
Activity of Drug Combinations against Mycobacterium abscessus Grown in Aerobic and Hypoxic Conditions

 , , , , and
, , , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Growth of Mab-10 under Aerobic (A) and Hypoxic (H) Conditions
2.3. Measurement of Drug Activity against A and H Cells
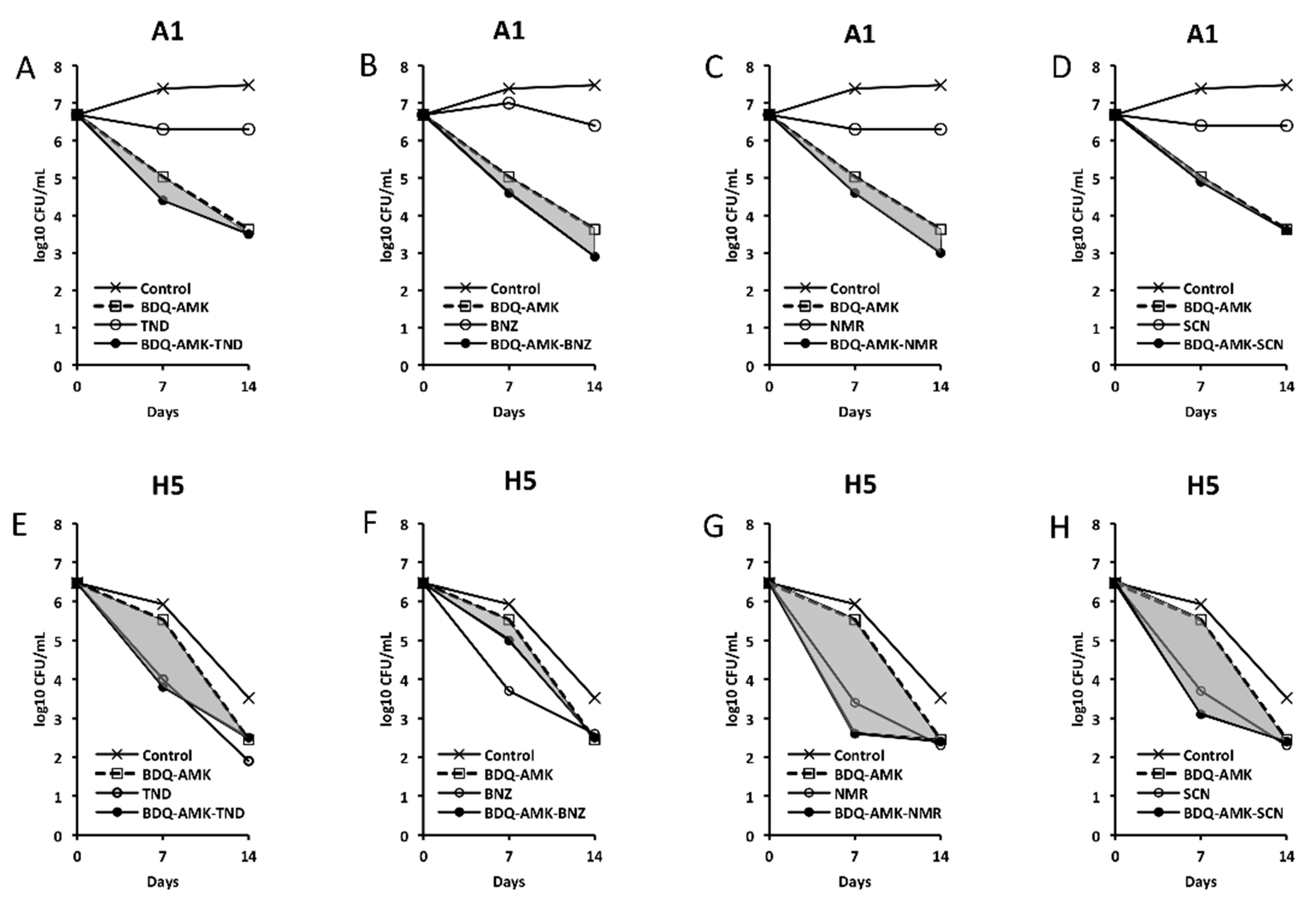
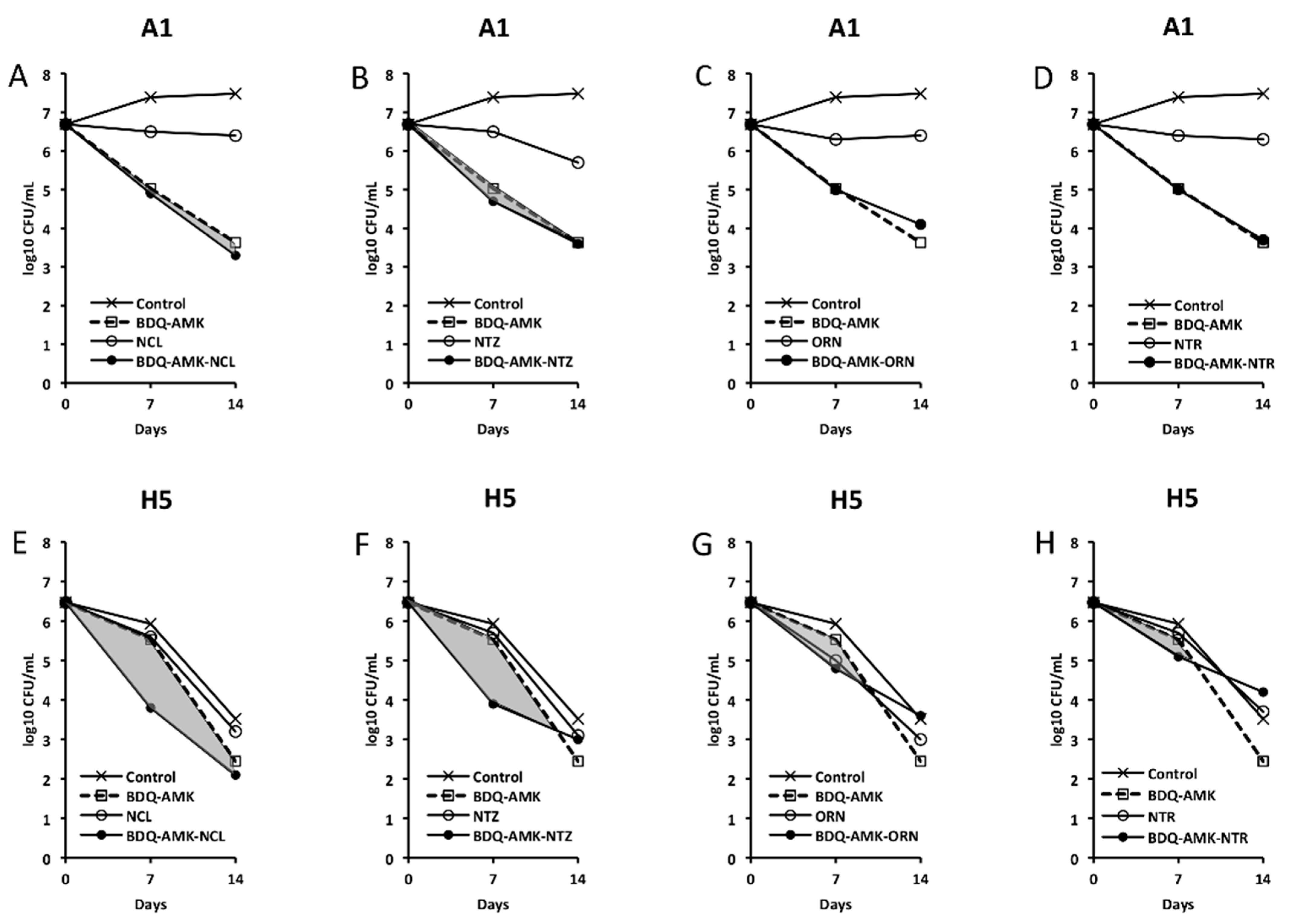
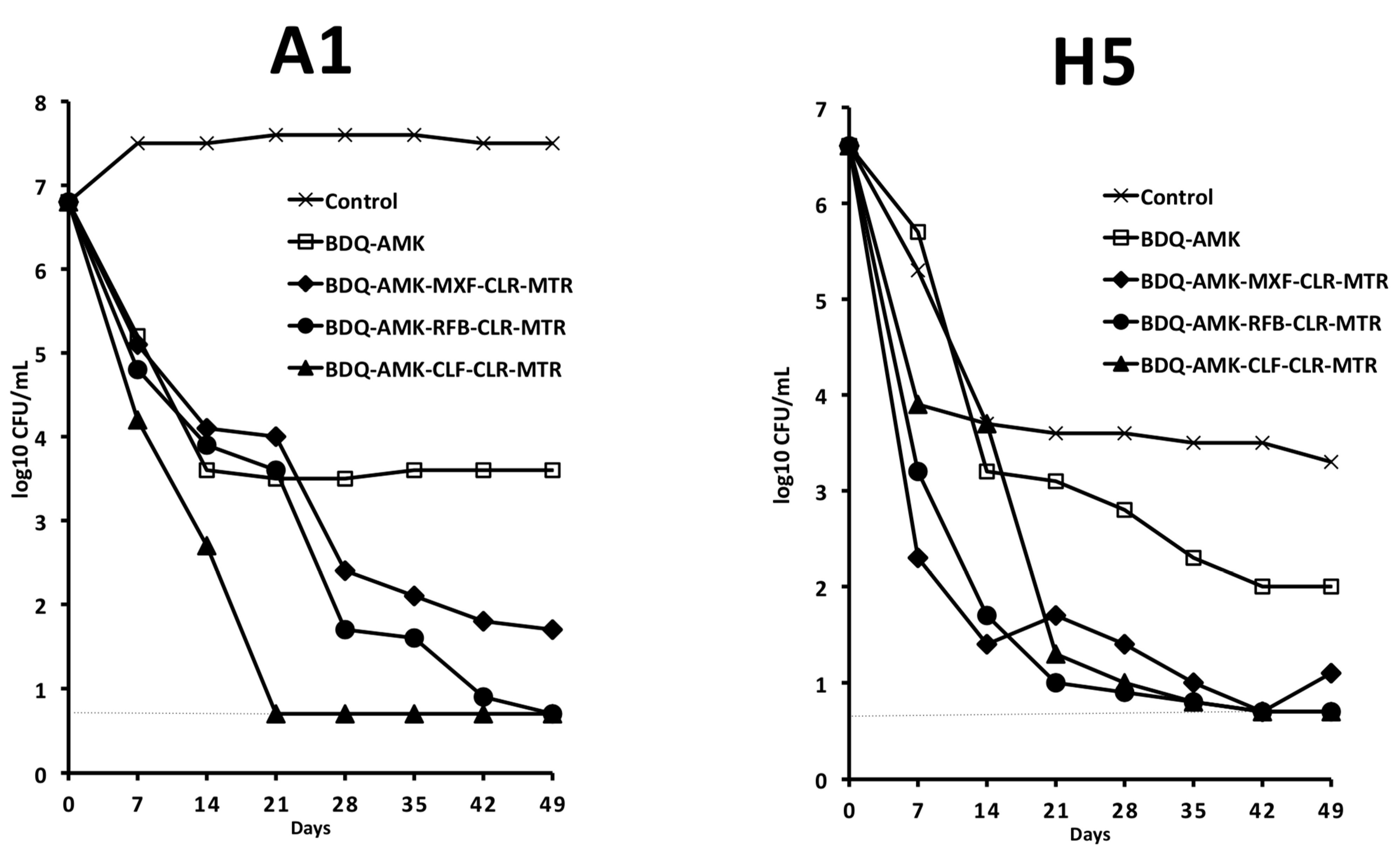
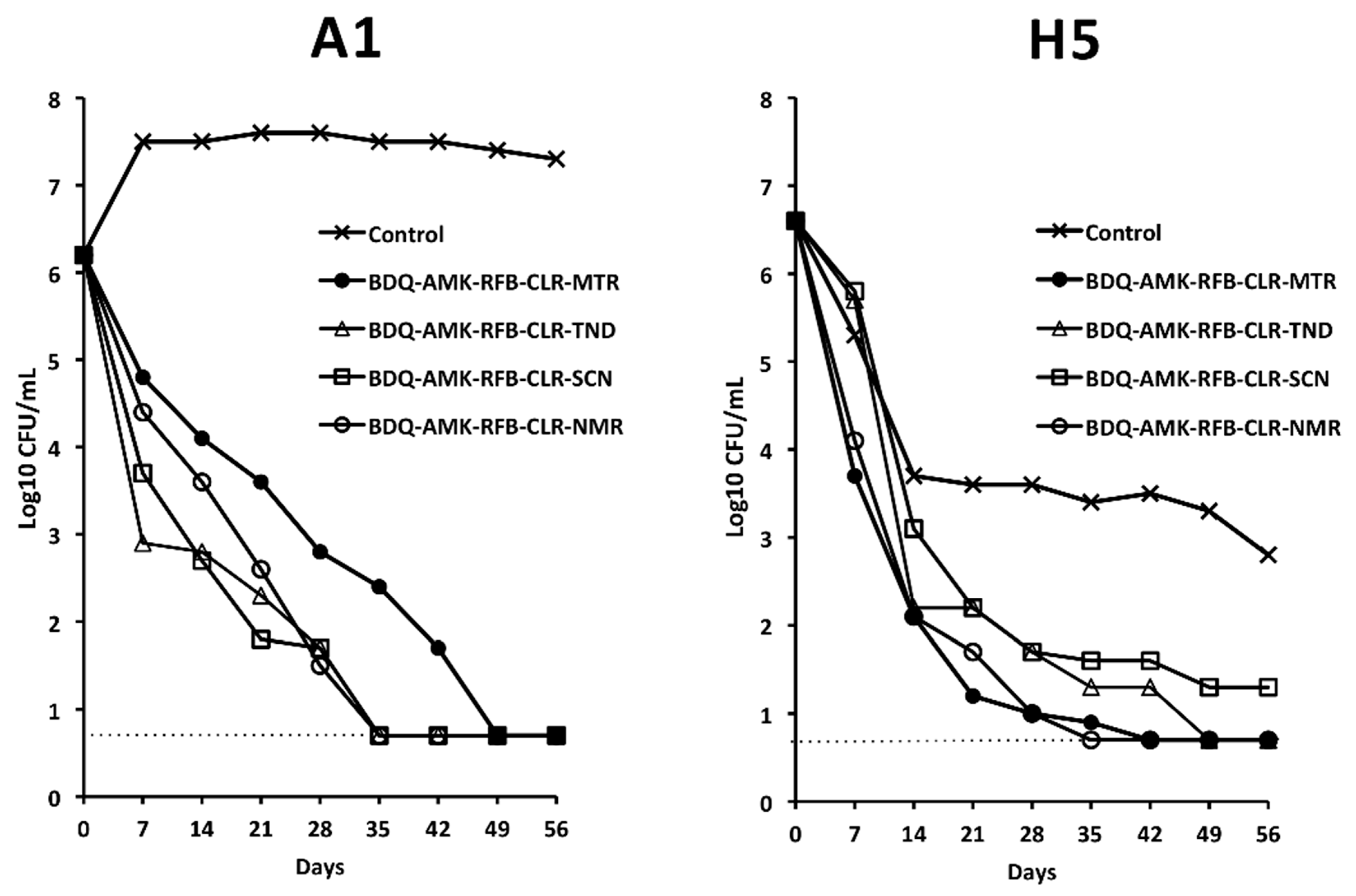
3. Results
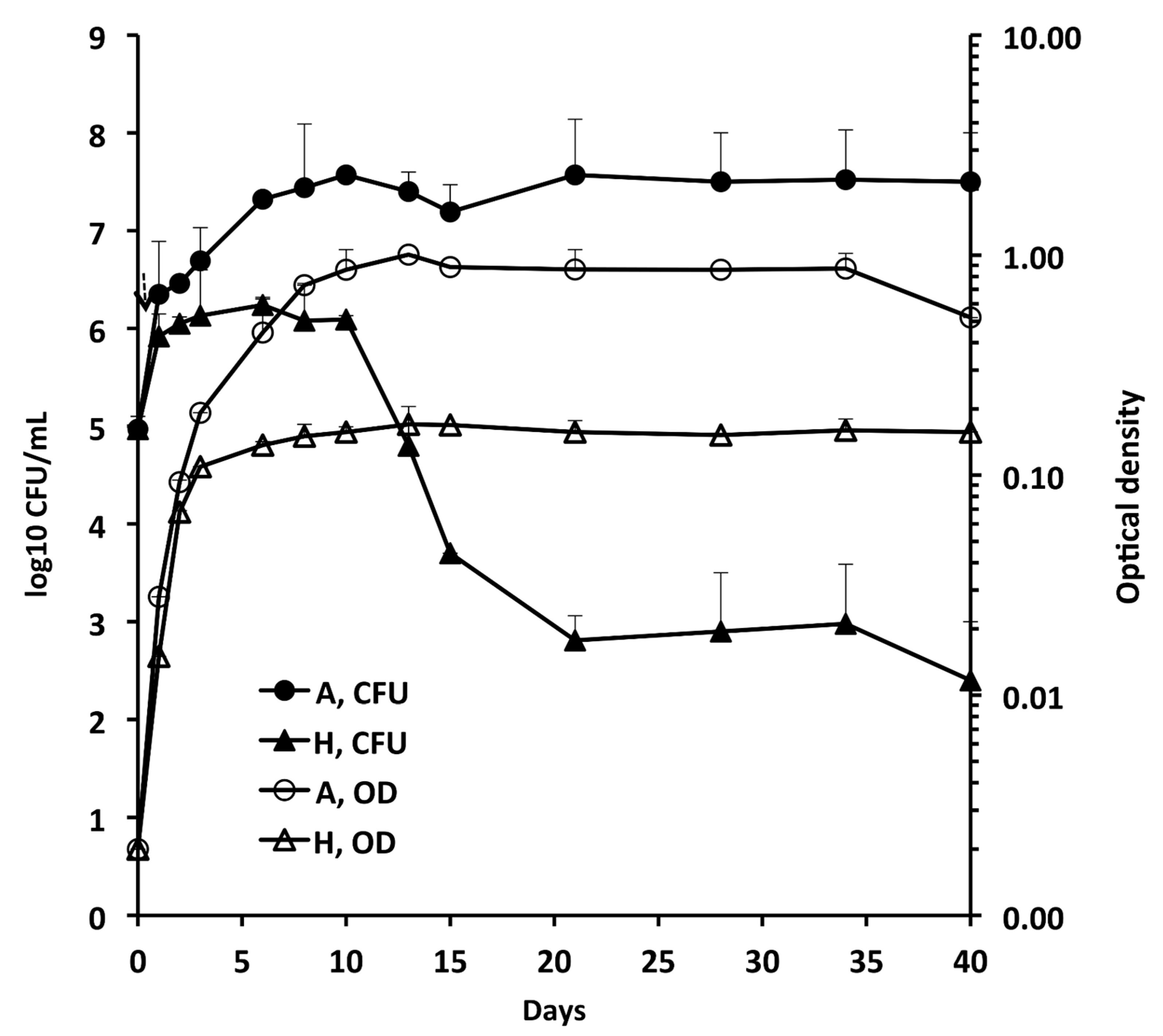
3.1. Growth of Mab-10 under A and H Conditions
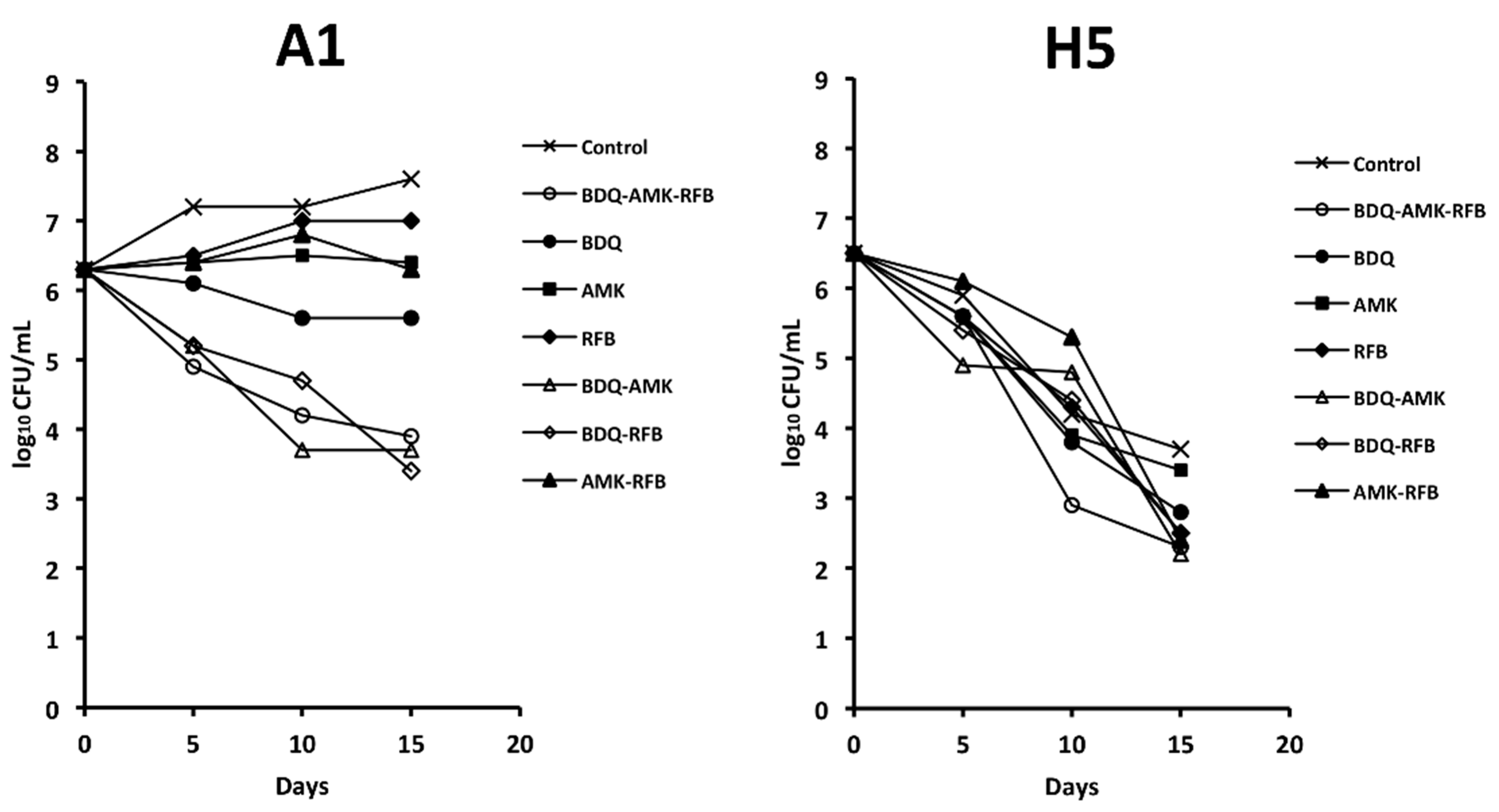
3.2. Activity of the Combination BDQ-AMK-RFB
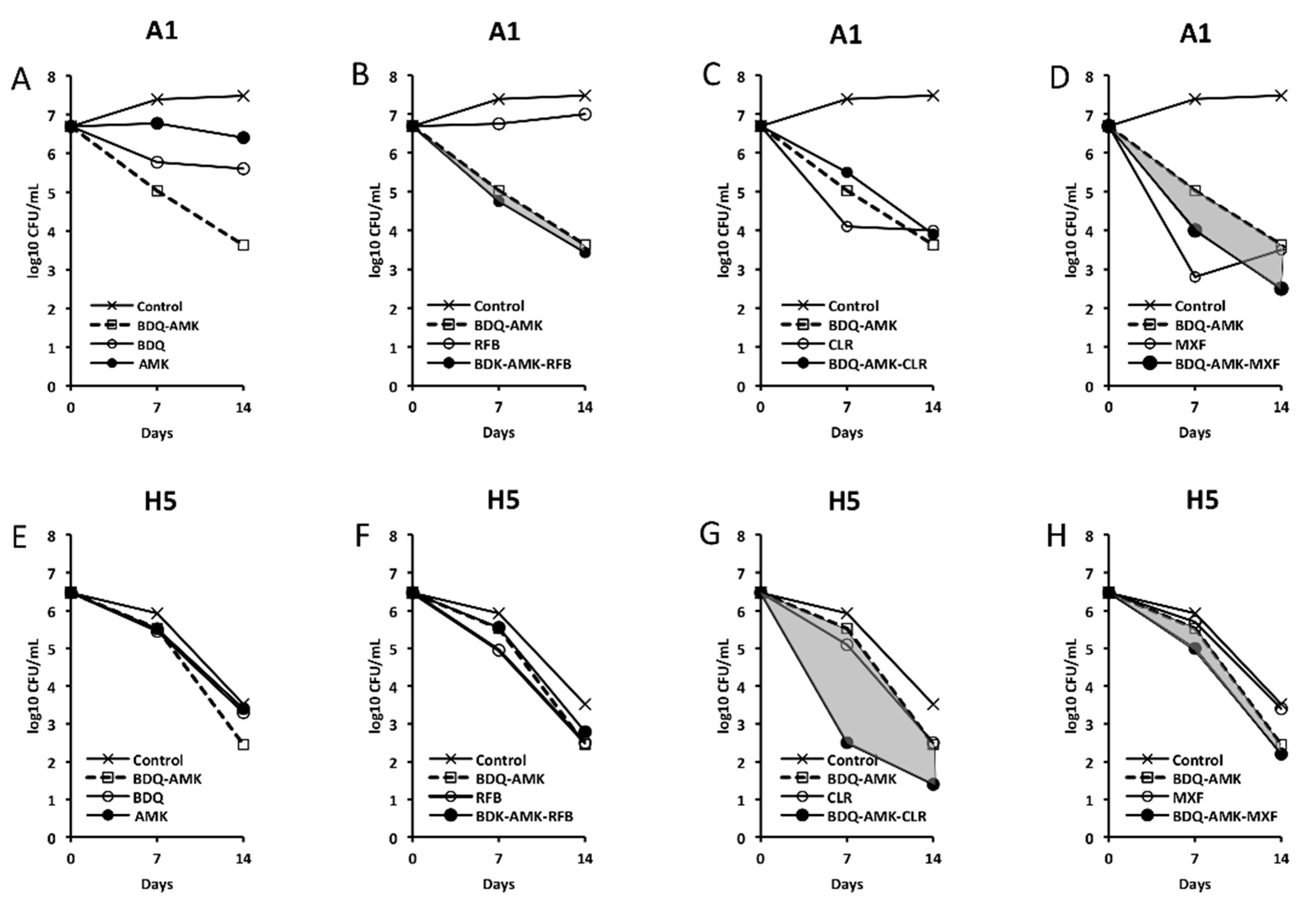
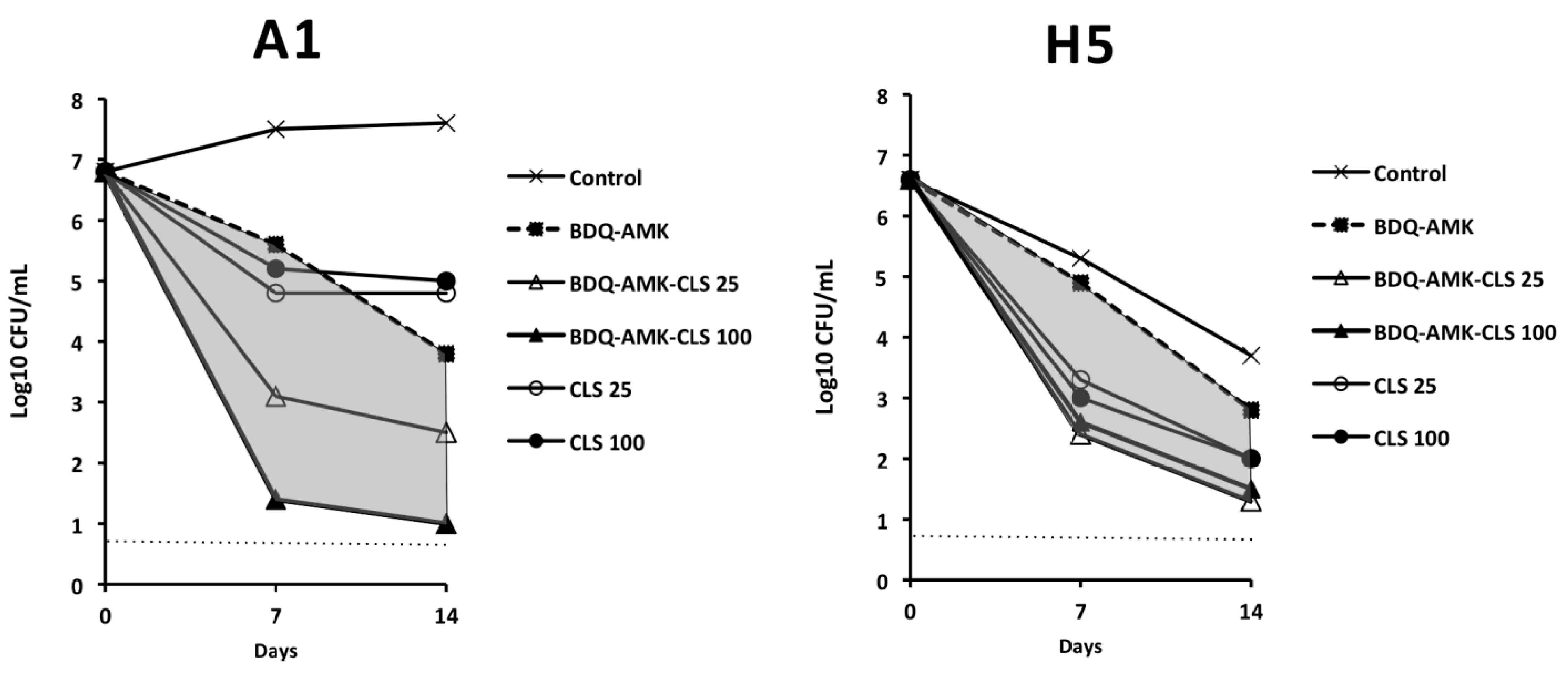
3.3. Activity of Three-Drug Combinations Containing BDQ-AMK
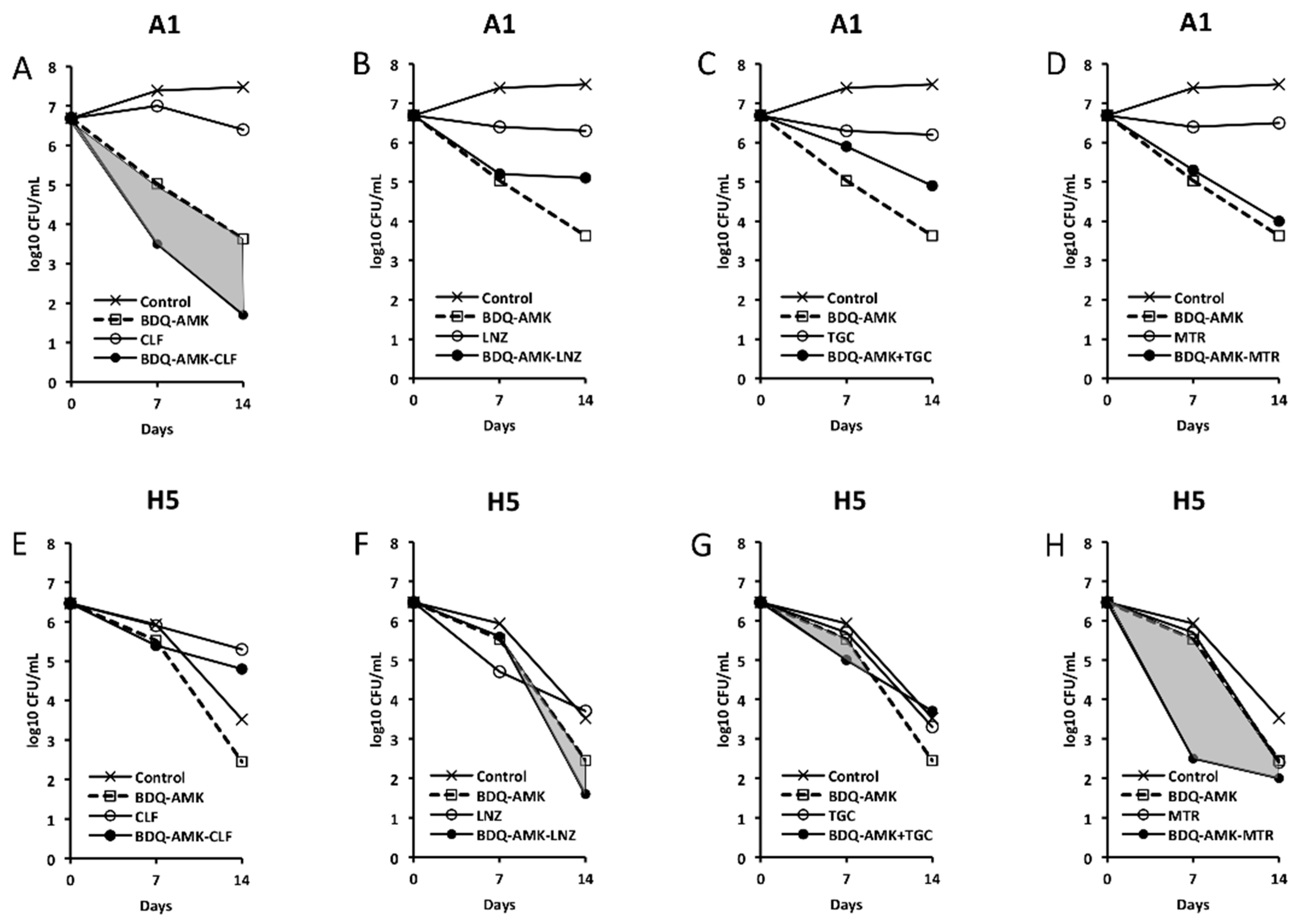
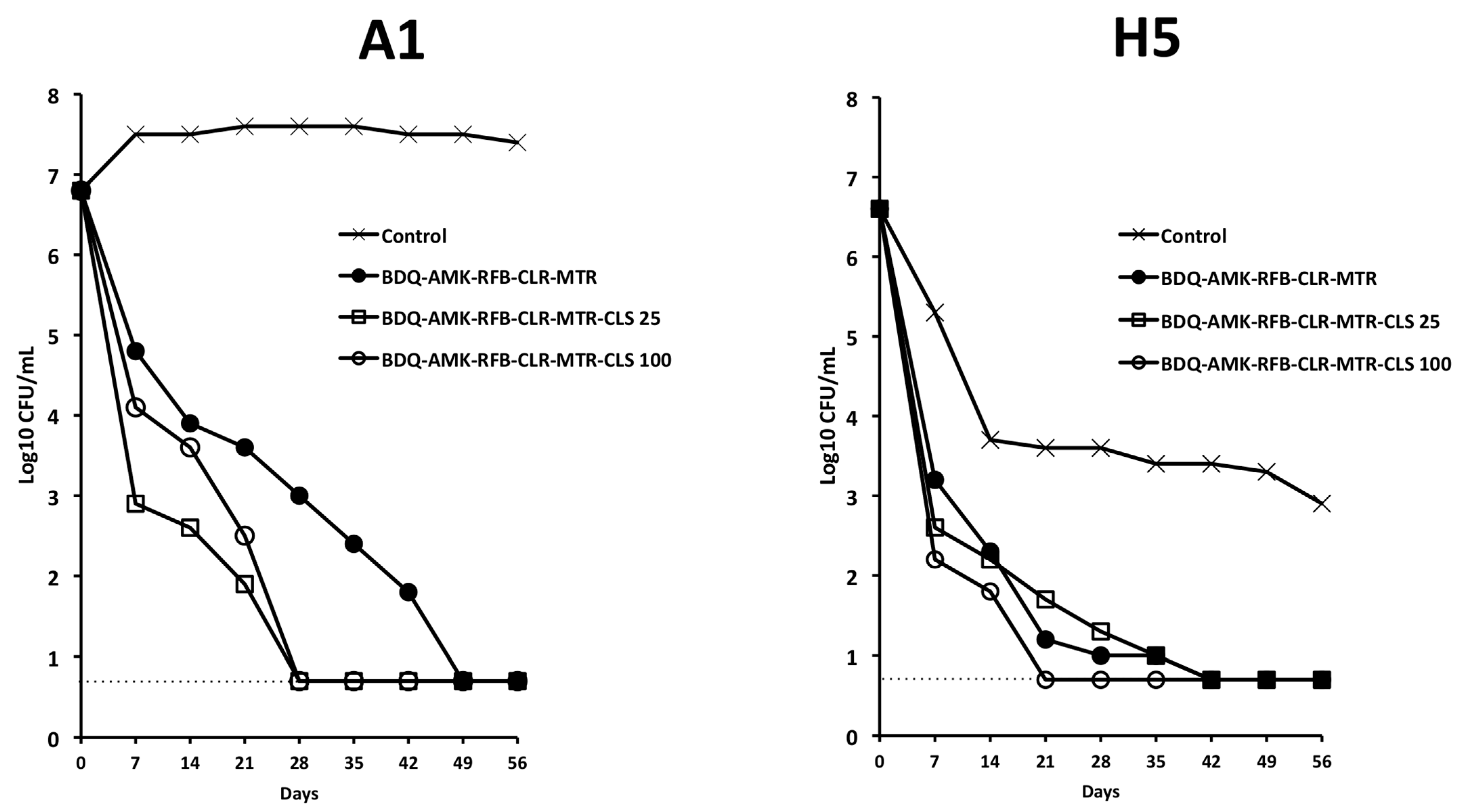
3.4. Activity of Five-Drug Combinations
3.5. Activity of BDQ-AMK-RFB-CLR-MTR plus CLS
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wu, M.L.; Aziz, D.B.; Dartois, V.; Dick, T. NTM drug discovery: Status, gaps and the way forward. Drug Discov. Today 2018, 23, 1502–1519. [Google Scholar] [CrossRef] [PubMed]
- Degiacomi, G.; Sammartino, J.C.; Chiarelli, L.R.; Riabova, O.; Makarov, V.; Pasca, M.R. Mycobacterium abscessus, an Emerging and Worrisome Pathogen among Cystic Fibrosis Patients. Int. J. Mol. Sci. 2019, 20, 5868. [Google Scholar] [CrossRef] [PubMed]
- Ryan, K.; Byrd, T.F. Mycobacterium abscessus: Shapeshifter of the Mycobacterial World. Front. Microbiol. 2018, 9, 2642. [Google Scholar] [CrossRef]
- Lopeman, R.C.; Harrison, J.; Desai, M.; Cox, J.A.G. Mycobacterium abscessus: Environmental Bacterium Turned Clinical Nightmare. Microorganisms 2019, 7, 90. [Google Scholar] [CrossRef]
- Johansen, M.D.; Herrmann, J.L.; Kremer, L. Non-tuberculous mycobacteria and the rise of Mycobacterium abscessus. Nat. Rev. Microbiol. 2020, 18, 392–407. [Google Scholar] [CrossRef] [PubMed]
- Brugha, R.; Spencer, H. Mycobacterium abscessus in cystic fibrosis. Science 2021, 372, 465–466. [Google Scholar] [CrossRef]
- Elborn, J.S. Cystic fibrosis. Lancet 2016, 388, 2519–2531. [Google Scholar] [CrossRef]
- Fennelly, K.P.; Ojano-Dirain, C.; Yang, Q.; Liu, L.; Lu, L.; Progulske-Fox, A.; Wang, G.P.; Antonelli, P.; Schultz, G. Biofilm Formation by Mycobacterium abscessus in a Lung Cavity. Am. J. Respir. Crit. Care Med. 2016, 193, 692–693. [Google Scholar] [CrossRef]
- Stewart, P.S.; White, B.; Boegli, L.; Hamerly, T.; Williamson, K.S.; Franklin, M.J.; Bothner, B.; James, G.A.; Fisher, S.; Vital-Lopez, F.G.; et al. Conceptual Model of Biofilm Antibiotic Tolerance That Integrates Phenomena of Diffusion, Metabolism, Gene Expression, and Physiology. J. Bacteriol. 2019, 201, e00307-19. [Google Scholar] [CrossRef]
- Yam, Y.K.; Alvarez, N.; Go, M.L.; Dick, T. Extreme Drug Tolerance of Mycobacterium abscessus “Persisters”. Front. Microbiol. 2020, 11, 359. [Google Scholar] [CrossRef]
- Clary, G.; Sasindran, S.J.; Nesbitt, N.; Mason, L.; Cole, S.; Azad, A.; McCoy, K.; Schlesinger, L.S.; Hall-Stoodley, L. Mycobacterium abscessus Smooth and Rough Morphotypes Form Antimicrobial-Tolerant Biofilm Phenotypes but Are Killed by Acetic Acid. Antimicrob. Agents Chemother. 2018, 62, e01782-17. [Google Scholar] [CrossRef] [PubMed]
- Bryant, J.M.; Grogono, D.M.; Rodriguez-Rincon, D.; Everall, I.; Brown, K.P.; Moreno, P.; Verma, D.; Hill, E.; Drijkoningen, J.; Gilligan, P.; et al. Emergence and spread of a human-transmissible multidrug-resistant nontuberculous mycobacterium. Science 2016, 354, 751–757. [Google Scholar] [CrossRef] [PubMed]
- Floto, R.A.; Olivier, K.N.; Saiman, L.; Daley, C.L.; Herrmann, J.L.; Nick, J.A.; Noone, P.G.; Bilton, D.; Corris, P.; Gibson, R.L.; et al. US Cystic Fibrosis Foundation and European Cystic Fibrosis Society. US Cystic Fibrosis Foundation and European Cystic Fibrosis Society consensus recommendations for the management of non-tuberculous mycobacteria in individuals with cystic fibrosis. Thorax 2016, 71 (Suppl. 1), i1–i22. [Google Scholar] [CrossRef] [PubMed]
- Ganapathy, U.S.; Dartois, V.; Dick, T. Repositioning rifamycins for Mycobacterium abscessus lung disease. Expert Opin. Drug Discov. 2019, 14, 867–878. [Google Scholar] [CrossRef]
- Gumbo, T.; Cirrincione, K.; Srivastava, S. Repurposing drugs for treatment of Mycobacterium abscessus: A view to a kill. J. Antimicrob. Chemother. 2020, 75, 1212–1217. [Google Scholar] [CrossRef]
- Le Moigne, V.; Raynaud, C.; Moreau, F.; Dupont, C.; Nigou, J.; Neyrolles, O.; Kremer, L.; Herrmann, J.L. Efficacy of Bedaquiline, Alone or in Combination with Imipenem, against Mycobacterium abscessus in C3HeB/FeJ Mice. Antimicrob. Agents Chemother. 2020, 64, e00114-20. [Google Scholar] [CrossRef]
- Meir, M.; Barkan, D. Alternative and Experimental Therapies of Mycobacterium abscessus Infections. Int. J. Mol. Sci. 2020, 21, 6793. [Google Scholar] [CrossRef]
- Lee, J.; Ammerman, N.; Agarwal, A.; Naji, M.; Li, S.Y.; Nuermberger, E. Differential In Vitro Activities of Individual Drugs and Bedaquiline-Rifabutin Combinations against Actively Multiplying and Nutrient-Starved Mycobacterium abscessus. Antimicrob. Agents Chemother. 2021, 65, e02179-20. [Google Scholar] [CrossRef]
- Nepali, K.; Lee, H.Y.; Liou, J.P. Nitro-Group-Containing Drugs. J. Med. Chem. 2019, 62, 2851–2893. [Google Scholar] [CrossRef]
- Olender, D.; Żwawiak, J.; Zaprutko, L. Multidirectional Efficacy of Biologically Active Nitro Compounds Included in Medicines. Pharmaceuticals 2018, 11, 54. [Google Scholar] [CrossRef]
- Piccaro, G.; Giannoni, F.; Filippini, P.; Mustazzolu, A.; Fattorini, L. Activities of drug combinations against Mycobacterium tuberculosis grown in aerobic and hypoxic acidic conditions. Antimicrob. Agents Chemother. 2013, 57, 1428–1433. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Piccaro, G.; Poce, G.; Biava, M.; Giannoni, F.; Fattorini, L. Activity of lipophilic and hydrophilic drugs against dormant and replicating Mycobacterium tuberculosis. J. Antibiot. 2015, 68, 711–714. [Google Scholar] [CrossRef] [PubMed]
- Iacobino, A.; Piccaro, G.; Giannoni, G.; Mustazzolu, A.; Fattorini, L. Fighting tuberculosis by drugs targeting nonreplicating Mycobacterium tuberculosis bacilli. Int. J. Mycobacteriol. 2017, 6, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Iacobino, A.; Piccaro, G.; Giannoni, F.; Mustazzolu, A.; Fattorini, L. Mycobacterium tuberculosis Is Selectively Killed by Rifampin and Rifapentine in Hypoxia at Neutral pH. Antimicrob. Agents Chemother. 2017, 61, e02296-16. [Google Scholar] [CrossRef] [PubMed]
- Iacobino, A.; Giannoni, F.; Pardini, M.; Piccaro, G.; Fattorini, L. The Combination Rifampin-Nitazoxanide, but Not Rifampin-Isoniazid-Pyrazinamide-Ethambutol, Kills Dormant Mycobacterium tuberculosis in Hypoxia at Neutral pH. Antimicrob. Agents Chemother. 2019, 63, e00273-19. [Google Scholar] [CrossRef] [PubMed]
- Mudde, S.E.; Upton, A.M.; Lenaerts, A.; Bax, H.I.; De Steenwinkel, J.E.M. Delamanid or pretomanid? A Solomonic judgement! J. Antimicrob. Chemother. 2022, 77, 880–902. [Google Scholar] [CrossRef]
- Tweed, C.D.; Dawson, R.; Burger, D.A.; Conradie, A.; Crook, A.M.; Mendel, C.M.; Conradie, F.; Diacon, A.H.; Ntinginya, N.E.; Everitt, D.E.; et al. Bedaquiline, moxifloxacin, pretomanid, and pyrazinamide during the first 8 weeks of treatment of patients with drug-susceptible or drug-resistant pulmonary tuberculosis: A multicentre, open-label, partially randomised, phase 2b trial. Lancet Respir. Med. 2019, 7, 1048–1058. [Google Scholar] [CrossRef]
- Conradie, F.; Diacon, A.H.; Ngubane, N.; Pauline Howell, P.; Everitt, D.; Crook, A.M.; Mendel, C.M.; Egizi, E.; Moreira, J.; Timm, J.; et al. Treatment of Highly Drug-Resistant Pulmonary Tuberculosis. N. Engl. J. Med. 2020, 382, 893–902. [Google Scholar] [CrossRef]
- Berube, B.J.; Castro, L.; Russell, D.; Ovechkina, Y.; Parish, T. Novel Screen to Assess Bactericidal Activity of Compounds Against Non-replicating Mycobacterium abscessus. Front. Microbiol. 2018, 9, 2417. [Google Scholar] [CrossRef]
- Mukherjee, T.; Boshoff, H.; Barry, C.E., 3rd. Comment on: Identification of antimicrobial activity among FDA-approved drugs for combating Mycobacterium abscessus and Mycobacterium chelonae. J. Antimicrob. Chemother. 2012, 67, 252–253. [Google Scholar] [CrossRef]
- Zhang, F.; Li, S.; Wen, S.; Zhang, T.; Shang, Y.; Huo, F.; Xue, Y.; Li, L.; Pang, Y. Comparison of in vitro Susceptibility of Mycobacteria Against PA-824 to Identify Key Residues of Ddn, the Deazoflavin-Dependent Nitroreductase from Mycobacterium tuberculosis. Infect. Drug Resist. 2020, 13, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. Performance Standards for Susceptibility Testing of Mycobacteria, Nocardia spp., and Other Aerobic Actinomycetes; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018; ISBN 978-1-68440-028-7. [Google Scholar]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Wayne, L.G.; Hayes, L.G. An in vitro model for sequential study of shiftdown of Mycobacterium tuberculosis through two stages of nonreplicating persistence. Infect. Immun. 1996, 64, 2062–2069. [Google Scholar] [CrossRef] [PubMed]
- Blaschke, T.F.; Skinner, M.H. The clinical pharmacokinetic of rifabutin. Clin. Infect. Dis. 1996, 22 (Suppl. 1), S15–S22. [Google Scholar] [CrossRef] [PubMed]
- Rodvold, K.A. Clinical Pharmacokinetics of Clarithromycin. Clin. Pharmacokinet. 1999, 37, 385–398. [Google Scholar] [CrossRef] [PubMed]
- Muralidharan, G.; Micalizzi, M.; Speth, J.; Raible, D.; Troy, S. Pharmacokinetics of tigecycline after single and multiple doses in healthy subjects. Antimicrob. Agents Chemother. 2005, 49, 220–229. [Google Scholar] [CrossRef]
- MacGowan, A.P. Pharmacokinetic and pharmacodynamic profile of linezolid in healthy volunteers and patients with Gram-positive infections. J. Antimicrob. Chemother. 2003, 51 (Suppl. S2), ii17–ii25. [Google Scholar] [CrossRef]
- Penkitis, H.; Adetoro, N. A review of the pharmacokinetic properties of SYM-1219, a single oral dose granule formulation containing 2 g of secnidazole, for the treatment of bacterial vaginosis. Am. J. Obstet. Gynecol. 2017, 217, 715–716. [Google Scholar] [CrossRef]
- Fung, H.B.; Doan, T.-L. Tinidazole: A nitroimidazole antiprotozoal agent. Clin. Ther. 2005, 27, 1859–1884. [Google Scholar] [CrossRef]
- Steib, A.; Jacoberger, B.; Von Bandel, M.; Beck, F.; Beller, J.P.; Boudjema, K.; Koffel, J.C.; Otteni, J.C. Concentrations in plasma and tissue penetration of ceftriaxone and ornidazole during liver transplantation. Antimicrob. Agents Chemother. 1993, 37, 1873–1876. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Overgaard, J.; Overgaard, M.; Timothy, A.R. Studies of the pharmacokinetic properties of nimorazole. Br. J. Cancer 1983, 48, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Montero, M.; Mir, M.; Sulleiro, E.; Avalos Esquivel, J.L.; García López, E.; Molina-Morant, D.; Montesinos, I.L.; Sorlí, L.; Veliz Espinosa, G.; Mounteis Oliva, E.; et al. High-dose benznidazole in a 62-year-old Bolivian kidney transplant recipient with Chagas central nervous system involvement. Int. J. Infect. Dis. 2019, 78, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Burock, D.; Daum, D.; Tröger, H.; Kim, T.D.; Krüger, S.; Rieke, D.T.; Ochsenreither, S.; Welter, K.; Herrmann, P.; Sleegers, A.; et al. Niclosamide a new chemotherapy agent? Pharmacokinetics of the potential anticancer drug in a patient cohort of the NIKOLO trial. J. Clin. Oncol. 2018, 36 (Suppl. 15), e14536. [Google Scholar] [CrossRef]
- Wijma, R.A.; Huttner, A.; Koch, B.C.P.; Mouton, J.W.; Muller, A.E. Review of the pharmacokinetic properties of nitrofurantoin and nitroxoline. J. Antimicrob. Chemother. 2018, 73, 2916–2926. [Google Scholar] [CrossRef]
- Schuster, A.; Haliburn, C.; Döring, G.; Goldman, M.H.; Freedom Study Group. Safety, efficacy and convenience of colistimethate sodium dry powder for inhalation (Colobreathe DPI) in patients with cystic fibrosis: A randomised study. Thorax 2013, 68, 344–350. [Google Scholar] [CrossRef]
- Yano, T.; Kassovska-Bratinova, S.; Teh, J.S.; Winkler, J.; Sullivan, K.; Isaacs, A.; Schechter, N.M.; Rubin, H. Reduction of clofazimine by mycobacterial type 2 NADH:quinone oxidoreductase: A pathway for the generation of bactericidal levels of reactive oxygen species. J. Biol. Chem. 2011, 286, 10276–10287. [Google Scholar] [CrossRef]
- Boldrin, F.; Provvedi, R.; Cioetto Mazzabò, L.; Segafreddo, G.; Manganelli, R. Tolerance and Persistence to Drugs: A Main Challenge in the Fight against Mycobacterium tuberculosis. Front. Microbiol. 2020, 11, 1924. [Google Scholar] [CrossRef]
- Singh, R.; Manjunatha, U.; Boshoff, H.I.; Ha, Y.H.; Niyomrattanakit, P.; Ledwidge, R.; Dowd, C.S.; Lee, I.Y.; Kim, P.; Zhang, L.; et al. PA-824 kills nonreplicating Mycobacterium tuberculosis by intracellular NO release. Science 2008, 322, 1392–1395. [Google Scholar] [CrossRef]
- Pamba, H.O.; Estambale, B.B.A.; Chunge, C.N.; Donno, L. Comparative study of aminosidine, etophamide and nimorazole, alone or in combination, in the treatment of intestinal amoebiasis in Kenya. Eur. J. Clin. Pharmacol. 1990, 9, 353–357. [Google Scholar] [CrossRef]
- Roy, R.B.; Laird, S.M.; Heasman, L. Treatment of trichomoniasis in the female. A comparison of metronidazole and nimorazole. Br. J. Vener Dis. 1975, 51, 281–284. [Google Scholar] [CrossRef] [PubMed]
- Metwally, M.A.H.; Jansen, J.A.; Overgaard, J. Study of the Population Pharmacokinetic Characteristics of Nimorazole in Head and Neck Cancer Patients Treated in the DAHANCA-5 Trial. Clin. Oncol. 2015, 27, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Maselli, D.J.; Keyt, H.; Restrepo, M.I. Inhaled Antibiotic Therapy in Chronic Respiratory Diseases. Int. J. Mol. Sci. 2017, 18, 1062. [Google Scholar] [CrossRef]
- Taccetti, G.; Francalanci, M.; Pizzamiglio, G.; Messore, B.; Carnovale, V.; Cimino, G.; Cipolli, M. Cystic Fibrosis: Recent Insights into Inhaled Antibiotic Treatment and Future Perspectives. Antibiotics 2021, 10, 338. [Google Scholar] [CrossRef]
- Bax, H.I.; de Steenwinkel, J.E.M.; ten Kate, M.T.; van der Meijden, A.; Verbon, A.; Bakker-Woudenberg, I.A.J.M. Colistin as a potentiator of anti-TB drug activity against Mycobacterium tuberculosis. J. Antimicrob. Chemother. 2015, 70, 2828–2837. [Google Scholar] [CrossRef]
- Koen, N.; van Breda, S.V.; Loots, D.T. Elucidating the antimicrobial mechanisms of colistin sulfate on Mycobacterium tuberculosis using metabolomics. Tuberculosis 2018, 111, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Salcedo-Sora, J.E.; Kell, D.B. A Quantitative Survey of Bacterial Persistence in the Presence of Antibiotics: Towards Antipersister Antimicrobial Discovery. Antibiotics 2020, 9, 508. [Google Scholar] [CrossRef]
- Kaur, A.; Sharma, P.; Capalash, N. Curcumin alleviates persistence of Acinetobacter baumannii against colistin. Sci. Rep. 2018, 8, 11029. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A1 | H5 | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DTP ± SD by Days of Drug Exposure | DTP ± SD by Days of Drug Exposure | |||||||||||||||
| Combination | 7 | 14 | 21 | 28 | 35 | 42 | 49 | 56 | 7 | 14 | 21 | 28 | 35 | 42 | 49 | 56 |
| Control | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 ± 1 | 3 ± 1 | 3 ± 1 | 3 ± 1 | 3 ± 2 | 3 ± 2 | 4 ± 1 |
| BDQ-AMK-RFB-CLR-MTR | 2 | 2 ± 1 | 3 ± 1 | 3 ± 1 | 4 ± 1 | 5 ± 1 | 7 ± 2 | 7 ± 3 | 3 ± 1 | 5 ± 1 | 6 ± 1 | 6 ± 2 | 6 ± 2 | 7 ± 3 | 7 ± 2 | >50 |
| BDQ-AMK-RFB-CLR-TND | 3 ± 1 | 3 ± 1 | 4 ± 1 | 4 ± 2 | 20 ± 5 | >50 | >50 | >50 | 2 ± 1 | 4 ± 1 | 4 ± 1 | 5 ± 1 | 5 ± 1 | 6 ± 2 | 7 ± 2 | 7 ± 4 |
| BDQ-AMK-RFB-CLR-SCN | 2 ± 1 | 3 ± 1 | 4 ± 1 | 6 ± 1 | 28 ± 4 | >50 | >50 | >50 | 2 ± 1 | 3 ± 1 | 4 ± 1 | 5 ± 1 | 5 ± 1 | 6 ± 1 | 7 ± 4 | 7 ± 3 |
| BDQ-AMK-RFB-CLR-NMR | 2 | 4 ± 1 | 5 ± 1 | 6 ± 1 | 25 ± 2 | >50 | >50 | >50 | 2 ± 1 | 5 ± 1 | 5 ± 1 | 6 ± 1 | 7 ± 3 | >50 | >50 | >50 |
| A1 | H5 | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DTP ± SD by Days of Drug Exposure | DTP ± SD by Days of Drug Exposure | |||||||||||||||
| Combination | 7 | 14 | 21 | 28 | 35 | 42 | 49 | 56 | 7 | 14 | 21 | 28 | 35 | 42 | 49 | 56 |
| Control | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 3 ± 1 | 3 ± 1 | 3 ± 1 | 3 ± 2 | 3 ± 1 | 3 ± 1 | 4 ± 1 |
| BDQ-AMK-RFB-CLR-MTR | 1 | 2 ± 1 | 2 ± 1 | 3 ± 1 | 4 ± 1 | 4 ± 1 | 6 ± 1 | 7 ± 2 | 3 ± 1 | 4 ± 1 | 5 ± 1 | 6 ± 1 | 6 ± 1 | 7 ± 2 | 7 ± 3 | >50 |
| BDQ-AMK-RFB-CLR-MTR-CLS 25 | 4 ± 1 | 4 ± 1 | 4 ± 2 | 13 ± 1 | 13 ± 5 | 28 ± 5 | 28 ± 4 | >50 | 3 ± 1 | 4 ± 1 | 5 ± 1 | 5 ± 1 | 5 ± 1 | 6 ± 1 | 7 ± 3 | 7 ± 2 |
| BDQ-AMK-RFB-CLR-MTR-CLS 100 | 2 ± 1 | 3 ± 1 | 4 ± 1 | 27 ± 4 | 27 ± 4 | >50 | >50 | >50 | 5 ± 1 | 5 ± 2 | 6 ± 1 | 7 ± 3 | 7 ± 2 | 7 ± 2 | 19 ± 4 | >50 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lanni, A.; Borroni, E.; Iacobino, A.; Russo, C.; Gentile, L.; Fattorini, L.; Giannoni, F. Activity of Drug Combinations against Mycobacterium abscessus Grown in Aerobic and Hypoxic Conditions. Microorganisms 2022, 10, 1421. https://doi.org/10.3390/microorganisms10071421
Lanni A, Borroni E, Iacobino A, Russo C, Gentile L, Fattorini L, Giannoni F. Activity of Drug Combinations against Mycobacterium abscessus Grown in Aerobic and Hypoxic Conditions. Microorganisms. 2022; 10(7):1421. https://doi.org/10.3390/microorganisms10071421
Chicago/Turabian StyleLanni, Alessio, Emanuele Borroni, Angelo Iacobino, Cristina Russo, Leonarda Gentile, Lanfranco Fattorini, and Federico Giannoni. 2022. "Activity of Drug Combinations against Mycobacterium abscessus Grown in Aerobic and Hypoxic Conditions" Microorganisms 10, no. 7: 1421. https://doi.org/10.3390/microorganisms10071421
APA StyleLanni, A., Borroni, E., Iacobino, A., Russo, C., Gentile, L., Fattorini, L., & Giannoni, F. (2022). Activity of Drug Combinations against Mycobacterium abscessus Grown in Aerobic and Hypoxic Conditions. Microorganisms, 10(7), 1421. https://doi.org/10.3390/microorganisms10071421