

Ameliorative Effects of Anthocyanin Metabolites on Western Diet-Induced NAFLD by Modulating Co-Occurrence Networks of Gut Microbiome

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
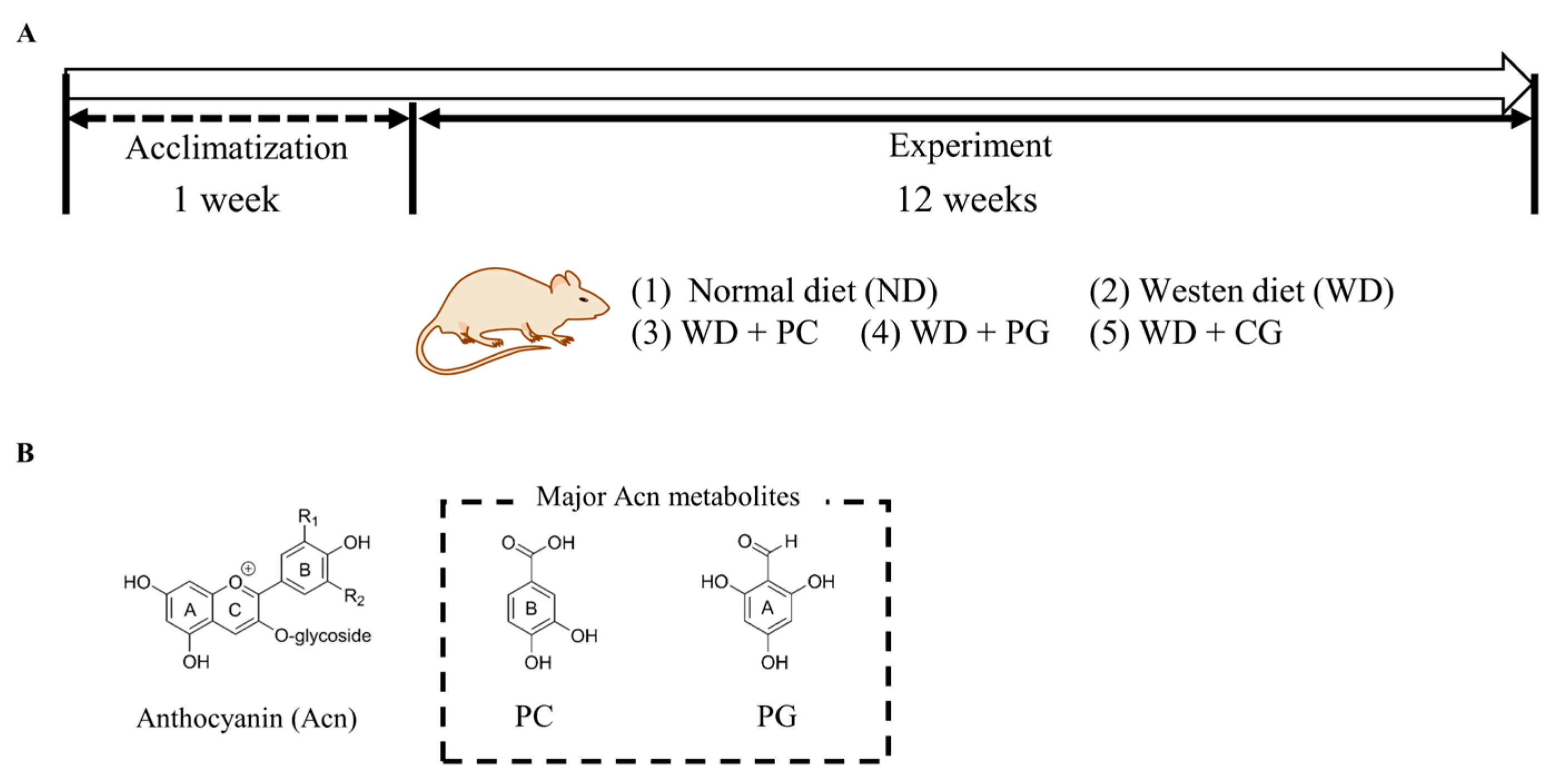
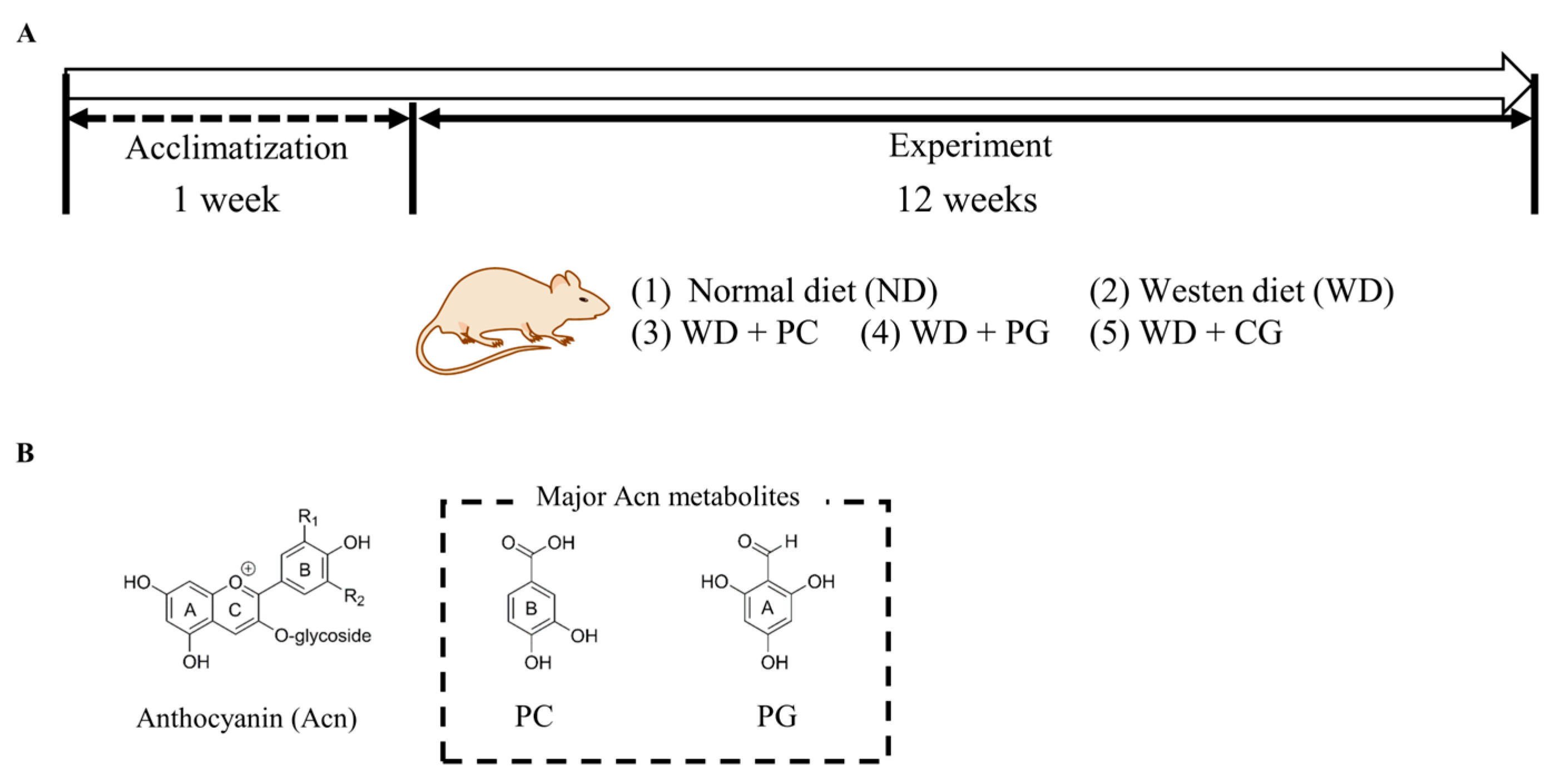
2.2. Animal Experiment Design
2.3. Measurement of Serum Biochemical Indexes
2.4. Measurement of 2-Thiobarbituric Acid Reactive Substances (TBARS)
2.5. Measurement of Hepatic Lipid
2.6. Gut Microbiome Analysis by 16S rRNA Gene Sequencing
2.7. Prediction of Functional Abundances Based on 16S rRNA Gene Sequences by PICRUSt2
2.8. Co-Occurrence Network Analysis in Gut Microbiome
2.9. Statistical Analysis
3. Results
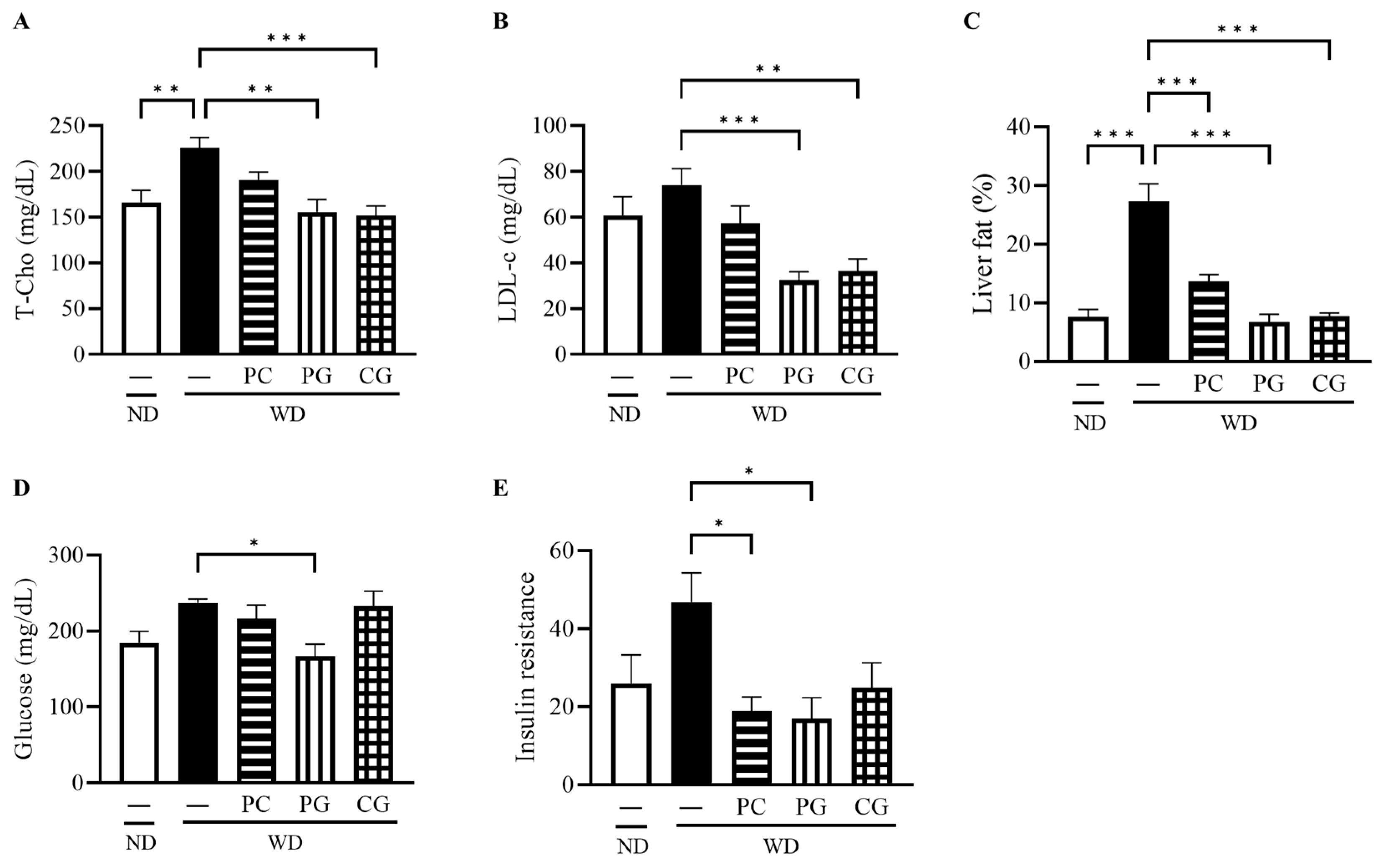
3.1. Organ Weight and Indexes of Lipid and Glucose
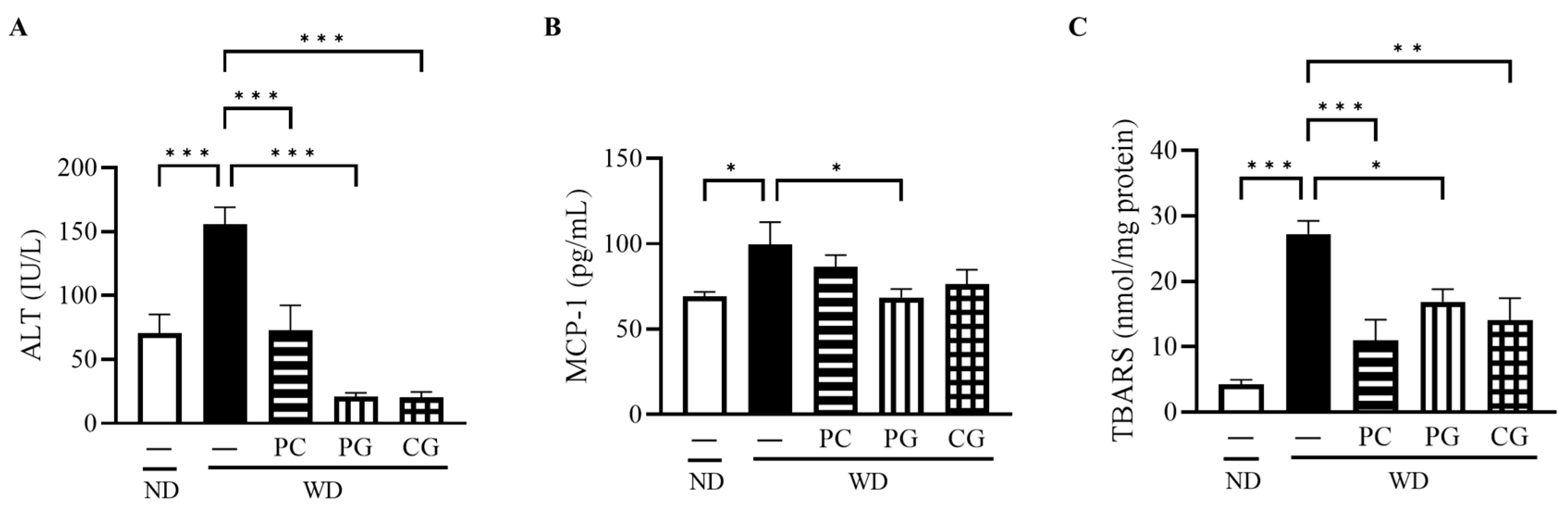
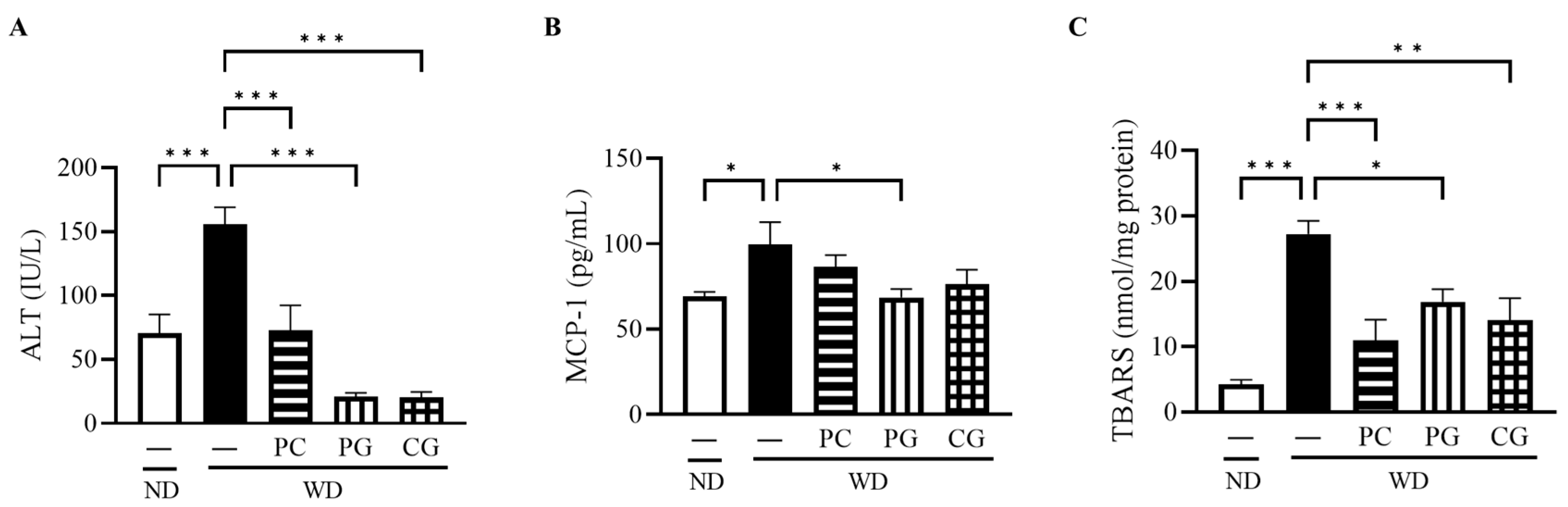
3.2. Liver Damage, Inflammation, and Oxidative Stress
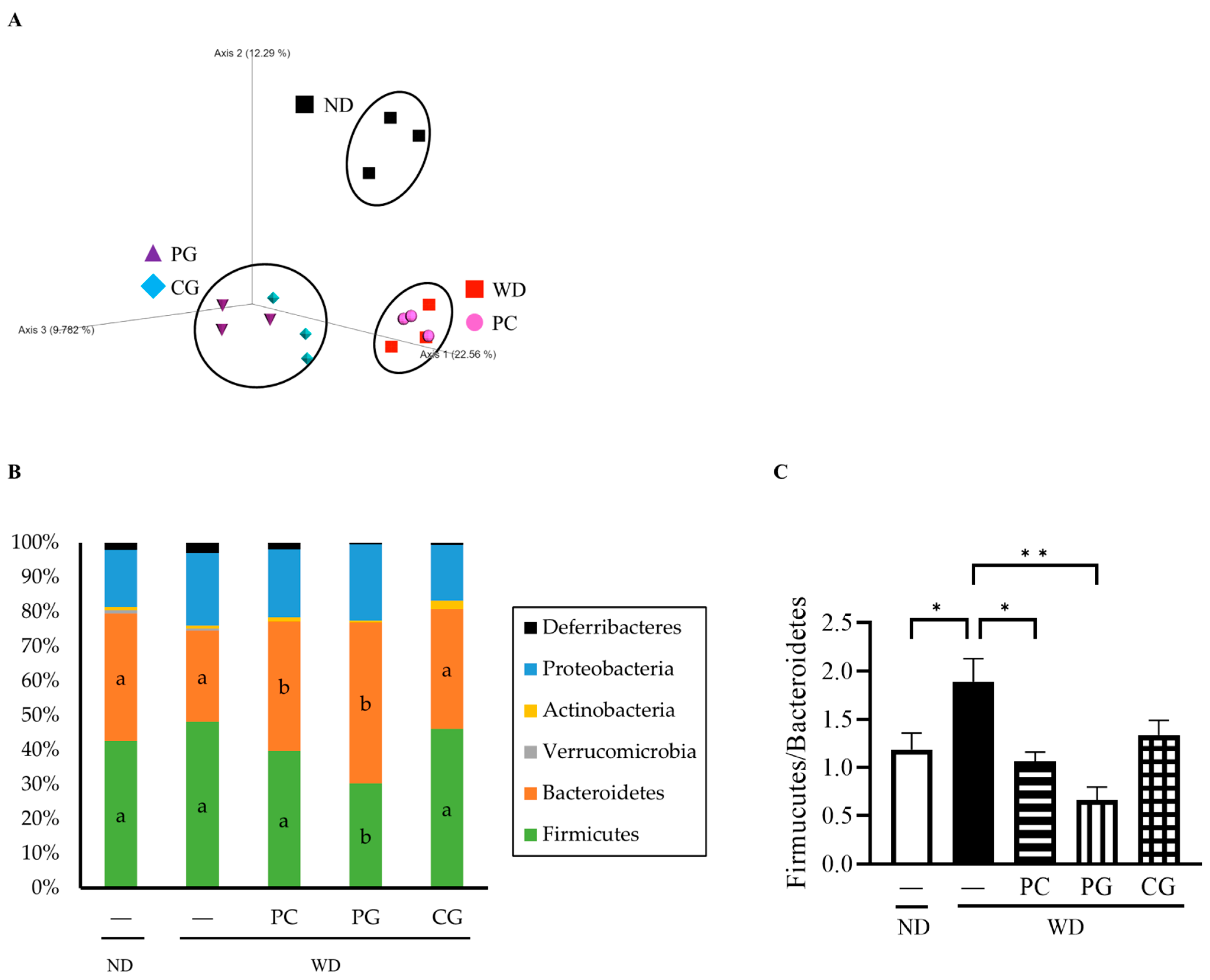
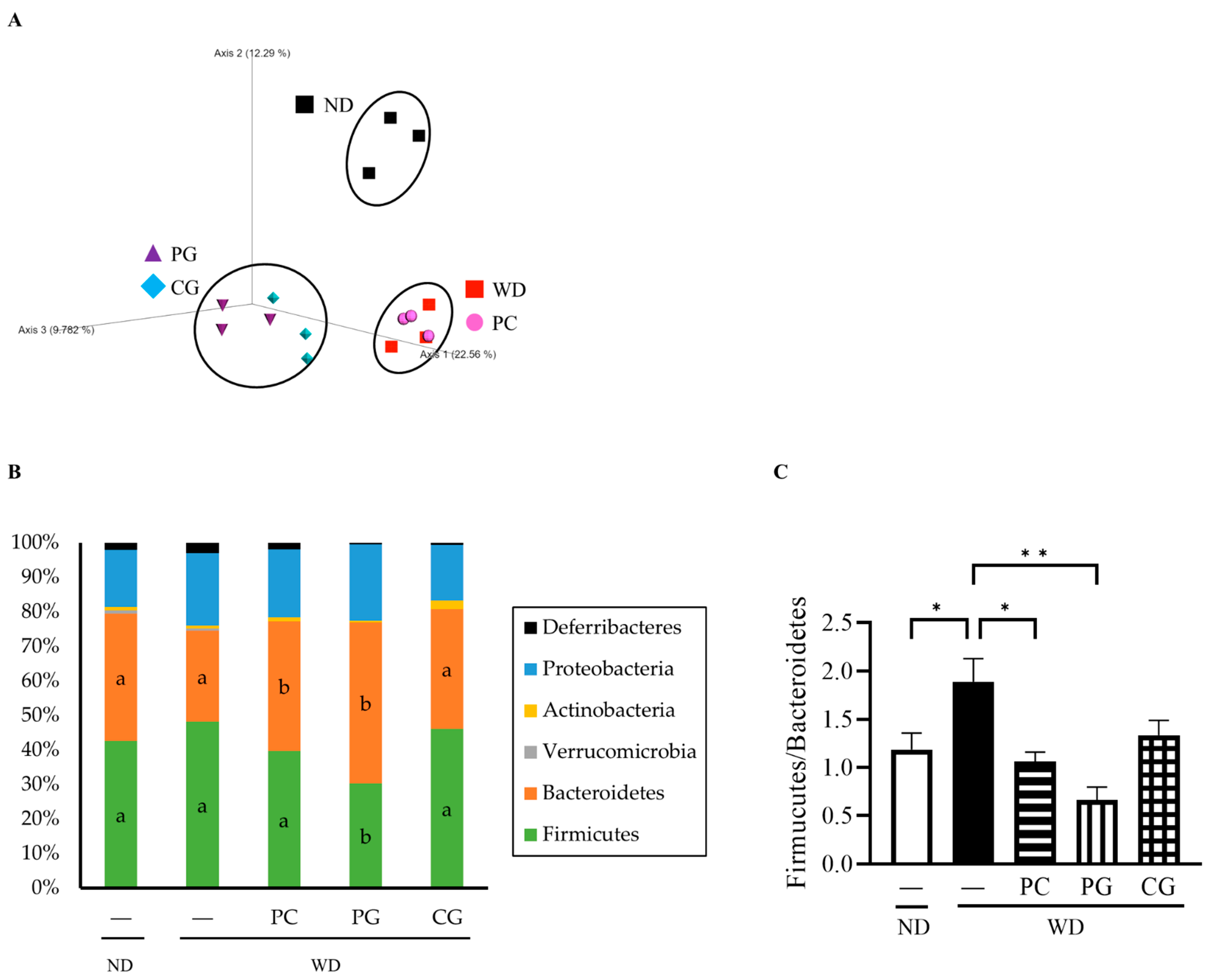
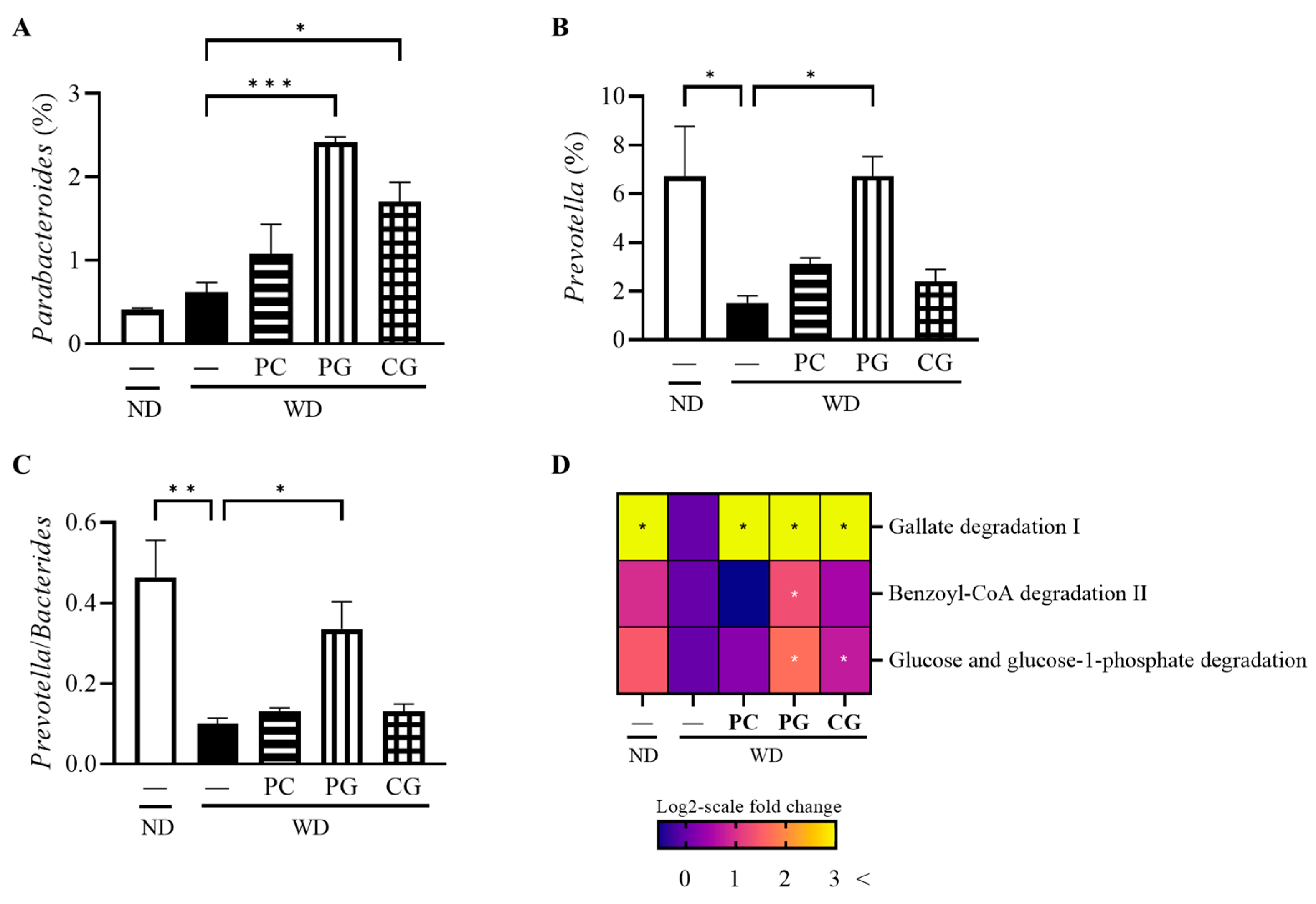
3.3. Gut Microbiome Structure Analysis by 16S rRNA Gene Sequencing and MetaCyc Pathway Analysis by Picrust2
3.4. A Co-Occurrence Network Analysis in the Gut Microbiome
4. Discussion
4.1. The Preventive Effect of Acn-M on Mice NAFLD Symptom
4.2. The Effect of Acn-M on Some Specific Gut Microbiome and Metagenomic Pathways
4.3. The Effect of Acn-M on Co-Occurrence Network of the Gut Microbiome
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Acn | anthocyanins |
| Acn-M | anthocyanins metabolites |
| ALT | alanine aminotransferase |
| BA | bilberry Acn powder |
| BW | body weight |
| CG | 0.25% PC + 0.25% PG |
| DPPH | 1,1-diphenyl-2-picrylhydrazyl |
| EFW | epididymis fat weight |
| FA | ferulic acid |
| FFA | free fatty acids |
| GA | gallic acid |
| HA | hippuric acid |
| HBA | 4-hydroxybenzoic acid |
| HFD | high fat diet |
| HMBA | 2-hydroxy-4methoxybenzoic acid |
| IR | insulin resistance |
| LDL-c | low-density lipoprotein cholesterol |
| LW | liver weight |
| MCP-1 | monocyte chemoattractant protein 1 |
| MetaCyc | metabolic pathways from all domains of life |
| NAFLD | nonalcoholic fatty liver disease |
| NASH | non-alcoholic steatohepatitis |
| ND | normal diet |
| PC | protocatechuic acid |
| PCoA | principal coordinate analysis |
| PG | phloroglucinol carboxaldehyde |
| PICRUSt2 | Phylogenetic Investigation of Communities by Reconstruction of Unobserved States |
| SA | syringic acid |
| SGLT1 | sodium-dependent glucose cotransporters |
| SparCC | Sparse Correlations for Compositional data |
| STZ | streptozotocin |
| TBARS | 2-thiobarbituric acid-reactive substances |
| T-Cho | total cholesterol |
| TG | triglycerides |
| VA | vanillic acid |
| WD | Western diet |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components (%) | ND | WD | |||
|---|---|---|---|---|---|
| − | − | PC | PG | CG | |
| Lard | 3 | 30 | 30 | 30 | 30 |
| Soybean oil | 3 | 3 | 3 | 3 | 3 |
| Corn Starch | 45 | 16.5 | 16 | 16 | 16 |
| Casein | 20 | 20 | 20 | 20 | 20 |
| Sucrose | 20 | 20 | 20 | 20 | 20 |
| Cellulose | 4 | 4 | 4 | 4 | 4 |
| Mineral Mix | 3.5 | 3.5 | 3.5 | 3.5 | 3.5 |
| Vitamin Mix | 1 | 1 | 1 | 1 | 1 |
| Cholesterol | 0 | 1.5 | 1.5 | 1.5 | 1.5 |
| Choline bitartrate | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 |
| Methionine | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 |
| PCA (PC) | 0.5 | 0.25 | |||
| PGA (PG) | 0.5 | 0.25 | |||
| Total calories (kcal/100 g) | 374 | 517 | 515 | 515 | 515 |
References
- Mehmood, A.; Zhao, L.; Wang, Y.; Pan, F.; Hao, S.; Zhang, H.; Iftikhar, A.; Usman, M. Dietary Anthocyanins as Potential Natural Modulators for the Prevention and Treatment of Non-Alcoholic Fatty Liver Disease: A Comprehensive Review. Food Res. Int. 2021, 142, 110180. [Google Scholar] [CrossRef] [PubMed]
- Rada, P.; González-Rodríguez, Á.; García-Monzón, C.; Valverde, Á.M. Understanding Lipotoxicity in NAFLD Pathogenesis: Is CD36 a Key Driver? Cell Death Dis. 2020, 11, 802. [Google Scholar] [CrossRef] [PubMed]
- Teng, M.L.; Ng, C.H.; Huang, D.Q.; Chan, K.E.; Tan, D.J.; Lim, W.H.; Yang, J.D.; Tan, E.; Muthiah, M.D. Global Incidence and Prevalence of Nonalcoholic Fatty Liver Disease. Clin. Mol. Hepatol. 2023, 29, S32–S42. [Google Scholar] [CrossRef]
- Hu, H.; Lin, A.; Kong, M.; Yao, X.; Yin, M.; Xia, H.; Ma, J.; Liu, H. Intestinal Microbiome and NAFLD: Molecular Insights and Therapeutic Perspectives. J. Gastroenterol. 2020, 55, 142–158. [Google Scholar] [CrossRef]
- Le Roy, T.; Llopis, M.; Lepage, P.; Bruneau, A.; Rabot, S.; Bevilacqua, C.; Martin, P.; Philippe, C.; Walker, F.; Bado, A.; et al. Intestinal Microbiota Determines Development of Non-Alcoholic Fatty Liver Disease in Mice. Gut 2013, 62, 1787–1794. [Google Scholar] [CrossRef]
- Csader, S.; Chen, X.; Leung, H.; Männistö, V.; Pentikäinen, H.; Tauriainen, M.-M.; Savonen, K.; El-Nezami, H.; Schwab, U.; Panagiotou, G. Gut Ecological Networks Reveal Associations between Bacteria, Exercise, and Clinical Profile in Non-Alcoholic Fatty Liver Disease Patients. mSystems 2023, e00224-23. [Google Scholar] [CrossRef] [PubMed]
- Faust, K.; Raes, J. Microbial Interactions: From Networks to Models. Nat. Rev. Microbiol. 2012, 10, 538–550. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Schlaeppi, K.; Van Der Heijden, M.G.A. Keystone Taxa as Drivers of Microbiome Structure and Functioning. Nat. Rev. Microbiol. 2018, 16, 567–576. [Google Scholar] [CrossRef]
- Wu, D.; Liu, L.; Jiao, N.; Zhang, Y.; Yang, L.; Tian, C.; Lan, P.; Zhu, L.; Loomba, R.; Zhu, R. Targeting Keystone Species Helps Restore the Dysbiosis of Butyrate-producing Bacteria in Nonalcoholic Fatty Liver Disease. iMeta 2022, 1, e61. [Google Scholar] [CrossRef]
- Nakano, H.; Wu, S.; Sakao, K.; Hara, T.; He, J.; Garcia, S.; Shetty, K.; Hou, D.-X. Bilberry Anthocyanins Ameliorate NAFLD by Improving Dyslipidemia and Gut Microbiome Dysbiosis. Nutrients 2020, 12, 3252. [Google Scholar] [CrossRef]
- Olivas-Aguirre, F.J.; Rodrigo-García, J.; Martínez-Ruiz, N.D.R.; Cárdenas-Robles, A.I.; Mendoza-Díaz, S.O.; Álvarez-Parrilla, E.; González-Aguilar, G.A.; De la Rosa, L.A.; Ramos-Jiménez, A.; Wall-Medrano, A. Cyanidin-3-O-Glucoside: Physical-Chemistry, Foodomics and Health Effects. Molecules 2016, 21, 1264. [Google Scholar] [CrossRef] [PubMed]
- Keppler, K.; Humpf, H.-U. Metabolism of Anthocyanins and Their Phenolic Degradation Products by the Intestinal Microflora. Bioorg. Med. Chem. 2005, 13, 5195–5205. [Google Scholar] [CrossRef] [PubMed]
- Kay, C.D.; Kroon, P.A.; Cassidy, A. The Bioactivity of Dietary Anthocyanins Is Likely to Be Mediated by Their Degradation Products. Mol. Nutr. Food Res. 2009, 53, 92–101. [Google Scholar] [CrossRef] [PubMed]
- De Ferrars, R.M.; Czank, C.; Zhang, Q.; Botting, N.P.; Kroon, P.; Cassidy, A.; Kay, C. The Pharmacokinetics of Anthocyanins and Their Metabolites in Humans. Br. J. Pharmacol. 2014, 171, 3268–3282. [Google Scholar] [CrossRef]
- Chen, Y.; Li, Q.; Zhao, T.; Zhang, Z.; Mao, G.; Feng, W.; Wu, X.; Yang, L. Biotransformation and Metabolism of Three Mulberry Anthocyanin Monomers by Rat Gut Microflora. Food Chem. 2017, 237, 887–894. [Google Scholar] [CrossRef]
- Sun, R.; Kang, X.; Zhao, Y.; Wang, Z.; Wang, R.; Fu, R.; Li, Y.; Hu, Y.; Wang, Z.; Shan, W.; et al. Sirtuin 3-Mediated Deacetylation of Acyl-CoA Synthetase Family Member 3 by Protocatechuic Acid Attenuates Non-Alcoholic Fatty Liver Disease. Br. J. Pharmacol. 2020, 177, 4166–4180. [Google Scholar] [CrossRef]
- Kim, K.-N.; Kang, M.-C.; Kang, N.; Kim, S.-Y.; Hyun, C.-G.; Roh, S.W.; Ko, E.-Y.; Cho, K.; Jung, W.-K.; Ahn, G.; et al. 2,4,6-Trihydroxybenzaldehyde, a Potential Anti-Obesity Treatment, Suppressed Adipocyte Differentiation in 3T3-L1 Cells and Fat Accumulation Induced by High-Fat Diet in C57BL/6 Mice. Environ. Toxicol. Pharmacol. 2015, 39, 962–968. [Google Scholar] [CrossRef]
- Chao, J.; Cheng, H.-Y.; Chang, M.-L.; Huang, S.-S.; Liao, J.-W.; Cheng, Y.-C.; Peng, W.-H.; Pao, L.-H. Gallic Acid Ameliorated Impaired Lipid Homeostasis in a Mouse Model of High-Fat Diet—And Streptozotocin-Induced NAFLD and Diabetes through Improvement of β-Oxidation and Ketogenesis. Front. Pharmacol. 2021, 11, 606759. [Google Scholar] [CrossRef]
- Ham, J.R.; Lee, H.-I.; Choi, R.-Y.; Sim, M.-O.; Seo, K.-I.; Lee, M.-K. Anti-Steatotic and Anti-Inflammatory Roles of Syringic Acid in High-Fat Diet-Induced Obese Mice. Food Funct. 2016, 7, 689–697. [Google Scholar] [CrossRef]
- Jung, Y.; Park, J.; Kang, J.; Lim, S.; Yang, W.M.; Ahn, K.S.; Kim, H.-L.; Sim, J.-E.; Youn, D.-H.; Jeong, M.-Y.; et al. Vanillic Acid Attenuates Obesity via Activation of the AMPK Pathway and Thermogenic Factors in Vivo and in Vitro. FASEB J. 2018, 32, 1388–1402. [Google Scholar] [CrossRef]
- Saravanan, N.; Nalini, N. Effect of 2-Hydroxy 4-Methoxy Benzoic Acid on an Experimental Model of Hyperlipidaemia, Induced by Chronic Ethanol Treatment. J. Pharm. Pharmacol. 2010, 59, 1537–1542. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yang, Y.; Zhang, J.; Hu, J.; Yan, X.; Zeng, S.; Huang, X.; Lin, S. Ferulic Acid Ameliorates Intrahepatic Triglyceride Accumulation in Vitro but Not in High Fat Diet-Fed C57BL/6 Mice. Food Chem. Toxicol. 2021, 149, 111978. [Google Scholar] [CrossRef] [PubMed]
- Warnick, G.R.; Knopp, R.H.; Fitzpatrick, V.; Branson, L. Estimating Low-Density Lipoprotein Cholesterol by the Friedewald Equation Is Adequate for Classifying Patients on the Basis of Nationally Recommended Cutpoints. Clin. Chem. 1990, 36, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Turner, R.C.; Holman, R.R.; Matthews, D.; Hockaday, T.D.; Peto, J. Insulin Deficiency and Insulin Resistance Interaction in Diabetes: Estimation of Their Relative Contribution by Feedback Analysis from Basal Plasma Insulin and Glucose Concentrations. Metabolism 1979, 28, 1086–1096. [Google Scholar] [CrossRef] [PubMed]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for Lipid Peroxides in Animal Tissues by Thiobarbituric Acid Reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Hu, R.; Nakano, H.; Chen, K.; Liu, M.; He, X.; Zhang, H.; He, J.; Hou, D.-X. Modulation of Gut Microbiota by Lonicera caerulea L. Berry Polyphenols in a Mouse Model of Fatty Liver Induced by High Fat Diet. Molecules 2018, 23, 3213. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for Prediction of Metagenome Functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- Friedman, J.; Alm, E.J. Inferring Correlation Networks from Genomic Survey Data. PLoS Comput. Biol. 2012, 8, e1002687. [Google Scholar] [CrossRef]
- Blondel, V.D.; Guillaume, J.-L.; Lambiotte, R.; Lefebvre, E. Fast Unfolding of Communities in Large Networks. J. Stat. Mech. Theory Exp. 2008, 2008, P10008. [Google Scholar] [CrossRef]
- Brandes, U. A Faster Algorithm for Betweenness Centrality*. J. Math. Sociol. 2001, 25, 163–177. [Google Scholar] [CrossRef]
- Shao, L.; Ling, Z.; Chen, D.; Liu, Y.; Yang, F.; Li, L. Disorganized Gut Microbiome Contributed to Liver Cirrhosis Progression: A Meta-Omics-Based Study. Front. Microbiol. 2018, 9, 3166. [Google Scholar] [CrossRef] [PubMed]
- Freeman, L.C. A Set of Measures of Centrality Based on Betweenness. Sociometry 1977, 40, 35. [Google Scholar] [CrossRef]
- Toju, H.; Yamamoto, S.; Tanabe, A.S.; Hayakawa, T.; Ishii, H.S. Network Modules and Hubs in Plant-Root Fungal Biomes. J. R. Soc. Interface 2016, 13, 20151097. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Wang, O.; Wang, Y.; Wang, K.; Ji, B.; Zhou, F. Phenolic Acids Alleviate High-Fat and High-Fructose Diet-Induced Metabolic Disorders in Rats. J. Food Biochem. 2017, 41, e12419. [Google Scholar] [CrossRef]
- Ho, G.T.T.; Kase, E.T.; Wangensteen, H.; Barsett, H. Phenolic Elderberry Extracts, Anthocyanins, Procyanidins, and Metabolites Influence Glucose and Fatty Acid Uptake in Human Skeletal Muscle Cells. J. Agric. Food Chem. 2017, 65, 2677–2685. [Google Scholar] [CrossRef]
- Savard, C.; Tartaglione, E.V.; Kuver, R.; Haigh, W.G.; Farrell, G.C.; Subramanian, S.; Chait, A.; Yeh, M.M.; Quinn, L.S.; Ioannou, G.N. Synergistic Interaction of Dietary Cholesterol and Dietary Fat in Inducing Experimental Steatohepatitis. Hepatology 2013, 57, 81–92. [Google Scholar] [CrossRef]
- Sancheti, S.; Sancheti, S.; Bafna, M.; Seo, S.-Y. 2,4,6-Trihydroxybenzaldehyde as a Potent Antidiabetic Agent Alleviates Postprandial Hyperglycemia in Normal and Diabetic Rats. Med. Chem. Res. 2011, 20, 1181–1187. [Google Scholar] [CrossRef]
- Giorgio, V.; Prono, F.; Graziano, F.; Nobili, V. Pediatric Non Alcoholic Fatty Liver Disease: Old and New Concepts on Development, Progression, Metabolic Insight and Potential Treatment Targets. BMC Pediatr. 2013, 13, 1–10. [Google Scholar] [CrossRef]
- Mathew, S.; Abraham, T.E.; Zakaria, Z.A. Reactivity of Phenolic Compounds towards Free Radicals under in Vitro Conditions. J. Food Sci. Technol. 2015, 52, 5790–5798. [Google Scholar] [CrossRef]
- Zhao, Y.; He, Z.; Hao, W.; Zhu, H.; Liu, J.; Ma, K.Y.; He, W.-S.; Chen, Z.-Y. Cholesterol-Lowering Activity of Protocatechuic Acid Is Mediated by Increasing the Excretion of Bile Acids and Modulating Gut Microbiota and Producing Short-Chain Fatty Acids. Food Funct. 2021, 12, 11557–11567. [Google Scholar] [CrossRef]
- Hu, R.; He, Z.; Liu, M.; Tan, J.; Zhang, H.; Hou, D.-X.; He, J.; Wu, S. Dietary Protocatechuic Acid Ameliorates Inflammation and Up-Regulates Intestinal Tight Junction Proteins by Modulating Gut Microbiota in LPS-Challenged Piglets. J. Anim. Sci. Biotechnol. 2020, 11, 92. [Google Scholar] [CrossRef] [PubMed]
- Armougom, F.; Henry, M.; Vialettes, B.; Raccah, D.; Raoult, D. Monitoring Bacterial Community of Human Gut Microbiota Reveals an Increase in Lactobacillus in Obese Patients and Methanogens in Anorexic Patients. PLoS ONE 2009, 4, e7125. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Liao, M.; Zhou, N.; Bao, L.; Ma, K.; Zheng, Z.; Wang, Y.; Liu, C.; Wang, W.; Wang, J.; et al. Parabacteroides Distasonis Alleviates Obesity and Metabolic Dysfunctions via Production of Succinate and Secondary Bile Acids. Cell Rep. 2019, 26, 222–235. [Google Scholar] [CrossRef] [PubMed]
- Verbrugghe, P.; Brynjólfsson, J.; Jing, X.; Björck, I.; Hållenius, F.; Nilsson, A. Evaluation of Hypoglycemic Effect, Safety and Immunomodulation of Prevotella Copri in Mice. Sci. Rep. 2021, 11, 21279. [Google Scholar] [CrossRef] [PubMed]
- Shen, F.; Zheng, R.-D.; Sun, X.-Q.; Ding, W.-J.; Wang, X.-Y.; Fan, J.-G. Gut Microbiota Dysbiosis in Patients with Non-Alcoholic Fatty Liver Disease. Hepatobiliary Pancreat. Dis. Int. 2017, 16, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Pan, P.; Lam, V.; Salzman, N.; Huang, Y.-W.; Yu, J.; Zhang, J.; Wang, L.-S. Black Raspberries and Their Anthocyanin and Fiber Fractions Alter the Composition and Diversity of Gut Microbiota in F-344 Rats. Nutr. Cancer 2017, 69, 943–951. [Google Scholar] [CrossRef]
- Plovier, H.; Everard, A.; Druart, C.; Depommier, C.; Van Hul, M.; Geurts, L.; Chilloux, J.; Ottman, N.; Duparc, T.; Lichtenstein, L.; et al. A Purified Membrane Protein from Akkermansia Muciniphila or the Pasteurized Bacterium Improves Metabolism in Obese and Diabetic Mice. Nat. Med. 2017, 23, 107–113. [Google Scholar] [CrossRef]
- Derrien, M.; Collado, M.C.; Ben-Amor, K.; Salminen, S.; de Vos, W.M. The Mucin Degrader Akkermansia Muciniphila Is an Abundant Resident of the Human Intestinal Tract. Appl. Environ. Microbiol. 2008, 74, 1646–1648. [Google Scholar] [CrossRef]
- Wright, D.P.; I Rosendale, D.; Roberton, A.M. Prevotella Enzymes Involved in Mucin Oligosaccharide Degradation and Evidence for a Small Operon of Genes Expressed during Growth on Mucin. FEMS Microbiol. Lett. 2000, 190, 73–79. [Google Scholar] [CrossRef]
- Zhao, R.; Shen, G.X. Impact of Anthocyanin Component and Metabolite of Saskatoon Berry on Gut Microbiome and Relationship with Fecal Short Chain Fatty Acids in Diet-Induced Insulin Resistant Mice. J. Nutr. Biochem. 2022, 111, 109201. [Google Scholar] [CrossRef]
- Su, H.; Xie, L.; Xu, Y.; Ke, H.; Bao, T.; Li, Y.; Chen, W. Pelargonidin-3-O-Glucoside Derived from Wild Raspberry Exerts Antihyperglycemic Effect by Inducing Autophagy and Modulating Gut Microbiota. J. Agric. Food Chem. 2020, 68, 13025–13037. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Carmody, R.N.; Kalariya, H.M.; Duran, R.M.; Moskal, K.; Poulev, A.; Kuhn, P.; Tveter, K.M.; Turnbaugh, P.J.; Raskin, I.; et al. Grape Proanthocyanidin-Induced Intestinal Bloom of Akkermansia Muciniphila Is Dependent on Its Baseline Abundance and Precedes Activation of Host Genes Related to Metabolic Health. J. Nutr. Biochem. 2018, 56, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Porter, A.W.; Young, L.Y. Benzoyl-CoA, a Universal Biomarker for Anaerobic Degradation of Aromatic Compounds. Adv. Appl. Microbiol. 2014, 88, 167–203. [Google Scholar] [CrossRef] [PubMed]
- Peekhaus, N.; Conway, T. What’s for Dinner?: Entner-Doudoroff Metabolism in Escherichia Coli. J. Bacteriol. 1998, 180, 3495–3502. [Google Scholar] [CrossRef]
- Wu, D.; Fang, S.; Yang, Z.-H.; Jiao, N.; Gao, W.; Lan, P.; Zhu, R.; Loomba, R.; Zhu, L. Mo1463 Analysis and Application of Keystone Species in Nnonalcoholic Fatty Liver Disease Based on Causal Inference and Dynamic Intervention Modeling. Gastroenterology 2020, 158, S-1412–S-1413. [Google Scholar] [CrossRef]
- Xie, G.; Wang, X.; Liu, P.; Wei, R.; Chen, W.; Rajani, C.; Hernandez, B.Y.; Alegado, R.; Dong, B.; Li, D.; et al. Distinctly Altered Gut Microbiota in the Progression of Liver Disease. Oncotarget 2016, 7, 19355–19366. [Google Scholar] [CrossRef]
- Zhang, J.; Song, L.; Wang, Y.; Liu, C.; Zhang, L.; Zhu, S.; Liu, S.; Duan, L. Beneficial Effect of Butyrate-producing Lachnospiraceae on Stress-induced Visceral Hypersensitivity in Rats. J. Gastroenterol. Hepatol. 2019, 34, 1368–1376. [Google Scholar] [CrossRef]
- Coppola, S.; Avagliano, C.; Calignano, A.; Canani, R.B. The Protective Role of Butyrate against Obesity and Obesity-Related Diseases. Molecules 2021, 26, 682. [Google Scholar] [CrossRef]
- Chen, L.; Gao, Y.; Zhao, Y.; Yang, G.; Wang, C.; Zhao, Z.; Li, S. Chondroitin Sulfate Stimulates the Secretion of H2S by Desulfovibrio to Improve Insulin Sensitivity in NAFLD Mice. Int. J. Biol. Macromol. 2022, 213, 631–638. [Google Scholar] [CrossRef]
- Boursier, J. The Severity of Nonalcoholic Fatty Liver Disease Is Associated with Gut Dysbiosis and Shift in the Metabolic Function of the Gut Microbiota. Hepatology 2016, 63, 764–775. [Google Scholar] [CrossRef]

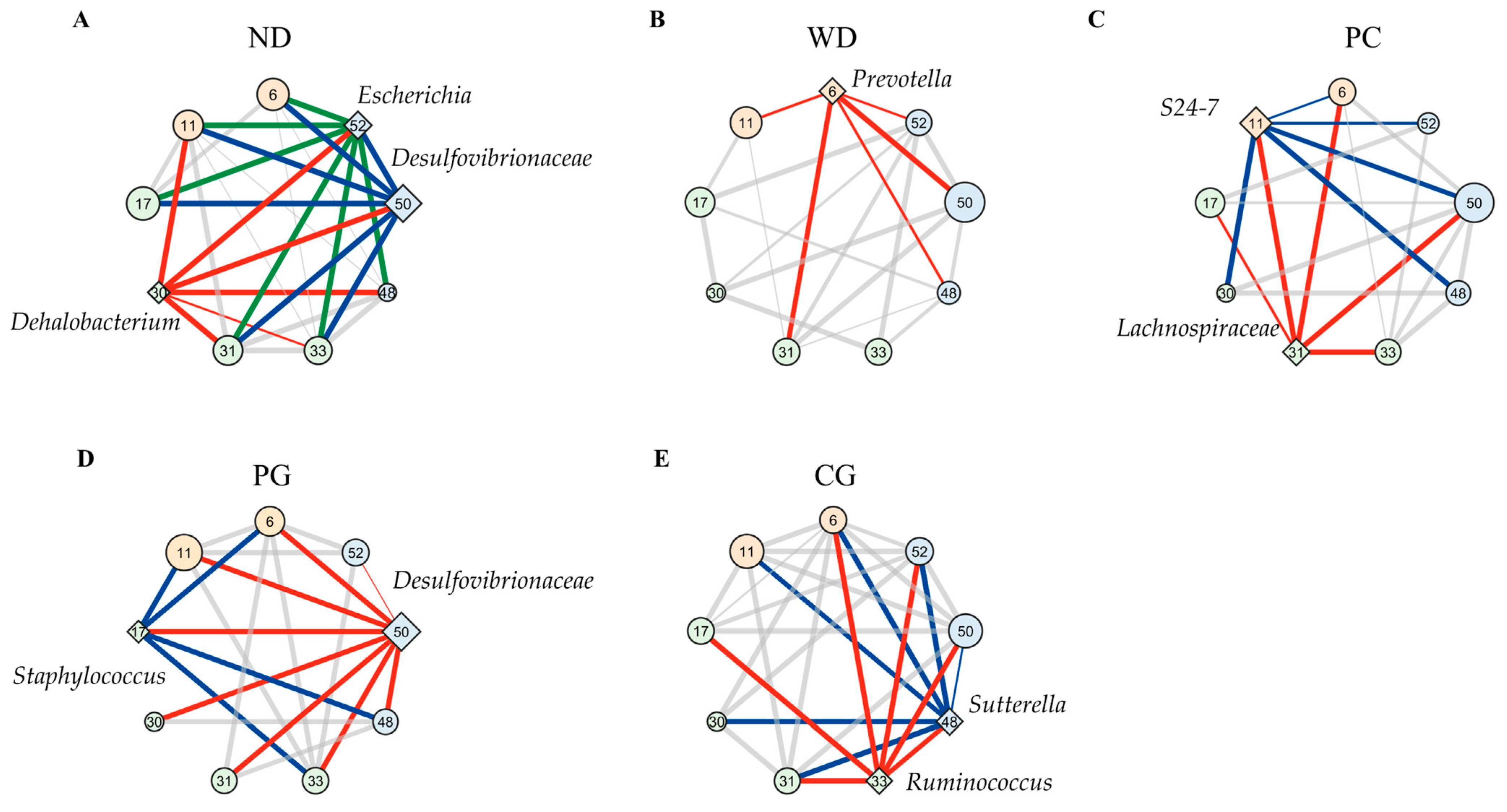
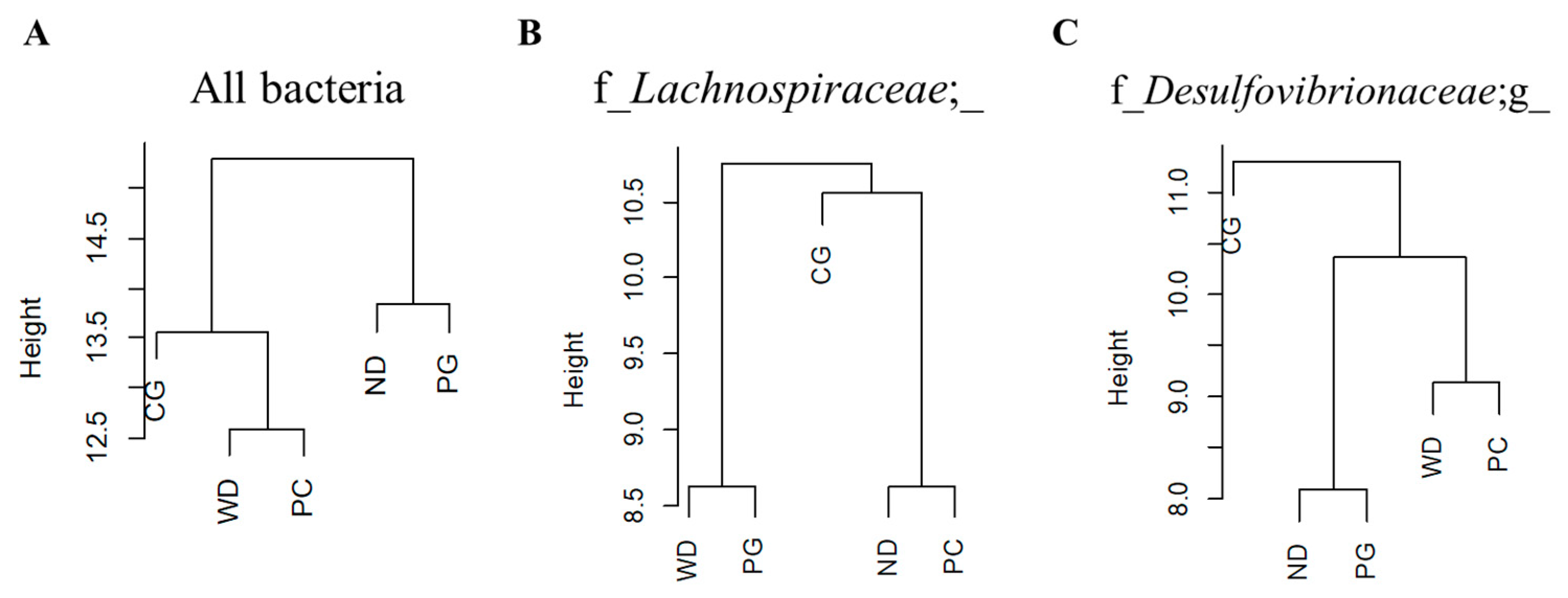
| z-Value | ND | WD | |||
|---|---|---|---|---|---|
| − | − | PC | PG | CG | |
| f_[Paraprevotellaceae]; g_[Prevotella] | 1.69 | 3.45 | −0.55 | 0.07 | 0.13 |
| f_S24-7; g_ | 0.78 | −0.07 | 2.24 | 0.14 | 1.65 |
| f_Staphylococcaceae; g_Staphylococcus | −0.78 | −0.32 | 0.12 | 2.03 | −0.35 |
| f_Dehalobacteriaceae; g_Dehalobacterium | 3.84 | −0.65 | −1.47 | −0.41 | −0.22 |
| f_Lachnospiraceae;_ | 0.24 | 0.69 | 3.05 | 0.43 | 1.12 |
| f_Lachnospiraceae; g_[Ruminococcus] | 0.41 | −0.46 | 1.22 | 0.14 | 2.97 |
| f_Alcaligenaceae; g_Sutterella | −0.75 | −0.62 | −0.89 | −0.08 | 2.70 |
| f_Desulfovibrionaceae;g_ | 2.51 | −0.26 | 0.82 | 4.40 | 0.48 |
| f_Enterobacteriaceae; g_Escherichia | 2.38 | 0.23 | 0.37 | −0.41 | 0.09 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakano, H.; Sakao, K.; Wada, K.; Hou, D.-X. Ameliorative Effects of Anthocyanin Metabolites on Western Diet-Induced NAFLD by Modulating Co-Occurrence Networks of Gut Microbiome. Microorganisms 2023, 11, 2408. https://doi.org/10.3390/microorganisms11102408
Nakano H, Sakao K, Wada K, Hou D-X. Ameliorative Effects of Anthocyanin Metabolites on Western Diet-Induced NAFLD by Modulating Co-Occurrence Networks of Gut Microbiome. Microorganisms. 2023; 11(10):2408. https://doi.org/10.3390/microorganisms11102408
Chicago/Turabian StyleNakano, Hironobu, Kozue Sakao, Koji Wada, and De-Xing Hou. 2023. "Ameliorative Effects of Anthocyanin Metabolites on Western Diet-Induced NAFLD by Modulating Co-Occurrence Networks of Gut Microbiome" Microorganisms 11, no. 10: 2408. https://doi.org/10.3390/microorganisms11102408
APA StyleNakano, H., Sakao, K., Wada, K., & Hou, D.-X. (2023). Ameliorative Effects of Anthocyanin Metabolites on Western Diet-Induced NAFLD by Modulating Co-Occurrence Networks of Gut Microbiome. Microorganisms, 11(10), 2408. https://doi.org/10.3390/microorganisms11102408