
Draft Genome of Nocardia canadensis sp. nov. Isolated from Petroleum-Hydrocarbon-Contaminated Soil
 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Media Culture of Bacteria
2.2. DNA Extraction and Whole-Genome Shot Gun Sequencing
2.3. Bioinformatics Pipeline and Processing of Data
2.4. Description of Nocardia canadensis sp. nov.
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Alotaibi, F.; Hijri, M.; St-Arnaud, M. Overview of Approaches to Improve Rhizoremediation of Petroleum Hydrocarbon-Contaminated Soils. Appl. Microbiol. 2021, 1, 329–351. [Google Scholar] [CrossRef]
- Alotaibi, F.; Lee, S.-J.; St-Arnaud, M.; Hijri, M. Salix purpurea and Eleocharis obtusa rhizospheres harbor a diverse rhizospheric bacterial community characterized by hydrocarbons degradation potentials and plant growth-promoting properties. Plants 2021, 10, 1987. [Google Scholar] [CrossRef] [PubMed]
- Gkorezis, P.; Daghio, M.; Franzetti, A.; Van Hamme, J.D.; Sillen, W.; Vangronsveld, J. The Interaction between Plants and Bacteria in the Remediation of Petroleum Hydrocarbons: An Environmental Perspective. Front. Microbiol. 2016, 7, 1836. [Google Scholar] [CrossRef] [PubMed]
- Pandey, P.; Pathak, H.; Dave, S. Microbial ecology of hydrocarbon degradation in the soil: A review. Res. J. Environ. Toxicol. 2016, 10, 1–15. [Google Scholar] [CrossRef]
- Pilon-Smits, E.A.; Freeman, J.L. Environmental cleanup using plants: Biotechnological advances and ecological considerations. Front. Ecol. Environ. 2006, 4, 203–210. [Google Scholar] [CrossRef]
- Khan, F.I.; Husain, T.; Hejazi, R. An overview and analysis of site remediation technologies. J. Environ. Manag. 2004, 71, 95–122. [Google Scholar] [CrossRef] [PubMed]
- Pilon-Smits, E. Phytoremediation. Annu. Rev. Plant Biol. 2005, 56, 15–39. [Google Scholar] [CrossRef]
- Marchand, C.; Mench, M.; Jani, Y.; Kaczala, F.; Notini, P.; Hijri, M.; Hogland, W. Pilot scale aided-phytoremediation of a co-contaminated soil. Sci. Total Environ. 2018, 618, 753–764. [Google Scholar] [CrossRef]
- Yergeau, E.; Bell, T.H.; Champagne, J.; Maynard, C.; Tardif, S.; Tremblay, J.; Greer, C.W. Transplanting soil microbiomes leads to lasting effects on willow growth, but not on the rhizosphere microbiome. Front. Microbiol. 2015, 6, 1436. [Google Scholar] [CrossRef]
- Kuzovkina, Y.A.; Volk, T.A. The characterization of willow (Salix L.) varieties for use in ecological engineering applications: Co-ordination of structure, function and autecology. Ecol. Eng. 2009, 35, 1178–1189. [Google Scholar] [CrossRef]
- Alotaibi, F.; St-Arnaud, M.; Hijri, M. In-Depth Characterization of Plant Growth Promotion Potentials of Selected Alkanes-Degrading Plant Growth-Promoting Bacterial Isolates. Front. Microbiol. 2022, 13, 863702. [Google Scholar] [CrossRef] [PubMed]
- Zhi, X.-Y.; Li, W.-J.; Stackebrandt, E. An update of the structure and 16S rRNA gene sequence-based definition of higher ranks of the class Actinobacteria, with the proposal of two new suborders and four new families and emended descriptions of the existing higher taxa. Int. J. Syst. Evol. Microbiol. 2009, 59, 589–608. [Google Scholar] [CrossRef] [PubMed]
- Brown-Elliott, B.A.; Brown, J.M.; Conville, P.S.; Wallace, R.J., Jr. Clinical and laboratory features of the Nocardia spp. based on current molecular taxonomy. Clin. Microbiol. Rev. 2006, 19, 259–282. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Hiessl, S.; Steinbüchel, A. Functional diversity of Nocardia in metabolism. Environ. Microbiol. 2014, 16, 29–48. [Google Scholar] [CrossRef] [PubMed]
- Mehta, H.H.; Shamoo, Y. Pathogenic Nocardia: A diverse genus of emerging pathogens or just poorly recognized? PLoS Pathog. 2020, 16, e1008280. [Google Scholar] [CrossRef] [PubMed]
- Nocard, E. Note sur la maladie des boeufs de la Guadeloupe connue sous le nom de farcin. Ann. L’Inst Pasteur 1888, 2, 293–302. [Google Scholar]
- Beaman, B.L.; Beaman, L. Nocardia species: Host-parasite relationships. Clin. Microbiol. Rev. 1994, 7, 213–264. [Google Scholar] [CrossRef]
- Vautrin, F.; Bergeron, E.; Dubost, A.; Abrouk, D.; Martin, C.; Cournoyer, B.; Louzier, V.; Winiarski, T.; Rodriguez-Nava, V.; Pujic, P. Genome Sequences of Three Nocardia cyriacigeorgica Strains and One Nocardia asteroides Strain. Microbiol. Resour. Announ. 2019, 8, e00600-19. [Google Scholar] [CrossRef]
- Schneider, K.; Rose, I.; Vikineswary, S.; Jones, A.L.; Goodfellow, M.; Nicholson, G.; Beil, W.; Süssmuth, R.D.; Fiedler, H.-P. Nocardichelins A and B, Siderophores from Nocardia Strain Acta 3026. J. Nat. Prod. 2007, 70, 932–935. [Google Scholar] [CrossRef]
- El-Gendy, M.M.; Hawas, U.W.; Jaspars, M. Novel bioactive metabolites from a marine derived Bacterium nocardia sp. ALAA 2000. J. Antibiot. 2008, 61, 379–386. [Google Scholar] [CrossRef]
- Brzeszcz, J.; Kaszycki, P. Aerobic bacteria degrading both n-alkanes and aromatic hydrocarbons: An undervalued strategy for metabolic diversity and flexibility. Biodegradation 2018, 29, 359–407. [Google Scholar] [CrossRef] [PubMed]
- Bell, T.H.; El-Din Hassan, S.E.; Lauron-Moreau, A.; Al-Otaibi, F.; Hijri, M.; Yergeau, E.; St-Arnaud, M. Linkage between bacterial and fungal rhizosphere communities in hydrocarbon-contaminated soils is related to plant phylogeny. ISME J. 2014, 8, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Yergeau, E.; Sanschagrin, S.; Maynard, C.; St-Arnaud, M.; Greer, C.W. Microbial expression profiles in the rhizosphere of willows depend on soil contamination. ISME J. 2014, 8, 344–358. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [PubMed]
- Krichevsky, M.; Moore, L.; Moore, W.; Murray, R.; Stackebrandt, E.; Starr, M.; Trper, H. International Committee on Systematic Bacteriology. Report of the ad hoc committee on reconciliation of approaches to bacterial systematics. Int. J. Syst. Bacteriol. 1987, 37, 463464. [Google Scholar] [CrossRef]
- Kim, M.; Oh, H.-S.; Park, S.-C.; Chun, J. Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 346–351. [Google Scholar] [CrossRef]
- Yoon, S.-H.; Ha, S.-M.; Lim, J.; Kwon, S.; Chun, J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Antonie Van Leeuwenhoek 2017, 110, 1281–1286. [Google Scholar] [CrossRef]
- Stackebrandt, E. Taxonomic parameters revisited: Tarnished gold standards. Microbiol. Today 2006, 33, 152–155. [Google Scholar]
- Tindall, B.J.; Rosselló-Móra, R.; Busse, H.-J.; Ludwig, W.; Kämpfer, P. Notes on the characterization of prokaryote strains for taxonomic purposes. Int. J. Syst. Evol. Microbiol. 2010, 60, 249–266. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kühnert, D.; De Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Fadeev, E.; Cardozo-Mino, M.G.; Rapp, J.Z.; Bienhold, C.; Salter, I.; Salman-Carvalho, V.; Molari, M.; Tegetmeyer, H.E.; Buttigieg, P.L.; Boetius, A. Comparison of two 16S rRNA primers (V3–V4 and V4–V5) for studies of arctic microbial communities. Front. Microbiol. 2021, 12, 283. [Google Scholar] [CrossRef]
- Bukin, Y.S.; Galachyants, Y.P.; Morozov, I.; Bukin, S.; Zakharenko, A.; Zemskaya, T. The effect of 16S rRNA region choice on bacterial community metabarcoding results. Sci. Data 2019, 6, 190007. [Google Scholar] [CrossRef]
- Roth, A.; Andrees, S.; Kroppenstedt, R.M.; Harmsen, D.; Mauch, H. Phylogeny of the genus Nocardia based on reassessed 16S rRNA gene sequences reveals underspeciation and division of strains classified as Nocardia asteroides into three established species and two unnamed taxons. J. Clin. Microbiol. 2003, 41, 851–856. [Google Scholar] [CrossRef]
- Ji, Y.; Mao, G.; Wang, Y.; Bartlam, M. Structural insights into diversity and n-alkane biodegradation mechanisms of alkane hydroxylases. Front. Microbiol. 2013, 4, 58. [Google Scholar] [CrossRef]
- Nie, Y.; Chi, C.-Q.; Fang, H.; Liang, J.-L.; Lu, S.-L.; Lai, G.-L.; Tang, Y.-Q.; Wu, X.-L. Diverse alkane hydroxylase genes in microorganisms and environments. Sci. Rep. 2014, 4, 4968. [Google Scholar] [CrossRef]
- Nie, Y.; Liang, J.-L.; Fang, H.; Tang, Y.-Q.; Wu, X.-L. Characterization of a CYP153 alkane hydroxylase gene in a Gram-positive Dietzia sp. DQ12-45-1b and its “team role” with alkW1 in alkane degradation. Appl. Microbiol. Biotechnol. 2014, 98, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Peng, R.-H.; Xiong, A.-S.; Xue, Y.; Fu, X.-Y.; Gao, F.; Zhao, W.; Tian, Y.-S.; Yao, Q.-H. Microbial biodegradation of polyaromatic hydrocarbons. FEMS Microbiol. Rev. 2008, 32, 927–955. [Google Scholar] [CrossRef] [PubMed]
- Duff, S.M.G.; Sarath, G.; Plaxton, W. The role of acid phosphatases in plant phosphorus metabolism. Physiol. Plant. 1994, 90, 791–800. [Google Scholar] [CrossRef]
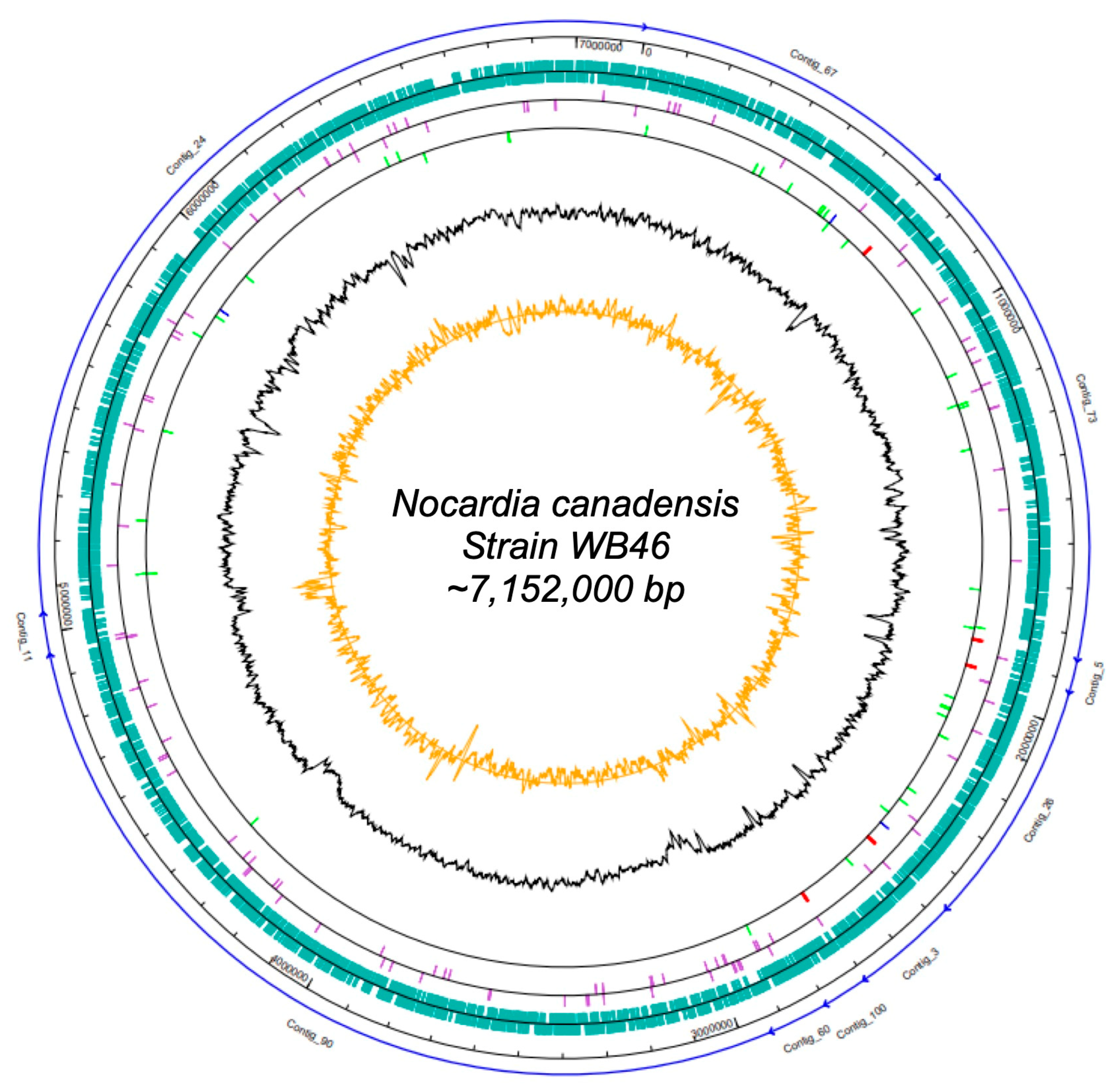



{kind=link}
{kind=link}
{kind=link}
| Traits | Assays | Activity |
|---|---|---|
| Hydrocarbon degradation potential b | Naphthalene | ++ |
| Phenanthrene | +++ | |
| Pyrene | ++ | |
| Dodecane | ++ | |
| Hexadecane | +++ | |
| Catabolic genes c | Alkane monooxygenase (alkB) | + |
| Cytochrome P450 hydroxylase (CYP153) | − | |
| Naphthalene dioxygenase (nah1) | + | |
| Cell growth measurement at 600 nm b | 1% diesel | ++++ |
| 1% hexadecane | ++++ | |
| 2% hexadecane | ++++ | |
| 3% hexadecane | ++++ | |
| Plant-growth-promoting traits | 1-Aminocyclopropane-1-carboxylate deaminase (ACCD) | − |
| Phosphate solubilization | − | |
| Siderophore production | + (8.2%) | |
| Nitrogen fixation | − | |
| Indole-3-acetic (IAA) production | + (1.46 μg mL−1) | |
| Ammonia production | + (2.9 μmol mL−1) | |
| Root elongation assay (cm) d | 0% | 13.4 |
| 1% | 12.2 | |
| 2% | 10.1 | |
| 3% | 8.6 |
| Analysis | Value | Comments |
|---|---|---|
| In silico DNA–DNA hybridization (DDH) | 63.4% | Below the threshold level of 70% |
| Type (Strain) Genome Server (TYGS) | No match | Tagged as novel species |
| Average nucleotide identity (ANI) | 88.63% | Below the species boundary cut-off of 95–96% |
| 16S rRNA gene sequence (RDP) | 98.8% | High similarity to Nocardia asteroides |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alotaibi, F.; Lee, S.-J.; Lahrach, Z.; St-Arnaud, M.; Hijri, M. Draft Genome of Nocardia canadensis sp. nov. Isolated from Petroleum-Hydrocarbon-Contaminated Soil. Microorganisms 2023, 11, 2972. https://doi.org/10.3390/microorganisms11122972
Alotaibi F, Lee S-J, Lahrach Z, St-Arnaud M, Hijri M. Draft Genome of Nocardia canadensis sp. nov. Isolated from Petroleum-Hydrocarbon-Contaminated Soil. Microorganisms. 2023; 11(12):2972. https://doi.org/10.3390/microorganisms11122972
Chicago/Turabian StyleAlotaibi, Fahad, Soon-Jae Lee, Zakaria Lahrach, Marc St-Arnaud, and Mohamed Hijri. 2023. "Draft Genome of Nocardia canadensis sp. nov. Isolated from Petroleum-Hydrocarbon-Contaminated Soil" Microorganisms 11, no. 12: 2972. https://doi.org/10.3390/microorganisms11122972
APA StyleAlotaibi, F., Lee, S.-J., Lahrach, Z., St-Arnaud, M., & Hijri, M. (2023). Draft Genome of Nocardia canadensis sp. nov. Isolated from Petroleum-Hydrocarbon-Contaminated Soil. Microorganisms, 11(12), 2972. https://doi.org/10.3390/microorganisms11122972