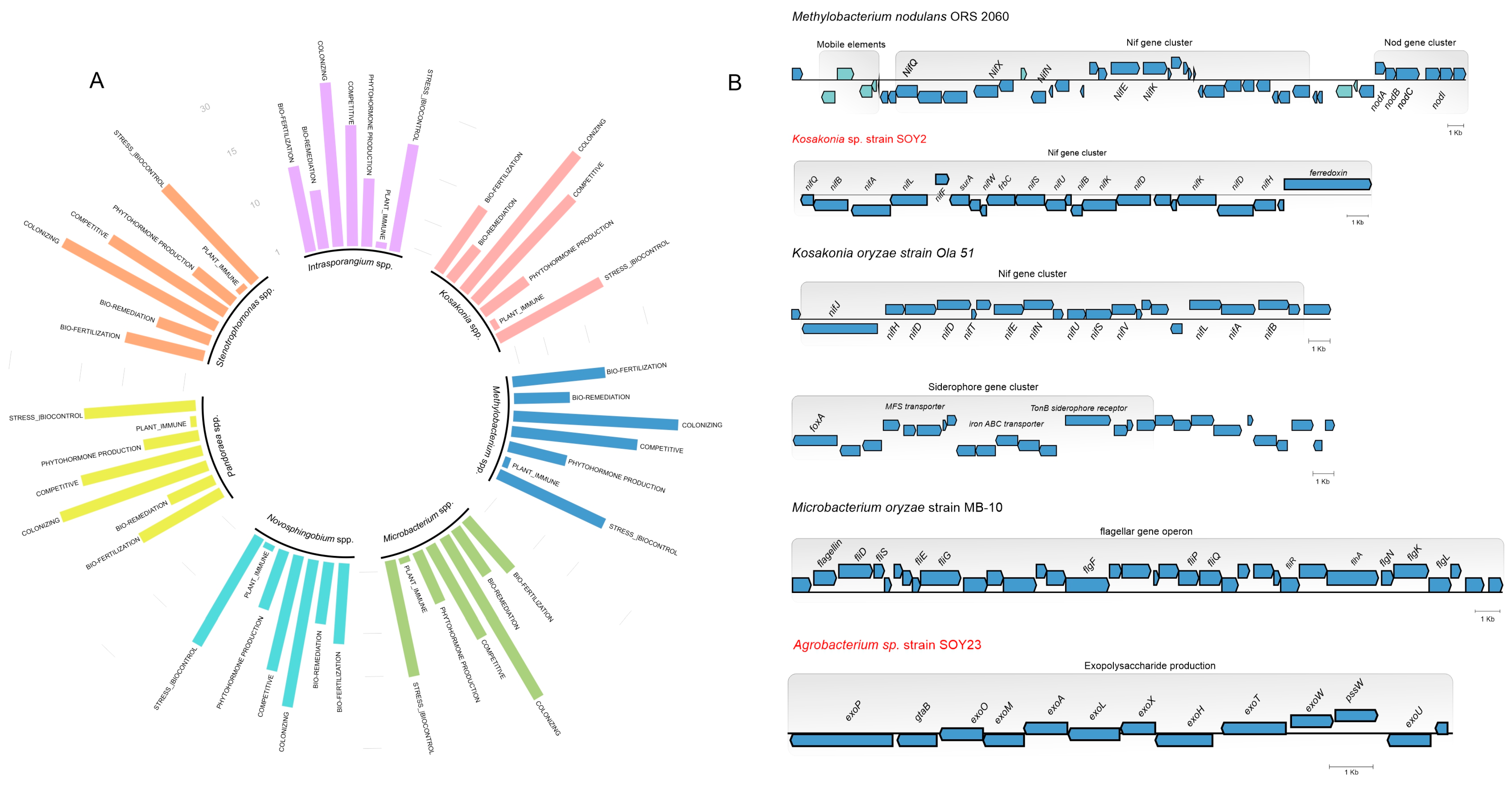
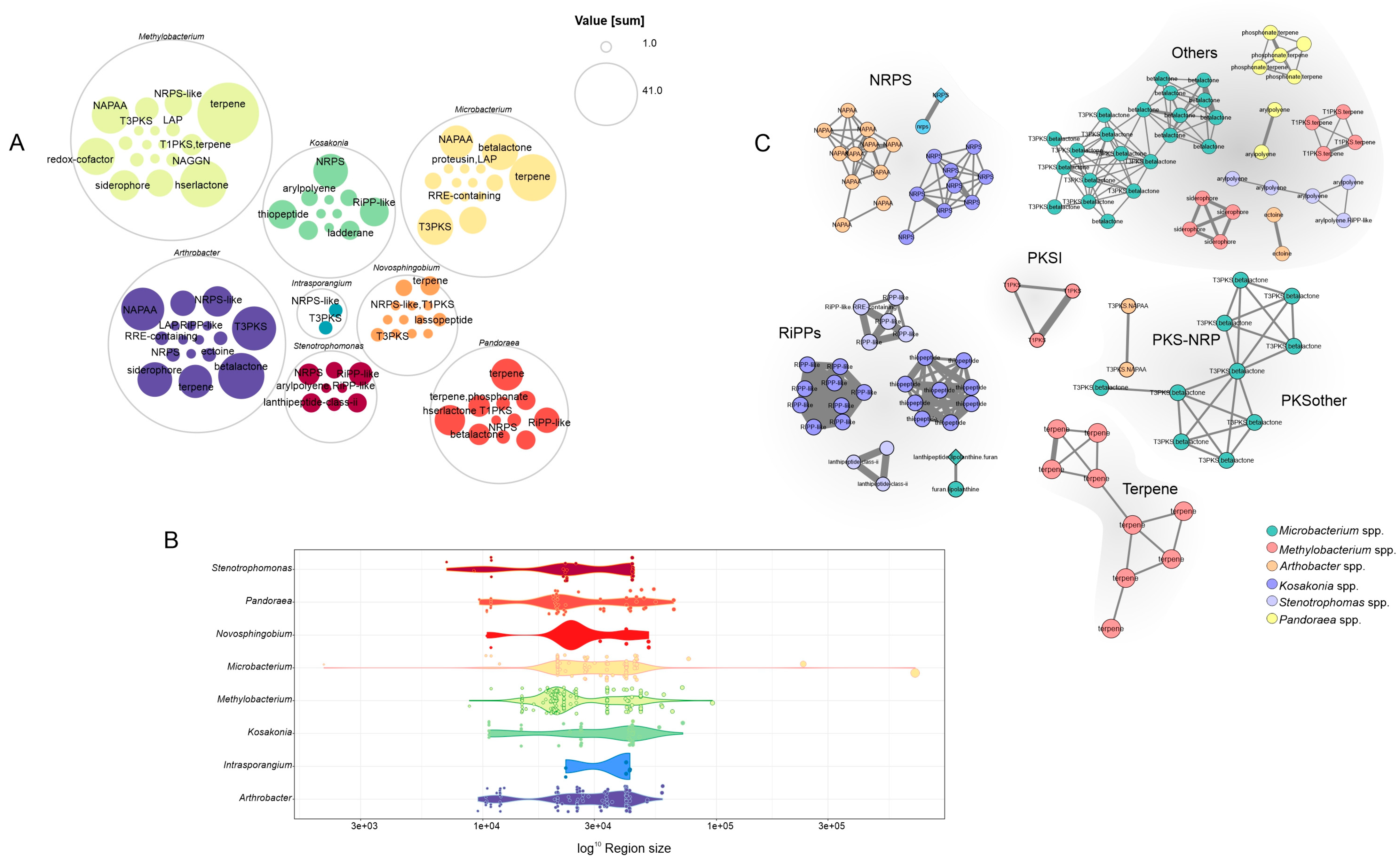
3.5. A Genome Mining Analysis of Novel Species of Soil Bacteria Revealed Several Proteins with Traits That Promote Plant Growth
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berg, G.; Grube, M.; Schloter, M.; Smalla, K. Unraveling the plant microbiome: Looking back and future perspectives. Front. Microbiol. 2014, 5, 148. [Google Scholar] [CrossRef] [Green Version]
- Dastogeer, K.M.G.; Tumpa, F.H.; Sultana, A.; Akter, M.A.; Chakraborty, A. Plant microbiome–an account of the factors that shape community composition and diversity. Curr. Plant Biol. 2020, 23, 100161. [Google Scholar] [CrossRef]
- Naylor, D.; Coleman-Derr, D. Drought stress and root-associated bacterial communities. Front. Plant Sci. 2018, 8, 2223. [Google Scholar] [CrossRef]
- Staley, J.T.; Konopka, A. Measurement of in situ activities of nonphotosynthetic microorganisms in aquatic and ter-restrial habitats. Annu. Rev. Microbiol. 1985, 39, 321–346. [Google Scholar] [CrossRef] [PubMed]
- Bardgett, R.D.; van der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef]
- Tiedje, J.M.; Asuming-Brempong, S.; Nüsslein, K.; Marsh, T.L.; Flynn, S.J. Opening the black box of soil microbial di-versity. Appl. Soil Ecol. 1999, 13, 109–122. [Google Scholar] [CrossRef]
- Sokol, N.W.; Slessarev, E.; Marschmann, G.L.; Nicolas, A.; Blazewicz, S.J.; Brodie, E.L.; Firestone, M.K.; Foley, M.M.; Hestrin, R.; Hungate, B.A.; et al. Life and death in the soil microbiome: How ecological processes influence biogeo-chemistry. Nat. Rev. Microbiol. 2022, 20, 415–430. [Google Scholar] [CrossRef]
- Andrews, J.H.; Harris, R.F. R- and K-Selection and Microbial Ecology BT—Advances in Microbial Ecology; Marshall, K.C., Ed.; Springer: Boston, MA, USA, 1986; pp. 99–147. ISBN 978-1-4757-0611-6. [Google Scholar]
- Kielak, A.M.; Cipriano, M.A.P.; Kuramae, E.E. Acidobacteria strains from subdivision 1 act as plant growth-promoting bacteria. Arch. Microbiol. 2016, 198, 987–993. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.S.; Shen, F.T. Bioprospecting of facultatively oligotrophic bacteria from non-rhizospheric soils. Appl. Soil Ecol. 2016, 108, 315–324. [Google Scholar] [CrossRef]
- Joseph, S.J.; Hugenholtz, P.; Sangwan, P.; Osborne, C.A.; Janssen, P.H. Laboratory cultivation of widespread and previously uncultured soil bacteria. Appl. Environ. Microbiol. 2003, 69, 7210–7215. [Google Scholar] [CrossRef]
- Kato, S.; Yamagishi, A.; Daimon, S.; Kawasaki, K.; Tamaki, H.; Kitagawa, W.; Abe, A.; Tanaka, M.; Sone, T.; Asano, K.; et al. Isolation of previously uncultured slow-growing bacteria by using a simple modification in the preparation of agar media. Appl. Environ. Microbiol. 2018, 84, e00807-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pulschen, A.A.; Bendia, A.G.; Fricker, A.D.; Pellizari, V.H.; Galante, D.; Rodrigues, F. Isolation of uncultured bacteria from antarctica using long incubation periods and low nutritional media. Front. Microbiol. 2017, 8, 1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tschech, A.; Pfennig, N. Growth yield increase linked to caffeate reduction in Acetobacterium Woodii. Arch. Microbiol. 1984, 137, 163–167. [Google Scholar] [CrossRef]
- Widdel, F.; Kohring, G.-W.; Mayer, F. Studies on dissimilatory sulfate-reducing bacteria that decompose fatty acids. Arch. Microbiol. 1983, 134, 286–294. [Google Scholar] [CrossRef]
- Koeuth, T.; Versalovic, J.; Lupski, J.R. Differential subsequence conservation of interspersed repetitive Streptococcus Pneumoniae BOX elements in diverse bacteria. Genome Res. 1995, 5, 408–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Versalovic, J.; Koeuth, T.; Lupski, J.R. Distribution of repetitive dna sequences in eubacteria and application to fin-gerprinting of bacterial genomes. Nucleic Acids Res. 1991, 19, 6823–6831. [Google Scholar] [CrossRef]
- Heuer, H.; Krsek, M.; Baker, P.; Smalla, K.; Wellington, E.M. Analysis of Actinomycete communities by specific am-plification of genes encoding 16S rRNA and gel-electrophoretic separation in denaturing gradients. Appl. Environ. Microbiol. 1997, 63, 3233–3241. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT Online Service: Multiple Sequence Alignment, Interactive Sequence Choice and Visualization. Brief Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Kavamura, V.N.; Santos, S.N.; da Silva, J.L.; Parma, M.M.; Ávila, L.A.; Visconti, A.; Zucchi, T.D.; Taketani, R.G.; Andreote, F.D.; Melo, I.S. de Screening of Brazilian cacti rhizobacteria for plant growth promotion under drought. Microbiol Res 2013, 168, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Paulo, E.M.; Vasconcelos, M.P.; Oliveira, I.S.; de Affe, H.M.J.; Nascimento, R.; de Melo, I.S.; de Roque, M.R.A.; de Assis, S.A. An Alternative method for screening lactic acid bacteria for the production of exopolysaccharides with rapid confirmation. Food Sci. Technol. 2012, 32, 710–714. [Google Scholar] [CrossRef] [Green Version]
- Gordon, S.A.; Weber, R.P. Colorimetric estimation of indoleacetic acid. Plant Physiol. 1951, 26, 192–195. [Google Scholar] [CrossRef] [Green Version]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing Microorganisms. FEMS Microbiol Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Chan, Y.; Chen, Y.; Shi, C.; Huang, Z.; Yong, Z.; Shengkang, L.; Li, Y.; Ye, J.; Yu, C.; Li, Z.; et al. SOAPnuke: A mapreduce acceleration supported software for integrated quality control and preprocessing of high-throughput se-quencing data. Gigascience 2017, 7, gix120. [Google Scholar] [CrossRef] [Green Version]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinfor-matics. 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [Green Version]
- Parks, D.; Imelfort, M.; Skennerton, C.; Philip, H.; Tyson, G. CheckM: Assessing the Quality of Microbial Genomes Recovered from Isolates, Single Cells, and Metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier-Kolthoff, J.P.; Göker, M. TYGS Is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patz, S.; Gautam, A.; Becker, M.; Ruppel, S.; Rodríguez-Palenzuela, P.; Huson, D.H. PLaBAse: A comprehensive web resource for analyzing the plant growth-promoting potential of plant-associated bacteria. bioRxiv 2021. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. AntiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [Green Version]
- Navarro-Munoz, J.; Selem, N.; Mullowney, M.; Kautsar, S.; Tryon, J.; Parkinson, E.; de los Santos, E.L.; Yeong, M.; Cruz-Morales, P.; Abubucker, S.; et al. A computational framework for systematic exploration of biosynthetic diversity from large-scale genomic data. Nat. Chem. Biol. 2020, 16, 60–68. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cy-toscape: A software environment for integrated models of biomolecular interaction networks. Genome Res 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Abawi, G.S.; Pastor Corrales, M.A.; Centro Internacional de Agricultura Tropical. Root Rots of Beans in Latin America and Africa: Diagnosis, Research Methodologies, and Management Strategies; Centro Internacional de Agricultura Tropical: Palmira, Colombian, 1990; ISBN 958918314X. [Google Scholar]
- Fenta, B.A.; Beebe, S.E.; Kunert, K.J.; Burridge, J.D.; Barlow, K.M.; Lynch, J.P.; Foyer, C.H. Field phenotyping of soy-bean roots for drought stress tolerance. Agronomy 2014, 4, 418–435. [Google Scholar] [CrossRef] [Green Version]
- Glick, B.R. Plant Growth-Promoting Bacteria: Mechanisms and Applications. Scientifica (Cairo) 2012, 2012, 963401. [Google Scholar] [CrossRef] [Green Version]
- Patten, C.L.; Glick, B.R. Bacterial Biosynthesis of Indole-3-Acetic Acid. Can. J. Microbiol. 1996, 42, 207–220. [Google Scholar] [CrossRef]
- Ehrlich, H.L. Geomicrobiology: Its Significance for Geology. Earth Sci. Rev. 1998, 45, 45–60. [Google Scholar] [CrossRef]
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil Beneficial Bacteria and Their Role in Plant Growth Promotion: A Review. Ann. Microbiol. 2010, 60, 579–598. [Google Scholar] [CrossRef]
- Yang, X.; Post, W.M.; Thornton, P.E.; Jain, A. The Distribution of Soil Phosphorus for Global Biogeochemical Modeling. Biogeosciences 2013, 10, 2525–2537. [Google Scholar] [CrossRef] [Green Version]
- Kuklinsky-Sobral, J.; Araújo, W.L.; Mendes, R.; Geraldi, I.O.; Pizzirani-Kleiner, A.A.; Azevedo, J.L. Isolation and characterization of soybean-associated bacteria and their potential for plant growth promotion. Environ. Microbiol. 2004, 6, 1244–1251. [Google Scholar] [CrossRef] [PubMed]
- Moretti, L.G.; Crusciol, C.A.C.; Kuramae, E.E.; Bossolani, J.W.; Moreira, A.; Costa, N.R.; Alves, C.J.; Pascoaloto, I.M.; Rondina, A.B.L.; Hungria, M. Effects of Growth-Promoting Bacteria on Soybean Root Activity, Plant Development, and Yield. Agron. J. 2020, 112, 418–428. [Google Scholar] [CrossRef]
- Schmidt, J.; Messmer, M.; Wilbois, K.-P. Beneficial Microorganisms for Soybean (Glycine max (L.) Merr), with a Focus on Low Root-Zone Temperatures. Plant Soil 2015, 397, 411–445. [Google Scholar] [CrossRef]
- Sait, M.; Hugenholtz, P.; Janssen, P.H. Cultivation of Globally Distributed Soil Bacteria from Phylogenetic Lineages Previously Only Detected in Cultivation-Independent Surveys. Environ. Microbiol. 2002, 4, 654–666. [Google Scholar] [CrossRef]
- Madhaiyan, M.; Poonguzhali, S.; Lee, J.-S.; Lee, K.-C.; Saravanan, V.S.; Santhanakrishnan, P. Microbacterium aza-dirachtae Sp. Nov., a Plant-Growth-Promoting Actinobacterium Isolated from the Rhizoplane of Neem Seedlings. Int. J. Syst. Evol. Microbiol. 2010, 60, 1687–1692. [Google Scholar] [CrossRef] [Green Version]
- Dourado, M.N.; Bogas, A.C.; Pomini, A.M.; Andreote, F.D.; Quecine, M.C.; Marsaioli, A.J.; Araújo, W.L. Methylobac-terium-Plant Interaction Genes Regulated by Plant Exudate and Quorum Sensing Molecules. Braz. J. Microbiol. 2013, 44, 1331–1339. [Google Scholar] [CrossRef]
- Grossi, C.E.M.; Fantino, E.; Serral, F.; Zawoznik, M.S.; Fernandez Do Porto, D.A.; Ulloa, R.M. Methylobacterium Sp. 2A Is a Plant Growth-Promoting Rhizobacteria That Has the Potential to Improve Potato Crop Yield Under Adverse Conditions. Front. Plant Sci. 2020, 11, 71. [Google Scholar] [CrossRef]
- Liu, Z.-P.; Wang, B.-J.; Liu, Y.-H.; Liu, S.-J. Novosphingobium Taihuense sp. nov., a Novel Aro-matic-Compound-Degrading Bacterium Isolated from Taihu Lake, China. Int. J. Syst. Evol. Microbiol. 2005, 55, 1229–1232. [Google Scholar] [CrossRef]
- Sohn, J.H.; Kwon, K.K.; Kang, J.-H.; Jung, H.-B.; Kim, S.-J. Novosphingobium Pentaromativorans sp. nov., a High-Molecular-Mass Polycyclic Aromatic Hydrocarbon-Degrading Bacterium Isolated from Estuarine Sediment. Int. J. Syst. Evol. Microbiol. 2004, 54, 1483–1487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westerberg, K.; Elväng, A.M.; Stackebrandt, E.; Jansson, J.K. Arthrobacter chlorophenolicus sp. nov., a New Species Capable of Degrading High Concentrations of 4-Chlorophenol. Int. J. Syst. Evol. Microbiol. 2000, 50, 2083–2092. [Google Scholar] [CrossRef] [PubMed]
- Salwan, R.; Sharma, V. Molecular and biotechnological aspects of secondary metabolites in actinobacteria. Microbiol. Res. 2020, 231, 126374. [Google Scholar] [CrossRef] [PubMed]
- Bruto, M.; Prigent-Combaret, C.; Muller, D.; Moënne-Loccoz, Y. Analysis of Genes Contributing to Plant-Beneficial Functions in Plant Growth-Promoting Rhizobacteria and Related Proteobacteria. Sci. Rep. 2014, 4, 6261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, V.; Bruto, M.; Bellvert, F.; Bally, R.; Muller, D.; Prigent-Combaret, C.; Moënne-Loccoz, Y.; Comte, G. Unex-pected Phytostimulatory Behavior for Escherichia coli and Agrobacterium Tumefaciens Model Strains. Mol. Plant Microbe Interact. 2013, 26, 495–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imam, J.; Singh, P.K.; Shukla, P. Plant Microbe Interactions in Post Genomic Era: Perspectives and Applications. Front. Microbiol. 2016, 7, 1488. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, H.; Bagyaraj, D.J.; Selvakumar, G.; Sundaram, S.P. Novel Plant Growth Promoting Rhizobacte-ria—Prospects and Potential. Appl. Soil Ecol. 2015, 95, 38–53. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonçalves, O.S.; Souza, T.S.; Gonçalves, G.d.C.; Fernandes, A.S.; Veloso, T.G.R.; Tupy, S.M.; Garcia, E.A.; Santana, M.F. Harnessing Novel Soil Bacteria for Beneficial Interactions with Soybean. Microorganisms 2023, 11, 300. https://doi.org/10.3390/microorganisms11020300
Gonçalves OS, Souza TS, Gonçalves GdC, Fernandes AS, Veloso TGR, Tupy SM, Garcia EA, Santana MF. Harnessing Novel Soil Bacteria for Beneficial Interactions with Soybean. Microorganisms. 2023; 11(2):300. https://doi.org/10.3390/microorganisms11020300
Chicago/Turabian StyleGonçalves, Osiel Silva, Thamires Santos Souza, Guilherme de Castro Gonçalves, Alexia Suellen Fernandes, Tomás Gomes Reis Veloso, Sumaya Martins Tupy, Ediones Amaro Garcia, and Mateus Ferreira Santana. 2023. "Harnessing Novel Soil Bacteria for Beneficial Interactions with Soybean" Microorganisms 11, no. 2: 300. https://doi.org/10.3390/microorganisms11020300
APA StyleGonçalves, O. S., Souza, T. S., Gonçalves, G. d. C., Fernandes, A. S., Veloso, T. G. R., Tupy, S. M., Garcia, E. A., & Santana, M. F. (2023). Harnessing Novel Soil Bacteria for Beneficial Interactions with Soybean. Microorganisms, 11(2), 300. https://doi.org/10.3390/microorganisms11020300