
Screening of Healthy Feral Pigeons (Columba livia domestica) in the City of Zurich Reveals Continuous Circulation of Pigeon Paramyxovirus-1 and a Serious Threat of Transmission to Domestic Poultry

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples of Feral Pigeons from the City of Zurich (Monitoring Samples)
2.2. Samples of Laying Hens from the Newcastle Disease Outbreak Due to PPMV-1, January 2022 (Outbreak Samples)
2.3. RNA Extraction from Tissue and Swab Samples
2.4. Real-Time Reverse Transcriptase PCR (rRT-PCR) for the Detection of AOAV-1
2.5. Sequencing of the F- and HN-Gene
2.6. Statistics
3. Results
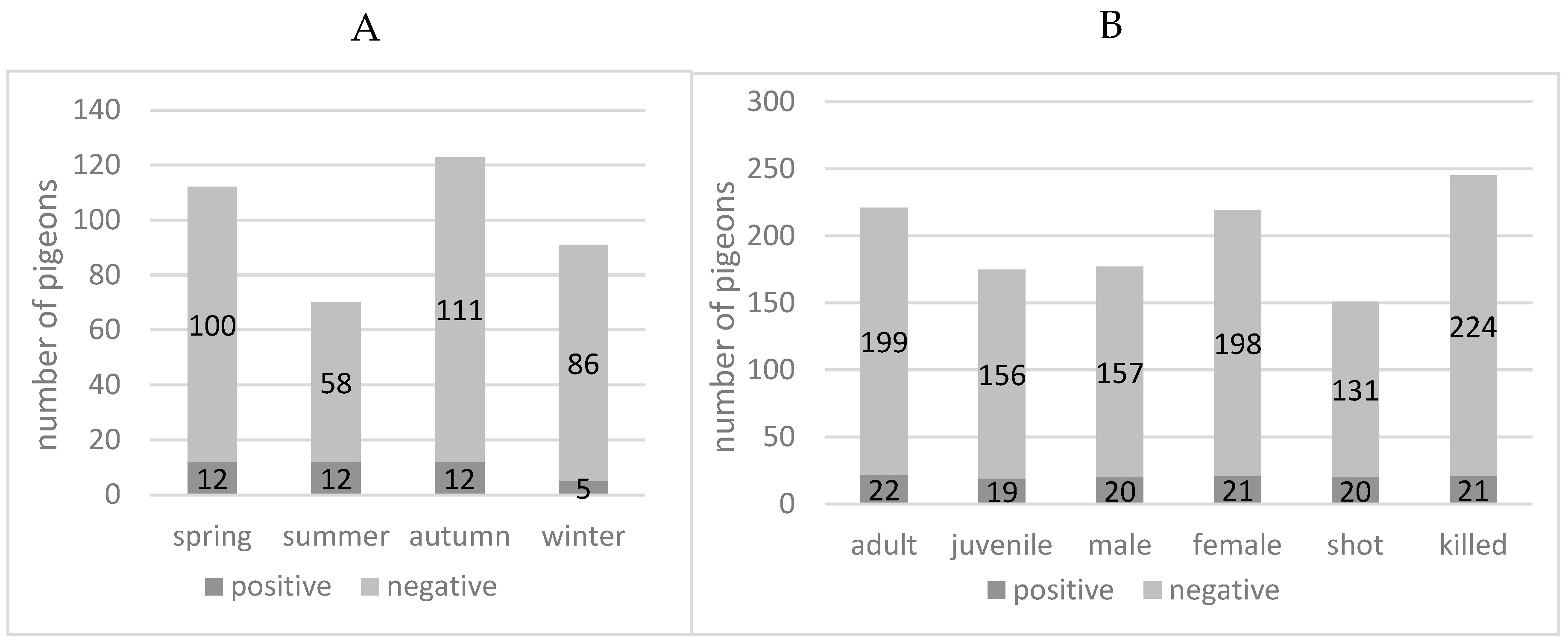
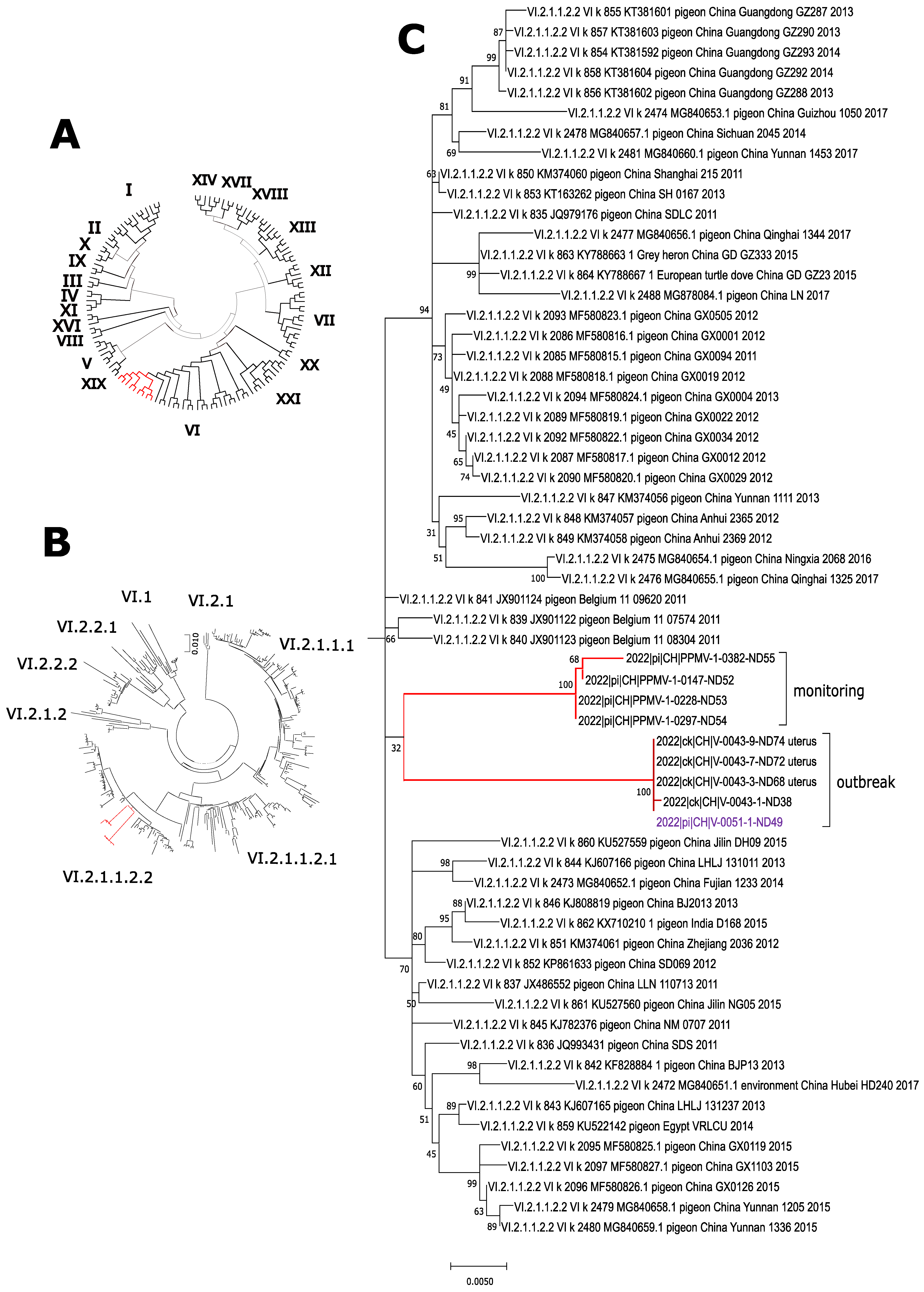
3.1. PPMV-1 in Feral Pigeons from the City of Zurich
3.2. Newcastle Disease Outbreak in Laying Hens Due to PPMV-1, January 2022
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Pigeon Carcass Data | Matrix (M)-Gen rRT-PCR | Fusion (F)-Gen r RT-PCR | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Season | Sample Nr. | Collection Data (Date/Location) | Age/ Sex | Observed Pathologies | Organs | Swabs | Organs | Swabs | |||||
| Date (dd.mm.yy) | ZIP Code | k | b | ch | cl | k | b | ch | cl | ||||
| winter | P 0115 | 22.01.21 | 8005 | a/m | none | pos | pos | neg | neg | neg | pos | neg | neg |
| P 0116 | 25.01.21 | 8051 | j/f | none | pos | pos | neg | pos | neg | pos | pos | pos | |
| P 0147 1 | 24.02.21 | 8005 | a/m | none | pos | pos | pos | pos | pos | pos | neg | pos | |
| P 0148 | 24.02.21 | 8005 | a/f | none | pos | pos | pos | pos | pos | pos | pos | pos | |
| P 0149 | 24.02.21 | 8005 | a/f | none | pos | pos | pos | pos | pos | pos | pos | neg | |
| spring | P 0172 | 30.03.21 | 8047 | j/m | none | pos | pos | pos | pos | neg | neg | pos | pos |
| P 0212 | 18.04.21 | 8005 | a/f | none | pos | neg | pos | neg | neg | neg | pos | neg | |
| P 0217 | 14.04.21 | 8050 | a/f | none | pos | pos | neg | neg | neg | pos | neg | neg | |
| P 0218 | 26.04.21 | 8047 | j/m | apathic | pos | pos | pos | n.a. | pos | pos | pos | n.a. | |
| P 0227 | 01.04.21 | 8052 | a/f | none | pos | pos | pos | pos | pos | pos | pos | pos | |
| P 0228 1 | 01.04.21 | 8052 | a/m | none | pos | pos | neg | pos | pos | pos | neg | pos | |
| P 0230 | 01.04.21 | 8052 | j/m | none | pos | pos | pos | pos | pos | neg | neg | neg | |
| P 0234 | 07.04.21 | 8051 | a/m | none | pos | pos | pos | neg | neg | neg | pos | pos | |
| P 0236 | 07.04.21 | 8051 | a/m | none | pos | neg | pos | neg | neg | neg | neg | neg | |
| P 0259 | 28.05.21 | 8047 | a/m | none | pos | neg | neg | pos | neg | neg | neg | neg | |
| P 0261 | 28.05.21 | 8047 | j/f | none | pos | pos | pos | pos | neg | neg | neg | neg | |
| P 0263 | 20.05.21 | 8004 | a/m | none | pos | pos | pos | pos | pos | neg | pos | pos | |
| summer | P 0286 | 01.06.21 | 8047 | a/f | none | pos | pos | pos | neg | neg | neg | neg | neg |
| P 0289 | 01.06.21 | 8047 | a/m | none | pos | pos | neg | pos | neg | neg | neg | neg | |
| P 0292 | 16.06.21 | 8038 | a/m | none | pos | pos | neg | pos | neg | neg | neg | neg | |
| P 0293 | 16.06.21 | 8038 | j/f | none | pos | pos | neg | pos | neg | neg | neg | neg | |
| P 0296 | 29.06.21 | 8005 | j/f | none | pos | pos | pos | neg | pos | neg | neg | neg | |
| P 0297 1 | 07.06.21 | 8004 | j/f | none | pos | pos | pos | pos | pos | neg | neg | neg | |
| P 0305 | 30.06.21 | 8038 | a/m | none | pos | pos | pos | pos | pos | neg | neg | neg | |
| P 0306 | 30.06.21 | 8038 | a/f | none | pos | neg | pos | pos | neg | neg | neg | neg | |
| P 0311 | 07.06.21 | 8044 | j/m | none | pos | pos | pos | pos | pos | neg | neg | neg | |
| P 0315 | 23.07.21 | 8001 | j/f | none | pos | pos | pos | pos | neg | neg | neg | neg | |
| P 0316 | 23.07.21 | 8001 | j/f | none | pos | neg | pos | pos | neg | neg | neg | neg | |
| P 0345 | 23.08.21 | 8001 | j/m | none | pos | pos | pos | pos | neg | neg | neg | neg | |
| autumn | P 0352 | 15.09.21 | 8001 | a/m | none | pos | neg | pos | pos | neg | neg | neg | neg |
| P 0355 | 27.09.21 | 8001 | j/f | none | pos | pos | pos | pos | neg | neg | neg | neg | |
| P 0359 | 27.09.21 | 8001 | j/m | none | pos | pos | neg | pos | neg | neg | neg | neg | |
| P 0360 | 20.09.21 | 8001 | j/f | none | pos | pos | pos | pos | neg | neg | neg | neg | |
| P 0366 | 21.09.21 | 8048 | a/? | none | pos | pos | neg | neg | neg | neg | neg | neg | |
| P 0368 | 21.09.21 | 8048 | j/f | none | pos | pos | neg | neg | neg | neg | neg | neg | |
| P 0369 | 21.09.21 | 8048 | j/f | none | pos | pos | pos | pos | neg | neg | neg | neg | |
| P 0371 | 15.09.21 | 8002 | a/m | none | pos | neg | neg | pos | neg | neg | neg | neg | |
| P 0374 | 15.09.21 | 8002 | a/f | none | pos | neg | pos | pos | neg | neg | neg | neg | |
| P 0377 | 13.09.21 | 8001 | a/f | none | pos | neg | pos | neg | neg | neg | neg | neg | |
| P 0379 | 13.09.21 | 8001 | j/m | none | pos | pos | neg | pos | neg | neg | neg | neg | |
| P 0382 1 | 02.09.21 | 8006 | j/m | apathic, moderate BC | pos | pos | pos | pos | neg | pos | neg | pos | |
References
- Dimitrov, K.M.; Abolnik, C.; Afonso, C.L.; Albina, E.; Bahl, J.; Berg, M.; Briand, F.-X.; Brown, I.H.; Choi, K.-S.; Chvala, I. Updated unified phylogenetic classification system and revised nomenclature for Newcastle disease virus. Infect. Genet. Evol. 2019, 74, 103917. [Google Scholar] [CrossRef] [PubMed]
- Rima, B.; Balkema-Buschmann, A.; Dundon, W.G.; Duprex, P.; Easton, A.; Fouchier, R.; Kurath, G.; Lamb, R.; Lee, B.; Rota, P. ICTV virus taxonomy profile: Paramyxoviridae. J. Gen. Virol. 2019, 100, 1593–1594. [Google Scholar] [CrossRef]
- WOAH (OIE). Newcastle disease (infection with newcastle disease virus). In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; World Organisation of Animal Health (Founded as Office International des Epizooties): Paris, France, 2021; Chapter 3.3.14. [Google Scholar]
- Suarez, D.L.; Miller, P.J.; Koch, G.; Mundt, E.; Rautenschlein, S. Newcastle disease, other avian paramyxoviruses, and avian metapneumovirus infections. In Diseases of Poultry, 14th ed.; Swayne, D.E., Boulianne, M., Logue, C.M., Mcdougald, L.R., Nair, V., Suares, D.L., Eds.; Wiley Blackwell: Hoboken, NJ, USA, 2020; Volume 1, pp. 109–166. ISBN 978-1-119-37116-8. [Google Scholar]
- Kaleta, E.F.; Baldauf, C. Newcastle disease in free-living and pet birds. In Newcastle Disease; Alexander, D.J., Ed.; Springer: Boston, MA, USA, 1988; pp. 197–246. ISBN 978-1-4612-8987-6. [Google Scholar]
- Aldous, E.W.; Fuller, C.M.; Mynn, J.K.; Alexander, D.J. A molecular epidemiological investigation of isolates of the variant avian paramyxovirus type 1 virus (PPMV-1) responsible for the 1978 to present panzootic in pigeons. Avian Pathol. 2004, 33, 258–269. [Google Scholar] [CrossRef] [PubMed]
- Kaleta, E.F.; Alexander, D.J.; Russell, P.H. The first isolation of the avian pmv-1 virus responsible for the current panzootic in pigeons? Avian Pathol. 1985, 14, 553–557. [Google Scholar] [CrossRef]
- Alexander, D.J.; Wilson, G.W.; Russell, P.H.; Lister, S.A.; Parsons, G. Newcastle disease outbreaks in fowl in Great Britain during 1984. Vet. Rec. 1985, 117, 429–434. [Google Scholar] [CrossRef]
- Alexander, D.J.; Russell, P.H.; Parsons, G.; Elzein, E.M.E.A.; Ballouh, A.; Cernik, K.; Engstrom, B.; Fevereiro, M.; Fleury, H.J.A.; Guittet, M.; et al. Antigenic and biological characterisation of avian paramyxovirus type I isolates from pigeons-an international collaborative study. Avian Pathol. 1985, 14, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Alexander, D.J. Newcastle disease. Br. Poult. Sci. 2001, 42, 5–22. [Google Scholar] [CrossRef]
- Alexander, D.J. Newcastle disease in the European Union 2000 to 2009. Avian Pathol. 2011, 40, 547–558. [Google Scholar] [CrossRef]
- Kaleta, E.F. Paramyxovirusinfektionen. In Krankheiten des Wirtschaftsgeflügels; Heider, G., Monreal, G., Eds.; Gustav Fischer Verlag Jena: Stuttgart, Germany, 1992; Band 1; pp. 587–661. ISBN 3-334-60348-2. [Google Scholar]
- Marlier, D.; Vindevogel, H. Viral infections in pigeons. Vet. J. 2006, 172, 40–51. [Google Scholar] [CrossRef]
- Alexander, D.J.; Wilson, G.W.; Thain, J.A.; Lister, S.A. Avian paramyxovirus type 1 infection of racing pigeons: 3 epizootiological considerations. Vet. Rec. 1984, 115, 213–216. [Google Scholar] [CrossRef]
- Vindevogel, H.; Duchatel, J.P. Panzootic Newcastle disease virus in pigeons. In Newcastle Disease; Alexander, D.J., Ed.; Springer: Boston, MA, USA, 1988; pp. 184–196. ISBN 978-1-4612-8987-6. [Google Scholar]
- O’Reilly, P.J.; McCullough, S.; Alexander, D.J.; De Burca, M. Recent avian paramyxovirus infections on the island of Ireland. In Proceedings of the Commission of the European Communities Meeting on Virus Diseases of Poultry-New and Evolving Pathogens, Brussels, Belgium, 18–19 October 1994; pp. 47–63. [Google Scholar]
- Werner, O.; Römer-Oberdörfer, A.; Köllner, B.; Manvell, R.J.; Alexander, D.J. Characterization of avian paramyxovirus type 1 strains isolated in Germany during 1992 to 1996. Avian Pathol. 1999, 28, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Dodovski, A.; Krstevski, K.; Ivancho, N. Classical and molecular characterization of pigeon paramyxovirus type 1 (PPMV-1) isolated from backyard poultry–first report in Macedonia. Maced. Vet. Rev. 2013, 36, 33–39. [Google Scholar]
- Abolnik, C.; Gerdes, G.H.; Kitching, J.; Swanepoel, S.; Romito, M.; Bisschop, S.P.R. Characterization of pigeon paramyxoviruses (Newcastle disease virus) isolated in South Africa from 2001 to 2006. Onderstepoort J. Vet. Res. 2008, 75, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.S.; Strong, I.; Alexander, D.J. Evaluation of the molecular basis of pathogenicity of the variant Newcastle disease viruses termed “pigeon PMV-1 viruses”. Arch. Virol. 1994, 134, 403–411. [Google Scholar] [CrossRef]
- Hüppi, L.; Ruggli, N.; Python, S.; Hoop, R.; Albini, S.; Grund, C.; Vögtlin, A. Experimental pigeon paramyxovirus-1 infection in chicken: Evaluation of infectivity, clinical and pathological manifestations and diagnostic methods. J. Gen. Virol. 2020, 101, 156–167. [Google Scholar] [CrossRef]
- Guo, H.; Liu, X.; Xu, Y.; Han, Z.; Shao, Y.; Kong, X.; Liu, S. A comparative study of pigeons and chickens experimentally infected with PPMV-1 to determine antigenic relationships between PPMV-1 and NDV strains. Vet. Microbiol. 2014, 168, 88–97. [Google Scholar] [CrossRef]
- Dortmans, J.C.; Fuller, C.M.; Aldous, E.W.; Rottier, P.J.; Peeters, B.P. Two genetically closely related pigeon paramyxovirus type 1 (PPMV-1) variants with identical velogenic fusion protein cleavage sites but with strongly contrasting virulence. Vet. Microbiol. 2010, 143, 139–144. [Google Scholar] [CrossRef]
- Kommers, G.D.; King, D.J.; Seal, B.S.; Brown, C.C. Virulence of six heterogeneous-origin Newcastle disease virus isolates before and after sequential passages in domestic chickens. Avian Pathol. 2003, 32, 81–93. [Google Scholar] [CrossRef]
- Dortmans, J.C.; Rottier, P.J.; Koch, G.; Peeters, B.P. Passaging of a Newcastle disease virus pigeon variant in chickens results in selection of viruses with mutations in the polymerase complex enhancing virus replication and virulence. J. Gen. Virol. 2011, 92, 336–345. [Google Scholar] [CrossRef]
- Anonymous. Swiss Animal Disease Ordinance (Tierseuchenverordnung TSV). Available online: https://www.fedlex.admin.ch/eli/cc/1995/3716_3716_3716/de (accessed on 11 August 2022).
- Meier, S.; Kreyenbühl, K.; Hüssy, D.; Grund, C.; Albini, S. Poultry medicine: What’s your diagnosis? Schweiz. Arch. Tierheilkd. 2021, 163, 153–156. [Google Scholar] [CrossRef]
- BLV. Bundesamt für Lebensmittelsicherheit und Veterinärwesen, Federal Food Safety and Veterinary Office. Available online: https://www.infosm.blv.admin.ch (accessed on 11 August 2022).
- Schemera, B.; Toro, H.; Herbst, W.; Kaleta, E.F. Conjunctivitis and general symptoms in man induced by the Newcastle Disease Virus. Dtsch. Tieraerztl. Wochenschr. 1987, 94, 383–384. [Google Scholar]
- Keeney, A.H.; Hunter, M.C. Human infection with the Newcastle virus of fowls. AMA Arch. Ophthalmol. 1950, 44, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Zehetbauer, G.; Kunz, C.; Thaler, A. Cases of pseudo fowl plaque (Newcastle disease) in man in Lower Austria. Wien. Klin. Wochenschr. 1971, 83, 878–880. [Google Scholar] [PubMed]
- Kim, L.M.; King, D.J.; Guzman, H.; Tesh, R.B.; Travassos Da Rosa, A.P.A.; Bueno, R.; Dennett, J.A.; Afonso, C.L. Biological and phylogenetic characterization of pigeon paramyxovirus serotype 1 circulating in wild North American pigeons and doves. J. Clin. Microbiol. 2008, 46, 3303–3310. [Google Scholar] [CrossRef] [PubMed]
- Mase, M.; Kanehira, K. Phylogenetic analysis of avian paramyxovirus serotype-1 in pigeons in Japan. J. Vet. Med. Sci. 2015, 77, 919–923. [Google Scholar] [CrossRef]
- Lillehaug, A.; Jonassen, C.M.; Bergsjø, B.; Hofshagen, M.; Tharaldsen, J.; Nesse, L.L.; Handeland, K. Screening of Feral Pigeon (Colomba livia), Mallard (Anas platyrhynchos) and Graylag Goose (Anser anser) Populations for Campylobacter spp., Salmonella spp., Avian Influenza Virus and Avian Paramyxovirus. Acta Vet. Scand. 2005, 46, 193–202. [Google Scholar] [CrossRef]
- Huthmann, E. Untersuchungen an Ringeltauben (Columba Columbus) zum Nachweis von Infektionen mit Dem Aviären Paramyxovirus 1 (APMV-1); Ludwig-Maximilians-Universität München: München, Germany, 2009. [Google Scholar]
- Bundesamt für Landwirtschaft (BLW). Agrarbericht 2021 (Produktionssystembeiträge RAUS-Programm). Available online: https://www.agrarbericht.ch/de/politik/direktzahlungen/produktionssystembeitraege (accessed on 11 August 2022).
- Baumer, A.; Feldmann, J.; Renzullo, S.; Müller, M.; Thür, B.; Hofmann, M.A. Epidemiology of avian influenza virus in wild birds in Switzerland between 2006 and 2009. Avian Dis. 2010, 54, 875–884. [Google Scholar] [CrossRef]
- Hoffmann, B.; Depner, K.; Schirrmeier, H.; Beer, M. A universal heterologous internal control system for duplex real-time RT-PCR assays used in a detection system for pestiviruses. J. Virol. Methods 2006, 136, 200–209. [Google Scholar] [CrossRef]
- Wise, M.G.; Suarez, D.L.; Seal, B.S.; Pedersen, J.C.; Senne, D.A.; King, D.J.; Kapczynski, D.R.; Spackman, E. Development of a real-time reverse-transcription PCR for detection of Newcastle disease virus RNA in clinical samples. J. Clin. Microbiol. 2004, 42, 329–338. [Google Scholar] [CrossRef]
- Hoffmann, B.; Beer, M.; Schelp, C.; Schirrmeier, H.; Depner, K. Validation of a real-time RT-PCR assay for sensitive and specific detection of classical swine fever. J. Virol. Methods 2005, 130, 36–44. [Google Scholar] [CrossRef]
- Lomniczi, B.; Wehmann, E.; Herczeg, J.; Ballagi-Pordany, A.; Kaleta, E.F.; Werner, O.; Meulemans, G.; Jorgensen, P.H.; Mante, A.P.; Gielkens, A.L.J. Newcastle disease outbreaks in recent years in western Europe were caused by an old (VI) and a novel genotype (VII). Arch. Virol. 1998, 143, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Minimap2: Pairwise alignment for nucleotide sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Signorell, A.; Aho, K.; Alfons, A.; Anderegg, N.; Aragon, T.; Arachchige, C.; Arppe, A.; Baddeley, A.; Barton, K.; Bolker, B.; et al. DescTools: Tools for Descriptive Statistics. R Package Version 0.99.44; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Ime4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Barbezange, C.; Jestin, V. Monitoring of pigeon paramyxovirus type-1 in organs of pigeons naturally infected with Salmonella Typhimurium. Avian Pathol. 2003, 32, 277–283. [Google Scholar] [CrossRef]
- Diel, D.G.; da Silva, L.H.A.; Liu, H.; Wang, Z.; Miller, P.J.; Afonso, C.L. Genetic diversity of avian paramyxovirus type 1: Proposal for a unified nomenclature and classification system of Newcastle disease virus genotypes. Infect. Genet. Evol. 2012, 12, 1770–1779. [Google Scholar] [CrossRef]
- Sabra, M.; Dimitrov, K.M.; Goraichuk, I.V.; Wajid, A.; Sharma, P.; Williams-Coplin, D.; Basharat, A.; Rehmani, S.F.; Muzyka, D.V.; Miller, P.J.; et al. Phylogenetic assessment reveals continuous evolution and circulation of pigeon-derived virulent avian avulaviruses 1 in Eastern Europe, Asia, and Africa. BMC Vet. Res. 2017, 13, 291. [Google Scholar] [CrossRef]
- Rose, E.; Nagel, P.; Haag-Wackernagel, D. Spatio-temporal use of the urban habitat by feral pigeons (Columba livia). Behav. Ecol. Sociobiol. 2006, 60, 242–254. [Google Scholar] [CrossRef]
- Janiga, M. Einige prinzipielle Erkenntnisse über die Wanderungen nach der Nahrung von Stadtpopulationen der Tauben (Columba livia f. domestica) von Bratislava hinaus in die Umgebung. Sylvia 1983, 22, 3–18. [Google Scholar]
- Sol, D.; Senar, J.C. Urban pigeon populations: Stability, home range, and the effect of removing individuals. Can. J. Zool. 1995, 73, 1154–1160. [Google Scholar] [CrossRef]
- Haag-Wackernagel, D. Regulation of the street pigeon in Basel. Wildl. Soc. Bull. 1995, 23, 256–260. [Google Scholar]
- Mattmann, P.; Marti, H.; Borel, N.; Jelocnik, M.; Albini, S.; Vogler, B.R. Chlamydiaceae in wild, feral and domestic pigeons in Switzerland and insight into population dynamics by Chlamydia psittaci multilocus sequence typing. PLoS ONE 2019, 14, e0226088. [Google Scholar] [CrossRef] [PubMed]
| Method | Target | Primers and Probes | Sequence (5′-3′) | Amplicon Size (bp) | Ref. |
|---|---|---|---|---|---|
| Avian Orthoavulavirus-1 (AOAV-1) real-time RT-PCR | M-protein gene | M+4100 | AGT GAT GTG CTC GGA CCT TC | 121 | [39] |
| M-4220 | CCT GAG GAG AGG CAT TTG CTA | ||||
| M+4169 | FAM-TTC TCT AGC AGT GGG ACA GCC TGC-TAMRA | ||||
| F-protein gene | F+4839 | TCC GGA GGA TAC AAG GGT CT | 101 | ||
| F-4939 | AGC TGT TGC AAC CCC AAG | ||||
| F+4894 | FAM-AAG CGT TTC CTC CTT CCT CCA-TAMRA | ||||
| eGFP | eGFP1-F | GAC CAC TAC CAG CAG AAC AC | 132 | [40] | |
| eGFP2-R | GAA CTC CAG CAG GAC CAT G | ||||
| eGFP1-Hex | HEX-AGC ACC CAG TCC GCC CTG AGC A-BHQ1 | ||||
| Amplification of fusion (F)- and hemagglutinin-neuraminidase (HN)-gene fragments for sequencing | F-protein gene | PPMV-1_fw4283 | CAA GCT GGC ACC CAA CGT | 1644 | |
| PPMV-1_rev5927 | TGC CTG TCA CGA TGA CTT GAG ART C | ||||
| F-protein gene | PPMV-1_fw5369 | TAT AYT GTA TGA CTC ACA GAC TC | 944 | ||
| PPMV-1_rev6340 | CCG TTC TAC CCG TGB ATT GC | ||||
| F-protein gene | PPMV-1_fw5841 | GGA UAA CYC URA GGC UCA G | 1797 | ||
| PPMV1_rev7638 | GGTACAAGAARTGAGAYGTCCCTA | ||||
| HN gene | PPMV1_fw7219 | CACATYAATGGTGCACGGAAG | 1306 | ||
| PPMV-1_rev8524 | CTG CTG ATA AYG AGA TGR TCR |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | JX901124|pi|BE|2011|09620 | ||||||||||||
| 2 | JX901122|pi|BE|2011|07574 | 0.0054 | |||||||||||
| 3 | JX901123|pi|BE|2011|08304 | 0.0054 | 0.0061 | ||||||||||
| monitoring | 4 | 2022|pi|CH|PPMV-1-0228-ND53 | 0.0184 | 0.0190 | 0.0202 | ||||||||
| 5 | 2022|pi|CH|PPMV-1-0147-ND52 | 0.0190 | 0.0196 | 0.0209 | 0.0006 | ||||||||
| 6 | 2022|pi|CH|PPMV-1-0297-ND54 | 0.0184 | 0.0190 | 0.0202 | 0.0000 | 0.0006 | |||||||
| 7 | 2022|pi|CH|PPMV-1-0382-ND55 | 0.0227 | 0.0234 | 0.0246 | 0.0042 | 0.0036 | 0.0042 | ||||||
| outbreak | 8 | 2022|ck|CH|V-0043-3-ND68_uterus | 0.0252 | 0.0271 | 0.0284 | 0.0366 | 0.0373 | 0.0366 | 0.0412 | ||||
| 9 | 2022|ck|CH|V-0043-7-ND72_uterus | 0.0252 | 0.0271 | 0.0284 | 0.0366 | 0.0373 | 0.0366 | 0.0412 | 0.0000 | ||||
| 10 | 2022|ck|CH|V-0043-9-ND74 uterus | 0.0252 | 0.0271 | 0.0284 | 0.0366 | 0.0373 | 0.0366 | 0.0412 | 0.0000 | 0.0000 | |||
| 11 | 2022|ck|CH|V-0043-1-ND38 | 0.0258 | 0.0277 | 0.0290 | 0.0373 | 0.0379 | 0.0373 | 0.0418 | 0.0006 | 0.0006 | 0.0006 | ||
| 12 | 2022|pi|CH|V-0051-1-ND49 | 0.0252 | 0.0271 | 0.0284 | 0.0366 | 0.0373 | 0.0366 | 0.0412 | 0.0000 | 0.0000 | 0.0000 | 0.0006 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Annaheim, D.; Vogler, B.R.; Sigrist, B.; Vögtlin, A.; Hüssy, D.; Breitler, C.; Hartnack, S.; Grund, C.; King, J.; Wolfrum, N.; et al. Screening of Healthy Feral Pigeons (Columba livia domestica) in the City of Zurich Reveals Continuous Circulation of Pigeon Paramyxovirus-1 and a Serious Threat of Transmission to Domestic Poultry. Microorganisms 2022, 10, 1656. https://doi.org/10.3390/microorganisms10081656
Annaheim D, Vogler BR, Sigrist B, Vögtlin A, Hüssy D, Breitler C, Hartnack S, Grund C, King J, Wolfrum N, et al. Screening of Healthy Feral Pigeons (Columba livia domestica) in the City of Zurich Reveals Continuous Circulation of Pigeon Paramyxovirus-1 and a Serious Threat of Transmission to Domestic Poultry. Microorganisms. 2022; 10(8):1656. https://doi.org/10.3390/microorganisms10081656
Chicago/Turabian StyleAnnaheim, Désirée, Barbara Renate Vogler, Brigitte Sigrist, Andrea Vögtlin, Daniela Hüssy, Christian Breitler, Sonja Hartnack, Christian Grund, Jacqueline King, Nina Wolfrum, and et al. 2022. "Screening of Healthy Feral Pigeons (Columba livia domestica) in the City of Zurich Reveals Continuous Circulation of Pigeon Paramyxovirus-1 and a Serious Threat of Transmission to Domestic Poultry" Microorganisms 10, no. 8: 1656. https://doi.org/10.3390/microorganisms10081656
APA StyleAnnaheim, D., Vogler, B. R., Sigrist, B., Vögtlin, A., Hüssy, D., Breitler, C., Hartnack, S., Grund, C., King, J., Wolfrum, N., & Albini, S. (2022). Screening of Healthy Feral Pigeons (Columba livia domestica) in the City of Zurich Reveals Continuous Circulation of Pigeon Paramyxovirus-1 and a Serious Threat of Transmission to Domestic Poultry. Microorganisms, 10(8), 1656. https://doi.org/10.3390/microorganisms10081656