
Analysis of Phenotypic and Genotypic Susceptibility to Clarithromycin and Amikacin of Mycobacterium abscessus Complex Strains Isolated from Cystic Fibrosis Patients


Abstract
:1. Introduction
2. Materials and Methods
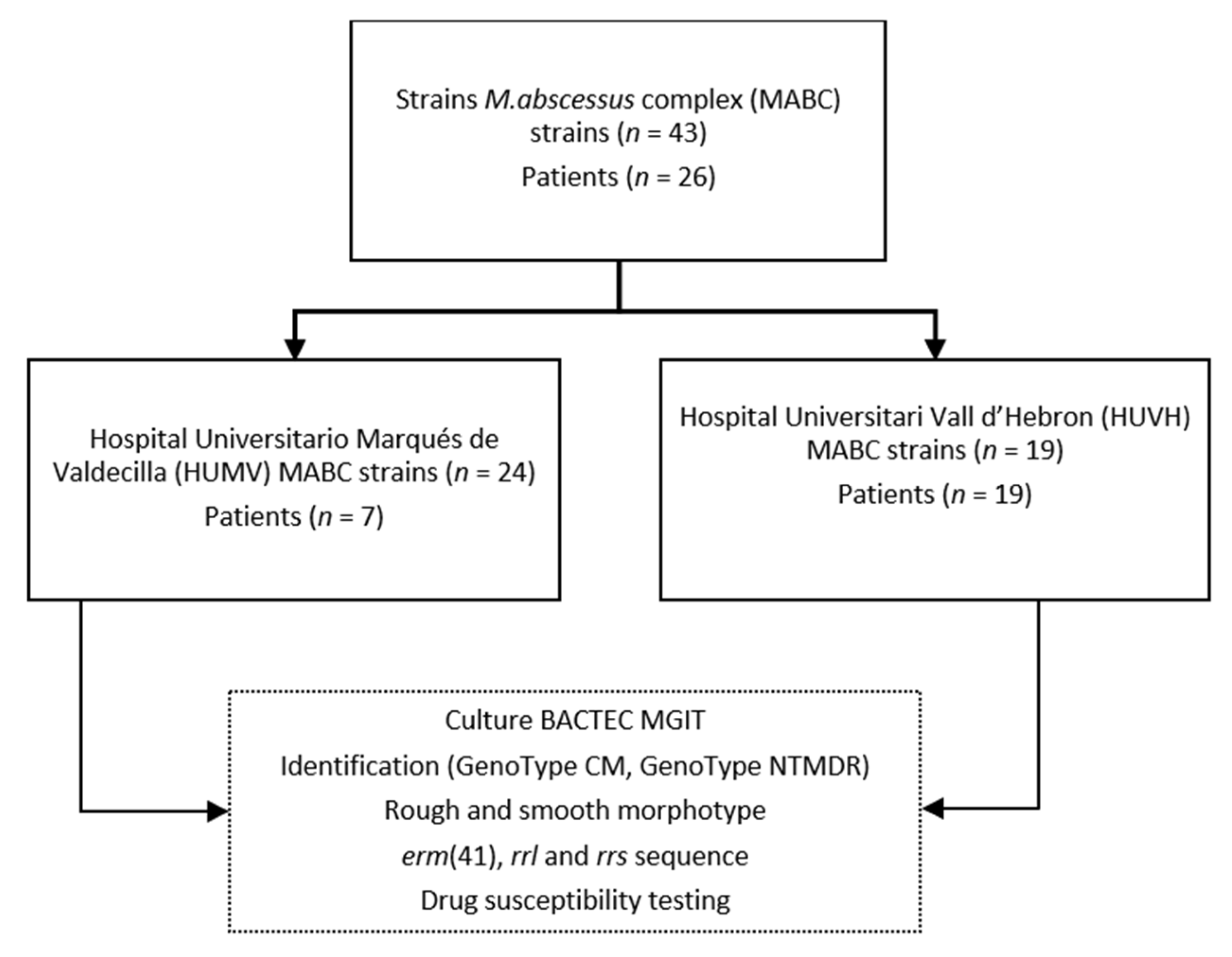
2.1. Strains
2.2. Isolation and Identification
2.3. Smooth and Rough Morphotypes
2.4. Susceptibility Testing
2.5. erm(41), rrl, and rrs PCR for Sequencing
3. Results
3.1. Identification of Isolates
3.2. Smooth and Rough Morphotypes
3.3. Genotyping of the erm and rrl Genes and Susceptibility Testing to Clarithromycin
3.4. Genotyping of the rrs Gene and Susceptibility Testing to Amikacin
3.5. Phenotypic Antibiotic Susceptibility Testing
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Edwards, Q.T.; Seibert, D.; Macri, C.; Covington, C.; Tilghman, J. Assessing Ethnicity in Preconception Counseling: Genetics—What Nurse Practitioners Need to Know. J. Am. Acad. Nurse Pract. 2004, 16, 472–480. [Google Scholar] [CrossRef] [PubMed]
- Elborn, J.S. Cystic Fibrosis. Lancet 2016, 388, 2519–2531. [Google Scholar] [CrossRef] [PubMed]
- Kiedrowski, M.R.; Bomberger, J.M. Viral-Bacterial Co-Infections in the Cystic Fibrosis Respiratory Tract. Front. Immunol. 2018, 9, 3067. [Google Scholar] [CrossRef]
- Strnad, L.; Winthrop, K.L. Treatment of Mycobacterium abscessus Complex. Semin. Respir. Crit. Care Med. 2018, 39, 362–376. [Google Scholar]
- Janahi, I.A.; Rehman, A. The Cystic Fibrosis Airway Microbiome and Pathogens. In Progress in Understanding Cystic Fibrosis; Sriramulu, D., Ed.; InTech: Rijeka, Croatia, 2017; pp. 45–71. [Google Scholar]
- Koh, W.; Jeon, K.; Lee, N.Y.; Kim, B.; Kook, Y.; Lee, S.; Park, Y.K.; Kim, C.K.; Shin, S.J.; Huitt, G.A.; et al. Clinical Significance of Differentiation of Mycobacterium massiliense from Mycobacterium abscessus. Am. J. Respir. Crit. Care Med. 2011, 183, 405–410. [Google Scholar] [CrossRef]
- Daley, C.L.; Iaccarino, J.M.; Lange, C.; Cambau, E.; Wallace, R.J., Jr.; Andrejak, C.; Böttger, E.C.; Brozek, J.; Griffith, D.E.; Guglielmetti, L.; et al. Treatment of Nontuberculous Mycobacterial Pulmonary Disease: An Official ATS/ERS/ESCMID/IDSA Clinical Practice Guideline. Eur. Respir. J. 2020, 56, 2000535. [Google Scholar] [CrossRef]
- Brown-Elliott, B.A.; Woods, G.L. Antimycobacterial Susceptibility Testing of Nontuberculous Mycobacteria. J. Clin. Microbiol. 2019, 57, e00834-19. [Google Scholar] [CrossRef]
- Bastian, S.; Veziris, N.; Roux, A.L.; Brossier, F.; Gaillard, J.L.; Jarlier, V.; Cambau, E. Assessment of Clarithromycin Susceptibility in Strains Belonging to the Mycobacterium abscessus Group by erm(41) and rrl Sequencing. Antimicrob. Agents Chemother. 2011, 55, 775–781. [Google Scholar] [CrossRef]
- Nash, K.A.; Brown-Elliott, A.B.; Wallace, R.J. A Novel Gene, erm(41) Confers Inducible Macrolide Resistance to Clinical Isolates of Mycobacterium abscessus but Is Absent from Mycobacterium chelonae. Antimicrob. Agents Chemother. 2009, 53, 1367–1376. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Kim, B.J.; Kook, Y.; Yun, Y.J.; Shin, J.H.; Kim, B.J.; Kook, Y.H. Mycobacterium massiliense Is Differentiated from Mycobacterium abscessus and Mycobacterium bolletii by Erythromycin Ribosome Methyltransferase Gene (erm) and Clarithromycin Susceptibility Patterns. Microbiol. Immunol. 2010, 54, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Prammananan, T.; Sander, P.; Brown, B.A.; Frischkorn, K.; Onyi, G.O.; Zhang, Y.; Böttger, E.C.; Wallace, R.J. A Single 16S Ribosomal RNA Substitution Is Responsible for Resistance to Amikacin and Other 2-Deoxystreptamine Aminoglycosides in Mycobacterium abscessus and Mycobacterium chelonae. J. Infect. Dis. 1998, 177, 1573–1581. [Google Scholar] [CrossRef] [PubMed]
- Johansen, M.D.; Herrmann, J.-L.; Kremer, L. Non-Tuberculous Mycobacteria and the Rise of Mycobacterium abscessus. Nat. Rev. Microbiol. 2020, 18, 392–407. [Google Scholar] [CrossRef] [PubMed]
- Ryan, K.; Byrd, T.F. Mycobacterium abscessus: Shapeshifter of the Mycobacterial World. Front. Microbiol. 2018, 9, 2642. [Google Scholar] [CrossRef]
- Ridell, M. Mycobacterium abscessus: An Environmental Mycobacteria Being a Human Pathogen. Int. J. Mycobacteriol. 2015, 4, 41. [Google Scholar] [CrossRef]
- Catherinot, E.; Roux, A.L.; Macheras, E.; Hubert, D.; Matmar, M.; Dannhoffer, L.; Chinet, T.; Morand, P.; Poyart, C.; Heym, B.; et al. Acute Respiratory Failure Involving an R Variant of Mycobacterium abscessus. J. Clin. Microbiol. 2009, 47, 271–274. [Google Scholar] [CrossRef]
- Griffith, D.E.; Aksamit, T.; Brown-Elliott, B.A.; Catanzaro, A.; Daley, C.; Gordin, F.; Holland, S.M.; Horsburgh, R.; Huitt, G.; Iademarco, M.F.; et al. An Official ATS/IDSA Statement: Diagnosis, Treatment, and Prevention of Nontuberculous Mycobacterial Diseases. Am. J. Respir. Crit. Care Med. 2007, 175, 367–416. [Google Scholar] [CrossRef] [PubMed]
- Woods, G.L. Susceptibility Testing of Mycobacteria, Nocardiae, and Other Aerobic Actinomycetes, Approved Standard, 2nd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2011; ISBN 1562387464. [Google Scholar]
- Meier, A.; Kirschner, P.; Springer, B.; Steingrube, V.A.; Brown, B.A.; Wallace, R.J.; Bottger, E.C. Identification of Mutations in 23S RRNA Gene of Clarithromycin-Resistant Mycobacterium intracellulare. Antimicrob. Agents Chemother. 1994, 38, 381–384. [Google Scholar] [CrossRef]
- Maiwald, M. Broad-Range PCR for Detection and Identification of Bacteria. In Molecular Microbiology: Diagnostic Principles and Practice; Persing, D.H., Tenover, F.C., Versalovic, J., Tang, Y.W., Uger, E.R., Relman, D., White, T.J., Eds.; ASM Press: Washington, DC, USA, 2004; pp. 379–390. [Google Scholar]
- Macheras, E.; Roux, A.L.; Ripoll, F.; Sivadon-Tardy, V.; Gutierrez, C.; Gaillard, J.L.; Heym, B. Inaccuracy of Single-Target Sequencing for Discriminating Species of the Mycobacterium abscessus Group. J. Clin. Microbiol. 2009, 47, 2596–2600. [Google Scholar] [CrossRef]
- Nakanaga, K.; Sekizuka, T.; Fukano, H.; Sakakibara, Y.; Takeuchi, F.; Wada, S.; Ishii, N.; Makino, M.; Kuroda, M.; Hoshino, Y. Discrimination of Mycobacterium abscessus subsp. massiliense from Mycobacterium abscessus subsp. abscessus in Clinical Isolates by Multiplex PCR. J. Clin. Microbiol. 2014, 52, 251–259. [Google Scholar] [CrossRef]
- Zelazny, A.M.; Root, J.M.; Shea, Y.R.; Colombo, R.E.; Shamputa, I.C.; Stock, F.; Conlan, S.; McNulty, S.; Brown-Elliott, B.A.; Wallace, R.J.; et al. Cohort Study of Molecular Identification and Typing of Mycobacterium abscessus, Mycobacterium massiliense, and Mycobacterium bolletii. J. Clin. Microbiol. 2009, 47, 1985–1995. [Google Scholar] [CrossRef]
- Huh, H.J.; Kim, S.Y.; Shim, H.J.; Kim, D.H.; Yoo, I.Y.; Kang, O.K.; Ki, C.S.; Shin, S.Y.; Jhun, B.W.; Shin, S.J.; et al. GenoType NTM-DR Performance Evaluation for Identification of Mycobacterium avium Complex and Mycobacterium abscessus and Determination of Clarithromycin and Amikacin Resistance. J. Clin. Microbiol. 2019, 57, e00516-19. [Google Scholar] [CrossRef] [PubMed]
- Kehrmann, J.; Kurt, N.; Rueger, K.; Bange, F.-C.; Buer, J. GenoType NTM-DR for Identifying Mycobacterium abscessus Subspecies and Determining Molecular Resistance. J. Clin. Microbiol. 2016, 54, 1653–1655. [Google Scholar] [CrossRef] [PubMed]
- Rubio, M.; March, F.; Garrigó, M.; Moreno, C.; Español, M.; Coll, P. Inducible and Acquired Clarithromycin Resistance in the Mycobacterium abscessus Complex. PLoS ONE 2015, 10, e0140166. [Google Scholar] [CrossRef] [PubMed]
- Esther, C.R.; Henry, M.M.; Molina, P.L.; Leigh, M.W. Nontuberculous Mycobacterial Infection in Young Children with Cystic Fibrosis. Pediatr. Pulmonol. 2005, 40, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Aitken, M.L.; Limaye, A.; Pottinger, P.; Whimbey, E.; Goss, C.H.; Tonelli, M.R.; Cangelosi, G.A.; Dirac, M.A.; Olivier, K.N.; Brown-Elliott, B.A. Respiratory Outbreak of Mycobacterium abscessus Subspecies massiliense in a Lung Transplant and Cystic Fibrosis Center. Am. J. Respir. Crit. Care Med. 2012, 185, 231–232. [Google Scholar] [CrossRef]
- Kim, S.Y.; Shin, S.J.; Jeong, B.H.; Koh, W.J. Successful Antibiotic Treatment of Pulmonary Disease Caused by Mycobacterium abscessus Subsp. abscessus with C-to-T Mutation at Position 19 in erm(41) Gene: Case Report. BMC Infect. Dis. 2016, 16, 207. [Google Scholar] [CrossRef]
- Howard, S.T.; Rhoades, E.; Recht, J.; Pang, X.; Alsup, A.; Kolter, R.; Lyons, C.R.; Byrd, T.F. Spontaneous Reversion of Mycobacterium abscessus from a Smooth to a Rough Morphotype Is Associated with Reduced Expression of Glycopeptidolipid and Reacquisition of an Invasive Phenotype. Microbiology 2006, 152, 1581–1590. [Google Scholar] [CrossRef]
- Jong, B.-E.; Wu, T.-S.; Chen, N.-Y.; Yang, C.-H.; Shu, C.-C.; Wang, L.-S.; Wu, T.-L.; Lu, J.-J.; Chiu, C.-H.; Lai, H.-C.; et al. Impact on Macrolide Resistance of Genetic Diversity of Mycobacterium abscessus Species. Microbiol. Spectr. 2022, 10, e02749-22. [Google Scholar] [CrossRef]
- Li, Y.M.; Tong, X.L.; Xu, H.T.; Ju, Y.; Cai, M.; Wang, C. Prevalence and Antimicrobial Susceptibility of Mycobacterium abscessus in a General Hospital, China. Biomed. Environ. Sci. 2016, 29, 85–90. [Google Scholar] [CrossRef]
- Carneiro, M.D.; Nunes, L.D.; David, S.M.; Barth, A.L. Lack of Association between rrl and erm(41) Mutations and Clarithromycin Resistance in Mycobacterium abscessus Complex. Mem. Inst. Oswaldo Cruz 2017, 112, 775–778. [Google Scholar] [CrossRef]
- Mougari, F.; Bouziane, F.; Crockett, F.; Nessar, R.; Veziris, N.; Sapriel, G.; Raskine, L.; Cambau, E. Selection of Resistance to Clarithromycin in Mycobacterium abscessus Subspecies. Antimicrob. Agents Chemother. 2017, 61, e00943-16. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, W.; Wang, Y.; Xue, Z.; Li, S.; Pang, Y. Inducible Resistance to Amikacin in Mycobacterium abscessus Isolated in Beijing, China. Infect. Drug Resist. 2022, 15, 2287–2291. [Google Scholar] [CrossRef] [PubMed]
- Ananta, P.; Kham-ngam, I.; Chetchotisakd, P.; Chaimanee, P.; Reechaipichitkul, W.; Namwat, W.; Lulitanond, V.; Faksri, K. Analysis of Drug-Susceptibility Patterns and Gene Sequences Associated with Clarithromycin and Amikacin Resistance in Serial Mycobacterium abscessus Isolates from Clinical Specimens from Northeast Thailand. PLoS ONE 2018, 13, e0208053. [Google Scholar] [CrossRef] [PubMed]
- Mougari, F.; Loiseau, J.; Veziris, N.; Bernard, C.; Bercot, B.; Sougakoff, W.; Jarlier, V.; Raskine, L.; Cambau, E.; Aubry, A.; et al. Evaluation of the New GenoType NTM-DR Kit for the Molecular Detection of Antimicrobial Resistance in Non-Tuberculous Mycobacteria. J. Antimicrob. Chemother. 2017, 72, 1669–1677. [Google Scholar] [CrossRef] [PubMed]
- Shaw, L.P.; Doyle, R.M.; Kavaliunaite, E.; Spencer, H.; Balloux, F.; Dixon, G.; Harris, K.A. Children with Cystic Fibrosis Are Infected with Multiple Subpopulations of Mycobacterium abscessus with Different Antimicrobial Resistance Profiles. Clin. Infect. Dis. 2019, 69, 1678–1686. [Google Scholar] [CrossRef]
- Chua, K.Y.L.; Bustamante, A.; Jelfs, P.; Chen, S.C.-A.; Sintchenko, V. Antibiotic Susceptibility of Diverse Mycobacterium abscessus Complex Strains in New South Wales, Australia. Pathology 2015, 47, 678–682. [Google Scholar] [CrossRef]
- Zhang, Z.; Lu, J.; Liu, M.; Wang, Y.; Zhao, Y.; Pang, Y. In Vitro Activity of Clarithromycin in Combination with Other Antimicrobial Agents against Mycobacterium abscessus and Mycobacterium massiliense. Int. J. Antimicrob. Agents 2017, 49, 383–386. [Google Scholar] [CrossRef]

{kind=link}
| Colony Morphology | Total | ||||
|---|---|---|---|---|---|
| Rough | Smooth | Rough | Mixed | ||
| M. abscessus subsp. abscessus | 9 | 1 | 9 | 4 | 17 |
| M. abscessus subsp. massiliense | 1 | 1 | 1 | 4 | 7 |
| M. abscessus subsp. bolletii | 2 | 0 | 0 | 2 | 2 |
| Isolation Chronology and Colony Morphotype | Total | ||||
|---|---|---|---|---|---|
| First Samples | Subsequent Samples | ||||
| Mixed (Rough and Smooth) | Rough | Smooth | |||
| Patient 1 | M. abscessus subsp. abscessus | 6 | 6 | 0 | 12 |
| Patient 2 | M. abscessus subsp. massiliense | 2 | 1 | 0 | 3 |
| Patient 3 | M. abscessus subsp. massiliense | 2 | 3 | 0 | 5 |
| Clarithromycin | Amikacin | |||||||
|---|---|---|---|---|---|---|---|---|
| S | I | R | S | I | R (rrs Gene Mutations) | |||
| Inducible | Acquired (rrl Gene Mutations) | |||||||
| M. abscessus subsp. abscessus (n = 18) | C28 sequevar | 1 | 1 | |||||
| T28 sequevar | 4 | 10 | 3 (WT + A2058G + A2059G, A2058C, and A2058C) | 13 | 1 | 3 (A1408G, WT + A1408G, and WT + A1408G) | ||
| M. abscessus subsp. massiliense (n = 8) | T28 sequevar | 5 | 3 (A2058C, A2058C, and A2058C) | 6 | 1 | 1 (A1408G) | ||
| M. abscessus subsp. bolletii (n = 2) | T28 sequevar | 1 | 1 | 2 | ||||
| Subsp. abscessus | Subsp. massiliense (n = 8) (%) | Subsp. bolletii (n = 2) (%) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C28 (n = 1) (%) | T28 (n = 17) (%) | ||||||||||||
| S | I | R | S | I | R | S | I | R | S | I | R | ||
| AMK | 1 | 13 (76.4) | 1 (5.8) | 3 (16.6) | 6 (75) | 1 (12.5) | 1 (12.5) | 2 (100) | |||||
| FOX | 1 | 1 (5.8) | 16 (94.1) | 2 (25) | 1 (12.5) | 5 (62.5) | 1 (50) | 1 (50) | |||||
| CIP | 1 | 1 (5.8) | 16 (94.1) | 1 (12.5) | 7 (87.5) | 2 (100) | |||||||
| CLA | IRT | 1 | 3 (16.6) | 5 (62.5) | 3 (37.5) | 2 (100) | |||||||
| LRT | 1 | 14 (82.3) | 13 (76.4) | 5 (62.5) | 3 (37.5) | 1 (50) | 1 (50) | ||||||
| DOX | 1 | 4 (23.6) | 17 (100) | 1 (12.5) | 7 (87.5) | 2 (100) | |||||||
| IMP | 1 | 17 (100) | 8 (100) | 2 (100) | |||||||||
| LNZ | 1 | 7 (41.1) | 4 (23.6) | 6 (35.2) | 3 (37.5) | 2 (25) | 3 (37.5) | 1 (50) | 1 (50) | ||||
| MIN | 1 | 17 (100) | 1 (12.5) | 7 (87.5) | 2 (100) | ||||||||
| MXF | 1 | 1 (5.8) | 1 (5.8) | 15 (88.2) | 2 (25) | 6 (75) | 2 (100) | ||||||
| SXT | 17 (100) | 2 (25) | 6 (75) | 2 (100) | |||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao Yao, J.C.; Navas Méndez, J.; Tórtola Fernández, M.T. Analysis of Phenotypic and Genotypic Susceptibility to Clarithromycin and Amikacin of Mycobacterium abscessus Complex Strains Isolated from Cystic Fibrosis Patients. Microorganisms 2023, 11, 2897. https://doi.org/10.3390/microorganisms11122897
Cao Yao JC, Navas Méndez J, Tórtola Fernández MT. Analysis of Phenotypic and Genotypic Susceptibility to Clarithromycin and Amikacin of Mycobacterium abscessus Complex Strains Isolated from Cystic Fibrosis Patients. Microorganisms. 2023; 11(12):2897. https://doi.org/10.3390/microorganisms11122897
Chicago/Turabian StyleCao Yao, Juan Carlos, Jesús Navas Méndez, and María Teresa Tórtola Fernández. 2023. "Analysis of Phenotypic and Genotypic Susceptibility to Clarithromycin and Amikacin of Mycobacterium abscessus Complex Strains Isolated from Cystic Fibrosis Patients" Microorganisms 11, no. 12: 2897. https://doi.org/10.3390/microorganisms11122897
APA StyleCao Yao, J. C., Navas Méndez, J., & Tórtola Fernández, M. T. (2023). Analysis of Phenotypic and Genotypic Susceptibility to Clarithromycin and Amikacin of Mycobacterium abscessus Complex Strains Isolated from Cystic Fibrosis Patients. Microorganisms, 11(12), 2897. https://doi.org/10.3390/microorganisms11122897