
Differentiation of Laboratory-Obtained Ixodes ricinus × Ixodes persulcatus Hybrid Ticks: Selection of Suitable Genes

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ticks
2.2. DNA Isolation
2.3. PCR
2.4. PCR Product Purification, TA-Cloning, and Plasmid Purification and Sequencing
2.5. qPCR
2.6. Bioinformatics
3. Results
3.1. Testing ITS Primer Sets
3.2. Searching for Prospective Genes for Hybrid Determination
3.3. Testing Lysozyme and Toll_R Amplicons as a Hybrid Differentiator
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-Peña, A.; George, J.C.; Golovljova, I.; Jaenson, T.G.T.; Jensen, J.K.; Jensen, P.M.; et al. Driving forces for changes in geographical distribution of Ixodes ricinus ticks in Europe. Parasites Vectors 2013, 6, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bugmyrin, S.V.; Bespyatova, L.A.; Korotkov, Y.S.; Burenkova, L.A.; Belova, O.A.; Iu, L.; Kozlovskaya, L.I.; Karganova, G.G.; Ieshko, E.P. Distribution of Ixodes ricinus and I. persulcatus ticks in southern Karelia (Russia). Ticks Tick-Borne Dis. 2013, 4, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Sormunen, J.J.; Andersson, T.; Aspi, J.; Bäck, J.; Cederberg, T.; Haavisto, N.; Halonen, H.; Hänninen, J.; Inkinen, J.; Kulha, N.; et al. Monitoring of ticks and tick-borne pathogens through a nationwide research station network in Finland. Ticks Tick-Borne Dis. 2020, 11, 101449. [Google Scholar] [CrossRef] [PubMed]
- Balashov, Y.; Grigoreva, L.; Oliver, J. Reproductive isolation and interspecific hybridization of ixodid ticks of the Ixodes ricinus—I. persulcatus group (Acarina Ixodidae). Entomol. Rev. 1998, 78, 500–508. [Google Scholar]
- Ciota, A.T.; Chin, P.A.; Kramer, L.D. The effect of hybridization of Culex pipiens complex mosquitoes on transmission of West Nile virus. Parasites Vectors 2013, 6, 2–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bugmyrin, S.V.; Belova, O.A.; Ieshko, E.P.; Bespyatova, L.A.; Karganova, G.G. Morphological differentiation of Ixodes persulcatus and I. ricinus hybrid larvae in experiment and under natural conditions. Ticks Tick-Borne Dis. 2015, 6, 129–133. [Google Scholar] [CrossRef]
- Bugmyrin, S.V.; Belova, O.A.; Bespyatova, L.A.; Ieshko, E.P.; Karganova, G.G. Morphological features of Ixodes persulcatus and I. ricinus hybrids: Nymphs and adults. Exp. Appl. Acarol. 2016, 69, 359–369. [Google Scholar] [CrossRef]
- Coleman, A.W. ITS2 is a double-edged tool for eukaryote evolutionary comparisons. Trends Genet. 2003, 19, 370–375. [Google Scholar] [CrossRef]
- Barker, S.C. Distinguishing Species and Populations of Rhipicephaline Ticks with ITS 2 Ribosomal RNA. J. Parasitol. 1998, 84, 887–892. [Google Scholar] [CrossRef]
- Van Zee, J.; Piesman, J.F.; Hojgaard, A.; Cormack, W.B. Nuclear Markers Reveal Predominantly North to South Gene Flow in Ixodes scapularis, the Tick Vector of the Lyme Disease Spirochete. PLoS ONE 2015, 10, e139630. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, J.M.; Goddard, J.; Rasgon, J.L. Population and Demographic Structure of Ixodes scapularis Say in the Eastern United States. PLoS ONE 2014, 9, e101389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovalev, S.Y.; Golovljova, I.V.; Mukhacheva, T.A. Natural hybridization between Ixodes ricinus and Ixodes persulcatus ticks evidenced by molecular genetics methods. Ticks Tick-Borne Dis. 2016, 7, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Kovalev, S.Y.; Mikhaylishcheva, M.S.; Mukhacheva, T.A. Natural hybridization of the ticks Ixodes persulcatus and Ixodes pavlovskyi in their sympatric populations in Western Siberia. Infect. Genet. Evol. 2015, 32, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Patterson, J.W.; Duncan, A.M.; Mcintyre, K.C.; Lloyd, V.K. Evidence for genetic hybridization between Ixodes scapularis and Ixodes cookei. Can. J. Zool. 2017, 537, 527–537. [Google Scholar] [CrossRef] [Green Version]
- Rar, V.; Livanova, N.; Sabitova, Y.; Igolkina, Y.; Tkachev, S.; Tikunov, A.; Babkin, I.; Golovljova, I.; Panov, V.; Tikunova, N. Ixodes persulcatus/pavlovskyi natural hybrids in Siberia: Occurrence in sympatric areas and infection by a wide range of tick-transmitted agents. Ticks Tick-Borne Dis. 2019, 10, 101254. [Google Scholar] [CrossRef]
- Araya-Anchetta, A.; Scoles, G.A.; Giles, J.; Busch, J.D.; Wagner, D.M. Hybridization in natural sympatric populations of Dermacentor ticks in northwestern North America. Ecol. Evol. 2013, 3, 714–724. [Google Scholar] [CrossRef]
- Kovalev, S.Y.; Mukhacheva, T.A. An improved real-time PCR method to identify hybrids between Ixodes persulcatus and Ixodes ricinus ticks. Ticks Tick-Borne Dis. 2018, 9, 37–38. [Google Scholar] [CrossRef]
- Belova, O.A.; Burenkova, L.A.; Karganova, G.G. Different tick-borne encephalitis virus (TBEV) prevalences in unfed versus partially engorged ixodid ticks—Evidence of virus replication and changes in tick behavior. Ticks Tick-Borne Dis. 2012, 3, 240–246. [Google Scholar] [CrossRef]
- Kholodilov, I.S.; Belova, O.A.; Morozkin, E.S.; Litov, A.G.; Ivannikova, A.Y.; Makenov, M.T.; Shchetinin, A.M.; Aibulatov, S.V.; Bazarova, G.K.; Bell-sakyi, L.; et al. Geographical and Tick-Dependent Distribution of Flavi-Like Alongshan and Yanggou Tick Viruses in Russia. Viruses 2021, 13, 458. [Google Scholar] [CrossRef]
- Filippova, N.A. Ixodid Ticks of the Subfamily Ixodinae. Fauna of the USSR: Arachnoides. Nauk. Leningr. 1977, 4, 396. [Google Scholar]
- Fukunaga, M.; Yabuki, M.; Hamase, A.; Oliver, J.H.; Nakao, M. Molecular phylogenetic analysis of ixodid ticks based on the ribosomal DNA spacer, internal transcribed spacer 2, sequences. J. Parasitol. 2000, 86, 38–43. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Munderloh, U.G.; Kurtti, T.J. Cytogenetic Characteristics of Cell Lines from Ixodes scapularis (Acari: Ixodidae). J. Med. Entomol. 1994, 31, 425–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, J.H. Cytogenetics of mites and ticks. Ann. Rev. Entomol. 1977, 22, 407–429. [Google Scholar] [CrossRef] [PubMed]
- Gulia-nuss, M.; Nuss, A.B.; Meyer, J.M.; Sonenshine, D.E.; Roe, R.M.; Waterhouse, R.M.; Sattelle, D.B.; de la Fuente, J.; Ribeiro, J.M.; Megy, K.; et al. Genomic insights into the Ixodes scapularis tick vector of Lyme disease. Nat. Commun. 2016, 7, 10507. [Google Scholar] [CrossRef] [Green Version]
- Lv, J.; Wu, S.; Zhang, Y.; Chen, Y.; Feng, C.; Yuan, X.; Jia, G.; Deng, J.; Wang, C.; Wang, Q.; et al. Assessment of four DNA fragments (COI, 16S rDNA, ITS2, 12S rDNA) for species identification of the Ixodida (Acari: Ixodida). Parasites Vectors 2014, 7, 93. [Google Scholar] [CrossRef] [Green Version]
- Long, E.O.; Dawid, I.B. Repeated genes in eukaryotes. Ann. Rev. Biochem. 1980, 49, 727–764. [Google Scholar] [CrossRef]
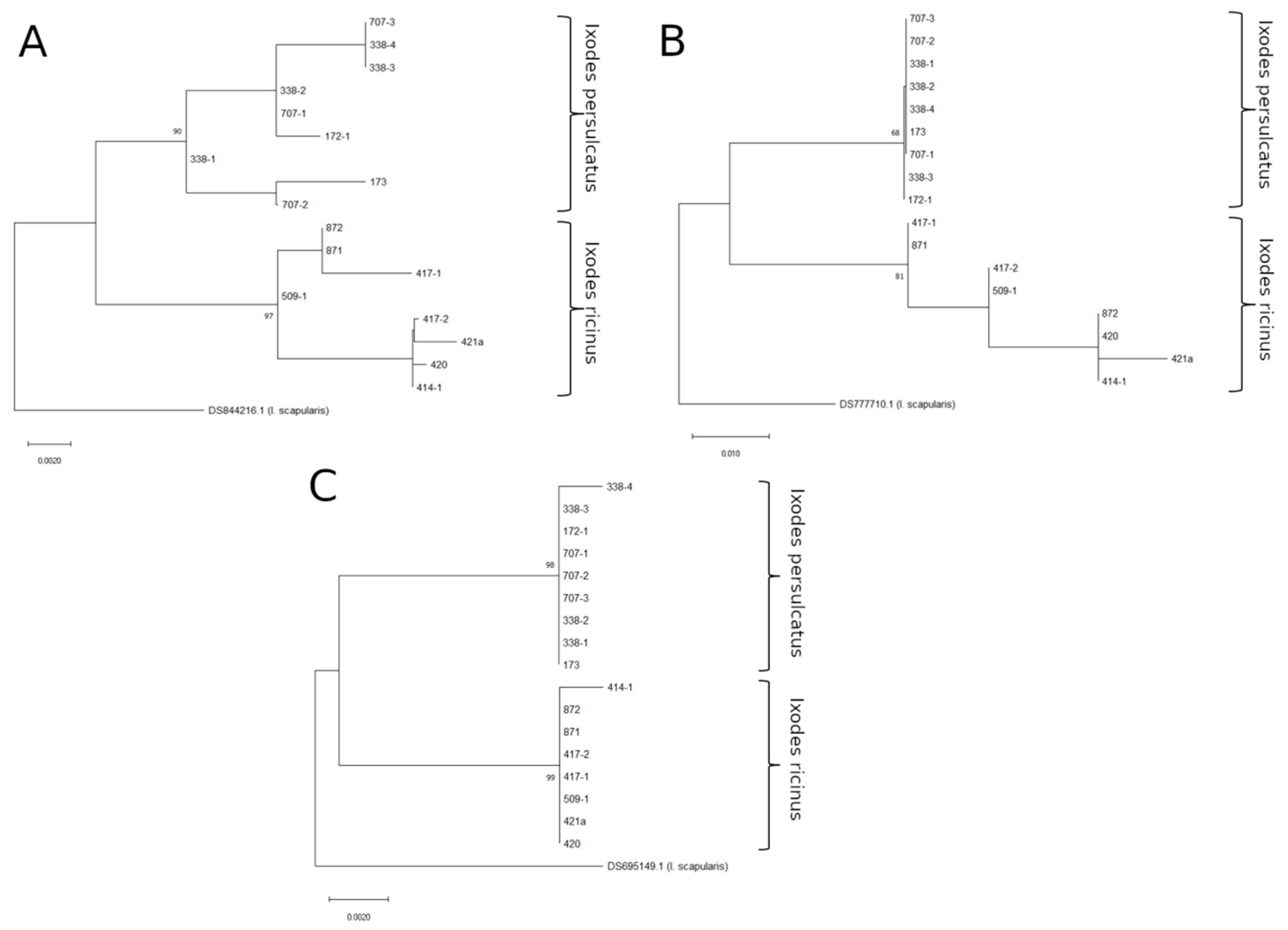

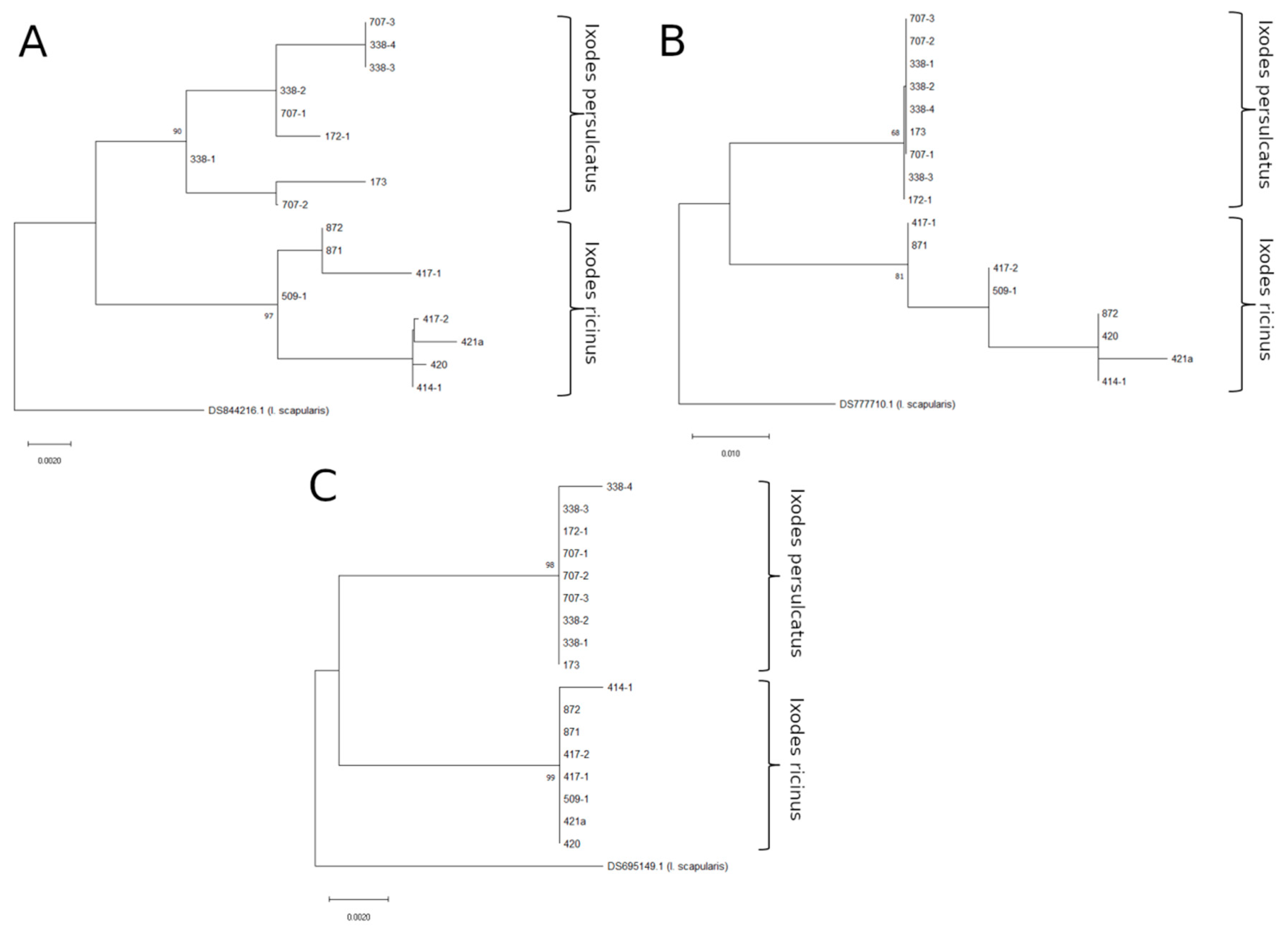{kind=link}
| Tick Specimen № | Tick Species | Sex | Collection Place | GPS |
|---|---|---|---|---|
| 172-1 | Ixodes persulcatus | ♀ | Russia, Republic of Tuva | 51.9923° 94.0842° |
| 173 | Ixodes persulcatus | ♀ | 51.6283° 94.4292° | |
| 338-1 | Ixodes persulcatus | ♂ | Russia, Republic of Karelia | 62.0586° 33.9427° |
| 338-2 | Ixodes persulcatus | ♂ | ||
| 338-3 | Ixodes persulcatus | ♂ | ||
| 338-4 | Ixodes persulcatus | ♂ | ||
| 707-1 | Ixodes persulcatus | ♀ | Russia, Republic of Karelia | 62.0747° 33.9495° |
| 707-2 | Ixodes persulcatus | ♀ | ||
| 707-3 | Ixodes persulcatus | ♀ | ||
| 414-1 | Ixodes ricinus | ♀ | Russia, Voronezh region | 51.7801° 39.4085° |
| 420 | Ixodes ricinus | ♀ | ||
| 509-1 | Ixodes ricinus | ♂ | ||
| 421a | Ixodes ricinus | ♂ | Russia, Voronezh region | 51.7850° 39.4054° |
| 417-1 | Ixodes ricinus | ♂ | Russia, Kaliningrad region | 55.1839° 20.8572° |
| 417-2 | Ixodes ricinus | ♂ | ||
| 871 | Ixodes ricinus | ♀ | Russia, Kaliningrad region | 55.1585° 20.8437° |
| 872 | Ixodes ricinus | ♀ |
| Primer Set Name | Oligonucleotide | Oligonucleotide Sequence | Primer Set Temperature | Reference | Amplicon Length |
|---|---|---|---|---|---|
| FITSMR-3SA | FITSMR | 5′-ccagtattcatcggggacgc-3′ | 55 °C | [9,21] | ≈660 |
| 3SA | 5′-ctaagcggtggatcactcgg-3′ | ||||
| JB9A-3SA | JB9A | 5′-gcactatcaagcaacacgactc-3′ | 52 °C | [9] | ≈1100 |
| 3SA | 5′-ctaagcggtggatcactcgg-3′ | ||||
| Lysozyme | Lys_eSNP1F | 5′-tgtctttggcttggatcgtc-3′ | 55 °C | [11] | 512 |
| Lys_eSNP1R | 5′-attcttccacctgccctacg-3′ | ||||
| Serpin 2 | Serp2Ae1-6_F | 5′-ttacgctcccgacgttattc-3′ | 50–55 °C * | [11] | 651 |
| Serp2Ae1-6_R | 5′-ttcgagggatcaaacaggtc-3′ | ||||
| Ixoderin | IxodBe2-3F | 5′-acacgtatgcctcaaagtgg-3′ | 50–55 °C * | [11] | 502 |
| IxodBe2-3R | 5′-gcactatatccagcgggaag-3′ | ||||
| Actin | Act_E6_F | 5′-ggagcacccgctccaggta-3′ | 55 °C | - ** | 268 |
| Act_E6_R | 5′-ctgtaattgcagctgccggac-3′ | ||||
| OdorR | OdorR_F | 5′-attccccacatcgcgcaa-3′ | 50–55 °C * | - | 536 |
| OdoR_R | 5′-ccctcatttctcagattctagcg-3′ | ||||
| JH_Met | JH_MT_F | 5′-gtctctaaacagatggtcgact-3′ | 52 °C | - | 308 |
| JH_MT_R | 5′-agactcacctcggcgta-3′ | ||||
| Toll_full | Toll_F | 5′-ctcgaacgtcatgaagact-3′ | 50 °C | - | 1403 |
| Toll_end_R | 5′-acttttgcggctatctgtttat-3′ | ||||
| Toll_L | Toll_F | 5′-ctcgaacgtcatgaagact-3′ | 50 °C | - | 811 |
| Toll_mid_R | 5′-ttcgatccagttgttacccaggct-3′ | ||||
| Toll_R | Toll_mid_F | 5′-gcgattgaatctctggaggg-3′ | 50 °C | - | 739 |
| Toll_end_R | 5′-acttttgcggctatctgtttat-3′ |
| Tick Specimen No | Sex | Tick Species | Ct I. persulcatus | Ct I. ricinus | Ct Delta | Tick Species (as Determined by Kovalev et al.’s System) |
|---|---|---|---|---|---|---|
| h604 | ♀ | hybrid ♀ I.persulcatus × ♂ I.ricinus | 15.93 | 15.49 | 0.44 | Hybrid 1:1 |
| h605 | ♀ | hybrid ♀ I.persulcatus × ♂ I.ricinus | 17.24 | 18.27 | 1.03 | Hybrid 1:2 |
| h606 | ♂ | hybrid ♀ I.persulcatus × ♂ I.ricinus | 23.00 | nd * | - ** | I. persulcatus |
| h730 | ♂ | hybrid ♀ I.persulcatus × ♂ I.ricinus | 15.05 | nd | - | I. persulcatus |
| h422-1 | ♀ | hybrid ♀ I.ricinus × ♂ I.persulcatus | 20.73 | 22.51 | 1.78 | Hybrid 1:4 |
| h422-2 | ♀ | hybrid ♀ I.ricinus × ♂ I.persulcatus | 17.79 | 18.99 | 1.20 | Hybrid 1:2 |
| h423-1 | ♂ | hybrid ♀ I.ricinus × ♂ I.persulcatus | 29.25 | 19.17 | 10.08 | Hybrid 1:∞ |
| h423-2 | ♂ | hybrid ♀ I.ricinus × ♂ I.persulcatus | 30.62 | 20.10 | 10.52 | Hybrid 1:∞ |
| 707-2 | ♀ | I. persulcatus | 14.33 | nd | - | I. persulcatus |
| 707-3 | ♀ | I. persulcatus | 15.05 | nd | - | I. persulcatus |
| 338-1 | ♂ | I. persulcatus | 15.09 | nd | - | I. persulcatus |
| 173 | ♀ | I. persulcatus | 14.95 | nd | - | I. persulcatus |
| 872 | ♀ | I. ricinus | nd | 15.20 | - | I. ricinus |
| 417-2 | ♂ | I. ricinus | nd | 16.88 | - | I. ricinus |
| 421a | ♂ | I. ricinus | nd | 17.01 | - | I. ricinus |
| 414-1 | ♀ | I. ricinus | nd | 14.60 | - | I. ricinus |
| Crossing | ♀ I. persulcatus × ♂ I. ricinus | ♀ I. ricinus × ♂ I. persulcatus | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Individual tick | h604 ♀ | h605 ♀ | h606 ♂ | h730 ♂ | h422-1 ♀ | h422-2 ♀ | h423-1 ♂ | h423-2 ♂ | |
| Clones from FITSMR-3SA ratio | Individual | 6:0 * | 4:2 | 6:0 * | 10:0 * | 10:0 * | 7:1 | 0:9 * | 1:8 |
| Total by sex | 10:2 | 16:0 * | 17:1 * | 1:17 * | |||||
| Clones from JB9A-3SA ratio | Individual | 8:2 | 5:5 | 10:0 * | 6:0 * | 6:0 * | 5:0 | 0:6 * | 0:5 |
| Total by sex | 13:7 | 16:0 * | 11:0 * | 0:11 * | |||||
| Crossing | Both ♀ I. persulcatus × ♂ I. ricinus and ♀ I. ricinus × ♂ I. persulcatus | |
|---|---|---|
| FITSMR-3SA ITS in | ||
| ♀ | ♂ | |
| I. persulcatus:I. ricinus ratio | 27:3 * | 17:17 |
| maternal:paternal ratio | 11:19 † | 33:1 # |
| JB9A-3SA ITS in | ||
| ♀ | ♂ | |
| I. persulcatus:I. ricinus ratio | 24:7 * | 16:11 |
| maternal:paternal ratio | 13:18 † | 27:0 # |
| Total ITS in | ||
| ♀ | ♂ | |
| maternal:paternal ratio | 24:37 † | 60:1 # |
| Crossing | ♀ I. persulcatus × ♂ I. ricinus | ♀ I. ricinus × ♂ I. persulcatus | |||
|---|---|---|---|---|---|
| Individual tick | h606 ♂ | h730 ♂ | h423-1 ♂ | h423-2 ♂ | |
| Clones from Lysozyme ratio | Individual | 1:4 | 0:5 | 0:5 | 0:4 |
| Total | 1:18 * | ||||
| Lysozyme total maternal:paternal ratio | 10:9 ‡ | ||||
| Clones from Toll_R ratio | Individual | 3:2 | 4:1 | 2:3 | 3:2 |
| Total | 12:8 | ||||
| Toll_R total maternal:paternal ratio | 12:8 ‡ | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Litov, A.G.; Belova, O.A.; Bugmyrin, S.V.; Kholodilov, I.S.; Romanova, L.I.; Karganova, G.G. Differentiation of Laboratory-Obtained Ixodes ricinus × Ixodes persulcatus Hybrid Ticks: Selection of Suitable Genes. Microorganisms 2022, 10, 1306. https://doi.org/10.3390/microorganisms10071306
Litov AG, Belova OA, Bugmyrin SV, Kholodilov IS, Romanova LI, Karganova GG. Differentiation of Laboratory-Obtained Ixodes ricinus × Ixodes persulcatus Hybrid Ticks: Selection of Suitable Genes. Microorganisms. 2022; 10(7):1306. https://doi.org/10.3390/microorganisms10071306
Chicago/Turabian StyleLitov, Alexander G., Oxana A. Belova, Sergey V. Bugmyrin, Ivan S. Kholodilov, Lidia Iu. Romanova, and Galina G. Karganova. 2022. "Differentiation of Laboratory-Obtained Ixodes ricinus × Ixodes persulcatus Hybrid Ticks: Selection of Suitable Genes" Microorganisms 10, no. 7: 1306. https://doi.org/10.3390/microorganisms10071306
APA StyleLitov, A. G., Belova, O. A., Bugmyrin, S. V., Kholodilov, I. S., Romanova, L. I., & Karganova, G. G. (2022). Differentiation of Laboratory-Obtained Ixodes ricinus × Ixodes persulcatus Hybrid Ticks: Selection of Suitable Genes. Microorganisms, 10(7), 1306. https://doi.org/10.3390/microorganisms10071306