
Identification and Characterization of Rhipicephalus microplus ATAQ Homolog from Haemaphysalis longicornis Ticks and Its Immunogenic Potential as an Anti-Tick Vaccine Candidate Molecule

 ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Ticks and Rabbits
2.1.1. Ticks and Tissues Collection
2.1.2. Rabbits
2.1.3. Ethical Statement
2.2. Identification of HlATAQ cDNA
2.3. Gene Sequence and Phylogenetic Analyses of HlATAQ
2.4. Analysis of HlATAQ Expression by Real-Time PCR
2.5. RNA Interference
2.6. Localization of HlATAQ Protein by Immunohistochemistry
2.7. Expression and Purification of Recombinant HlATAQ
2.8. Vaccine Experiments
2.9. Statistical Analysis
3. Results
3.1. Identification of HlATAQ
3.2. Characterization of HlATAQ Amino Acid Sequence
3.3. BLAST and Phylogenetic Analyses of HlATAQ Amino Acid Sequence
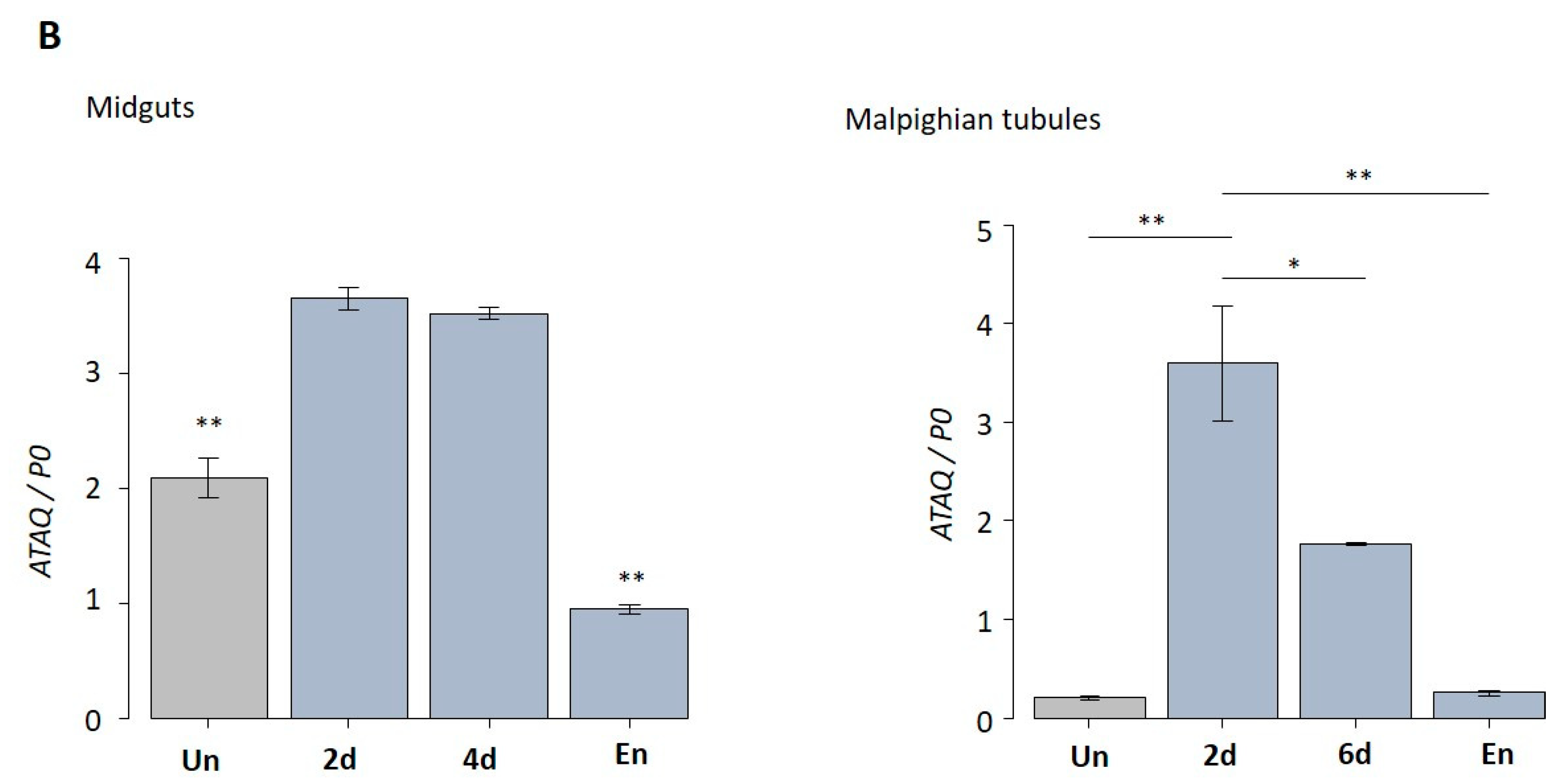
3.4. Expression Patterns of HlATAQ in Developmental Stages, Feeding Phases, and Tissues
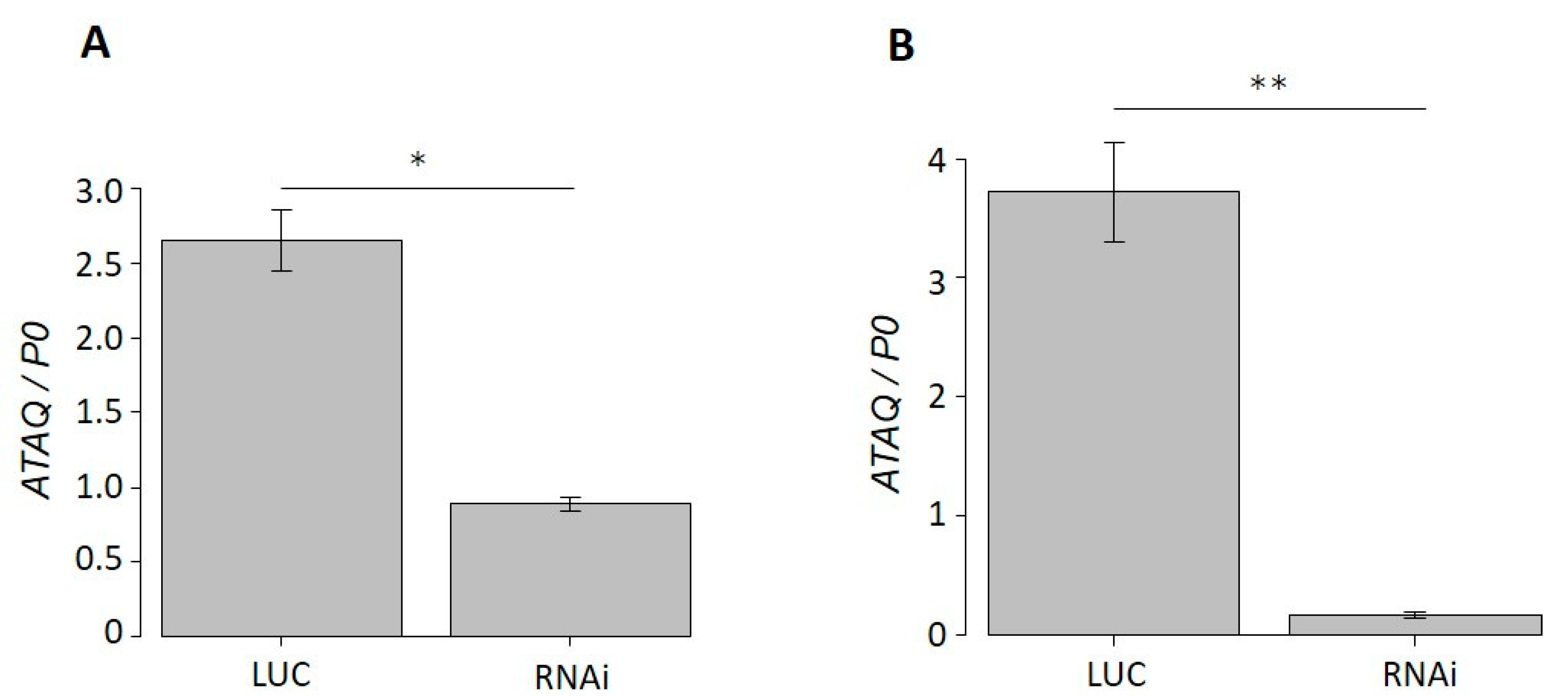
3.5. Phenotype of HlATAQ Gene-Knockdown Ticks
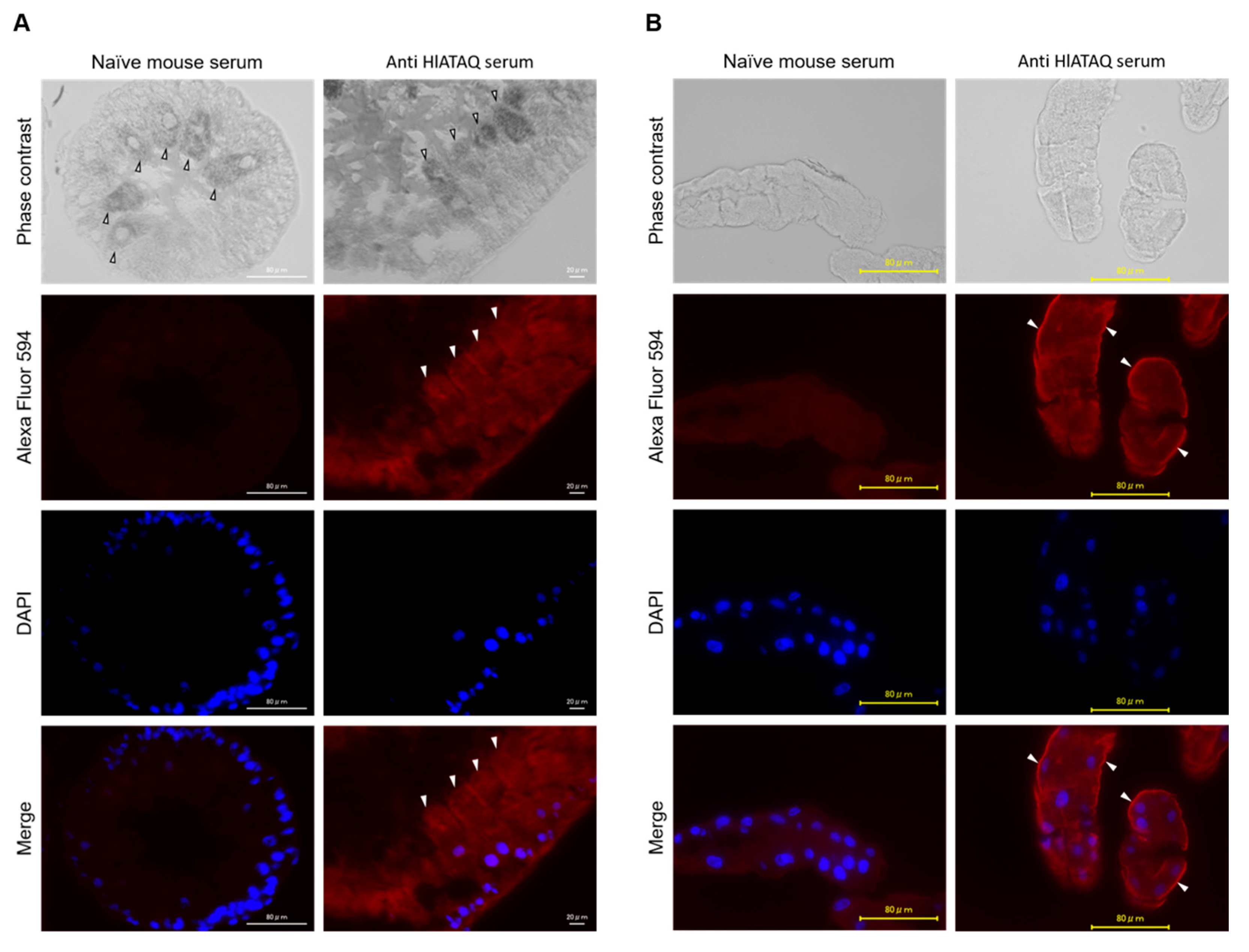
3.6. Detection of HlATAQ Protein in Tick Tissues
3.7. Expression, Purification, and Verification of Recombinant Proteins
3.8. Immune Response of Rabbit to Vaccination with Recombinant Proteins
3.9. Effect of Vaccination on H. longicornis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heath, A.C.G. Biology, ecology and distribution of the tick, Haemaphysalis longicornis Neumann (Acari: Ixodidae) in New Zealand. New Zealand Vet. J. 2016, 64, 10–20. [Google Scholar] [CrossRef]
- Hoogstraal, H.; Roberts, F.H.; Kohls, G.M.; Tipton, V.J. Review of Haemaphysalis (Kaiseriana) longicornis Neumann (resurrected) of Australia, New Zealand, New Caledonia, Fiji, Japan, Korea, and Northeastern China and USSR, and its parthenogenetic and bisexual populations (Ixodoidea, Ixodidae). J. Parasitol. 1968, 54, 1197–1213. [Google Scholar] [CrossRef] [PubMed]
- Egizi, A.; Bulaga-Seraphin, L.; Alt, E.; Bajwa, W.I.; Bernick, J.; Bickerton, M.; Campbell, S.R.; Connally, N.; Doi, K.; Falco, R.C.; et al. First glimpse into the origin and spread of the Asian longhorned tick, Haemaphysalis longicornis, in the United States. Zoonoses Public Health 2020, 67, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, T.; Okura, N.; Kakuda, H.; Yano, Y. Reproduction in a Metastriata tick, Haemaphysalis longicornis (Acari: Ixodidae). J. Acarol. Soc. Japan 2013, 22, 1–23. [Google Scholar] [CrossRef]
- Schappach, B.L.; Krell, R.K.; Hornbostel, V.L.; Connally, N.P. Exotic Haemaphysalis longicornis (Acari: Ixodidae) in the United States: Biology, Ecology, and Strategies for Management. J. Integr. Pest Manag. 2021, 11, 21. [Google Scholar] [CrossRef]
- Heath, A. Vector competence of Haemaphysalis longicornis with particular reference to blood parasites. Surveillance 2002, 29, 12–14. [Google Scholar]
- USDA. Monitoring Haemaphysalis longicornis, the Asian longhorned tick, populations in the United States; USDA: Washington, DC, USA, 2019; p. 14.
- Bickerton, M.; McSorley, K.; Alvaro, T. A life stage-targeted acaricide application approach for the control of Haemaphysalis longicornis. Ticks Tick. Borne. Dis. 2021, 12, 101581. [Google Scholar] [CrossRef]
- Lee, D.W.; Chang, K.S.; Kim, M.J.; Ahn, Y.J.; Jo, H.C.; Kim, S.-I.L. Acaricidal activity of commercialized insecticides against Haemaphysalis longicornis (Acari: Ixodidae) nymphs. J. Asia. Pac. Entomol. 2015, 18, 715–718. [Google Scholar] [CrossRef]
- Park, G.H.; Kim, H.K.; Lee, W.G.; Cho, S.H.; Kim, G.H. Evaluation of the acaricidal activity of 63 commercialized pesticides against Haemaphysalis longicornis (Acari: Ixodidae). Entomol. Res. 2019, 49, 330–336. [Google Scholar] [CrossRef]
- Heath, A.C.G.; Tenquist, J.D.; Bishop, D.M. Effects of pour-on organophosphate insecticides and a diamidide acaricide on the cattle tick, Haemaphysalis longicornis. New Zeal. J. Exp. Agric. 1980, 8, 79–87. [Google Scholar] [CrossRef]
- de la Fuente, J.; Contreras, M.; Estrada-Peña, A.; Cabezas-Cruz, A. Targeting a global health problem: Vaccine design and challenges for the control of tick-borne diseases. Vaccine 2017, 35, 5089–5094. [Google Scholar] [CrossRef]
- You, M.J. Resistance and control of cypermethrin and chlorpyrifos as acaricide for control of hard tick Haemaphysalis longicornis (Acari: Ixodidae). Korean J. Vet. Res. 2014, 54, 117–120. [Google Scholar] [CrossRef]
- de la Fuente, J.; Almazán, C.; Canales, M.; Pérez de la Lastra, J.M.; Kocan, K.M.; Willadsen, P. A ten-year review of commercial vaccine performance for control of tick infestations on cattle. Anim. Health Res. Rev. 2007, 8, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Valle, M.R.; Guerrero, F.D. Anti-tick vaccines in the omics era. Front. Biosci. 2018, 10, 122–136. [Google Scholar] [CrossRef] [PubMed]
- Nijhof, A.M.; Balk, J.A.; Postigo, M.; Marie, A.; Taoufik, A.; Jongejan, F. Bm86 homologues and novel ATAQ proteins with multiple epidermal growth factor (EGF)–like domains from hard and soft ticks. Int. J. Parasitol. 2010, 40, 1587–1597. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, F.D.; Miller, R.J.; Pérez de León, A.A. Cattle tick vaccines: Many candidate antigens, but will a commercially viable product emerge? Int. J. Parasitol. 2012, 42, 421–427. [Google Scholar] [CrossRef]
- Schetters, T.; Bishop, R.; Crampton, M.; Kopáček, P.; Lew-Tabor, A.; Maritz-Olivier, C.; Miller, R.; Mosqueda, J.; Patarroyo, J.; Rodriguez-Valle, M.; et al. Cattle tick vaccine researchers join forces in CATVAC. Parasites Vectors 2016, 9, 1–7. [Google Scholar] [CrossRef]
- Aguirre, A.D.A.R.; Lobo, F.P.; Cunha, R.C.; Garcia, M.V.; Andreotti, R. Design of the ATAQ peptide and its evaluation as an immunogen to develop a Rhipicephalus vaccine. Vet. Parasitol. 2016, 221, 30–38. [Google Scholar] [CrossRef]
- Lugo-Caro del Castillo, S.M.; Hernández-Ortiz, R.; Gómez-Romero, N.; Martínez-Velázquez, M.; Castro-Saines, E.; Lagunes-Quintanilla, R. Genetic diversity of the ATAQ gene in Rhipicephalus microplus collected in Mexico and implications as anti-tick vaccine. Parasitol. Res. 2020, 119, 3523–3529. [Google Scholar] [CrossRef]
- Fujisaki, K. Development of acquired resistance precipitating antibody in rabbits experimentally infested with females of Haemaphysalis longicornis (Ixodoidea: Ixodidae). Natl. Inst. Anim. Health Q. 1978, 18, 27–38. [Google Scholar]
- Kato, S.; Ohtoko, K.; Ohtake, H.; Kimura, T. Vector-capping: A simple method for preparing a high-quality full-length cDNA library. DNA Res. 2005, 12, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Liao, M.; Zhou, J.; Hatta, T.; Umemiya, R.; Miyoshi, T. Molecular characterization of Rhipicephalus (Boophilus) microplus Bm86 homologue from Haemaphysalis longicornis ticks. Vet. Parasitol. 2007, 146, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Umemiya-Shirafuji, R.; Zhou, J.; Liao, M.; Battsetseg, B.; Boldbaatar, D.; Hatta, T.; Kuboki, T.; Sakaguchi, T.; Chee, H.S.; Miyoshi, T.; et al. Data from expressed sequence tags from the organs and embryos of parthenogenetic Haemaphysalis longicornis. BMC Res. Notes 2021, 14, 14–17. [Google Scholar] [CrossRef]
- Sela, I.; Ashkenazy, H.; Katoh, K.; Pupko, T. GUIDANCE2: Accurate detection of unreliable alignment regions accounting for the uncertainty of multiple parameters. Nucleic Acids Res. 2015, 43, W7–W14. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Kuniyori, M.; Sato, N.; Yokoyama, N.; Kawazu, S.I.; Xuan, X.; Suzuki, H.; Fujisaki, K.; Umemiya-Shirafuji, R. Vitellogenin-2 accumulation in the fat body and hemolymph of Babesia-infected Haemaphysalis longicornis ticks. Front. Cell. Infect. Microbiol. 2022, 12, 908142. [Google Scholar] [CrossRef] [PubMed]
- Hatta, T.; Tsuji, N.; Miyoshi, T.; Islam, M.K.; Alim, M.A.; Yamaji, K.; Fujisaki, K. Leucine aminopeptidase, HlLAP, from the ixodid tick Haemaphysalis longicornis, plays vital roles in the development of oocytes. Parasitol. Int. 2010, 59, 286–289. [Google Scholar] [CrossRef]
- Miyoshi, T.; Tsuji, N.; Islam, M.K.; Kamio, T.; Fujisaki, K. Gene silencing of a cubilin-related serine proteinase from the hard tick Haemaphysalis longicornis by RNA interference. J. Vet. Med. Sci. 2004, 66, 1471–1473. [Google Scholar] [CrossRef]
- Kamio, T.; Fujisaki, K.; Minami, T. The improvement of “Ear bag” method for tick infestation. In Proceedings of the Japanese Association for Acarology; National Institute of Animal Health, Ed.: Tsukuba, Japan, 1987; pp. 1–4. [Google Scholar]
- Rand, K.N.; Moore, T.; Sriskantha, A.; Spring, K.; Tellam, R.; Willadsen, P.; Cobon, G.S. Cloning and expression of a protective antigen from the cattle tick Boophilus microplus. Proc. Natl. Acad. Sci. USA 1989, 86, 9657–9661. [Google Scholar] [CrossRef]
- Fass, D.; Blacklow, S.; Kim, P.S.; Berger, J.M. Molecular basis of familial hypercholesterolaemia from structure of LDL receptor module. Nature 1997, 388, 691–693. [Google Scholar] [CrossRef]
- Beffert, U.; Stolt, P.C.; Herz, J. Functions of lipoprotein receptors in neurons. J. Lipid Res. 2004, 45, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Almazan, C.; Tipacamu, G.A.; Rodriguez, S.; Mosqueda, J.; Perez de Leon, A. Immunological control of ticks and tick-borne diseases that impact cattle health and production. Front. Biosci. 2018, 23, 1535–1551. [Google Scholar] [CrossRef] [PubMed]
- Hulpiau, P.; van Roy, F. Molecular evolution of the cadherin superfamily. Int. J. Biochem. Cell Biol. 2009, 41, 349–369. [Google Scholar] [CrossRef] [PubMed]
- Naghibalhossaini, F.; Stanners, C. Minimal mutations are required to effect a radical change in function in CEA family members of the lg superfamily. J. Cell Sci. 2004, 117, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.A.; Rose, J.K. Sorting of GPI-anchored proteins to glycolipid-enriched membrane subdomains during transport to the apical cell surface. Cell 1992, 68, 533–544. [Google Scholar] [CrossRef]
- Ilangumaran, S.; Briol, A.; Hoessli, D.C. Distinct interactions among GPI-anchored, transmembrane and membrane associated intracellular proteins, and sphingolipids in lymphocyte and endothelial cell plasma membranes. Biochim. Biophys. Acta Biomembr. 1997, 1328, 227–236. [Google Scholar] [CrossRef]
- Chatterjee, S.; Mayor, S. The GPI-anchor and protein sorting. Cell. Mol. Life Sci. 2001, 58, 1969–1987. [Google Scholar] [CrossRef]
- Ball, A.; Campbell, E.M.; Jacob, J.; Hoppler, S.; Bowman, A.S. Identification, functional characterization and expression patterns of a water-specific aquaporin in the brown dog tick, Rhipicephalus sanguineus. Insect Biochem. Mol. Biol. 2009, 39, 105–112. [Google Scholar] [CrossRef]
- Trentelman, J.J.A.; Teunissen, H.; Kleuskens, J.A.G.M.; Van De Crommert, J.; De La Fuente, J.; Hovius, J.W.R.; Schetters, T.P.M. A combination of antibodies against Bm86 and Subolesin inhibits engorgement of Rhipicephalus australis (formerly Rhipicephalus microplus) larvae in vitro. Parasites Vectors 2019, 12, 1–10. [Google Scholar] [CrossRef]
- Nijhof, A.M.; Balk, J.A.; Postigo, M.; Jongejan, F. Selection of reference genes for quantitative RT-PCR studies in Rhipicephalus (Boophilus) microplus and Rhipicephalus appendiculatus ticks and determination of the expression profile of Bm86. BMC Mol. Biol. 2009, 10, 112. [Google Scholar] [CrossRef]
- Sonenshine, D.E. Excretion and Water Balance: Hindgut, Malpighian Tubules, and Coxal Glands. In Biology of Ticks; Oxford University Press: Oxford, UK, 2014; Volume 1, pp. 206–219. [Google Scholar]
- Nijhof, A.M.; Taoufik, A.; de la Fuente, J.; Kocan, K.M.; de Vries, E.; Jongejan, F. Gene silencing of the tick protective antigens, Bm86, Bm91 and subolesin, in the one-host tick Boophilus microplus by RNA interference. Int. J. Parasitol. 2007, 37, 653–662. [Google Scholar] [CrossRef] [PubMed]
- Narasimhan, S.; Montgomery, R.R.; DePonte, K.; Tschudi, C.; Marcantonio, N.; Anderson, J.F.; Sauer, J.R.; Cappello, M.; Kantor, F.S.; Fikrig, E. Disruption of Ixodes scapularis anticoagulation by using RNA interference. Proc. Natl. Acad. Sci. USA 2004, 101, 1141–1146. [Google Scholar] [CrossRef] [PubMed]
- Wouters, M.A.; Rigoutsos, I.; Chu, C.K.; Feng, L.L.; Sparrow, D.B.; Dunwoodie, S.L. Evolution of distinct EGF domains with specific functions. Protein Sci. 2005, 14, 1091–1103. [Google Scholar] [CrossRef] [PubMed]
- Riddell, D.R.; Graham, A.; Owen, J.S. Apolipoprotein E inhibits platelet aggregation through the L-arginine: Nitric oxide pathway: Implications for vascular disease. J. Biol. Chem. 1997, 272, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Riddell, D.R.; Vinogradov, D.V.; Stannard, A.K.; Chadwick, N.; Owen, J.S. Identification and characterization of LRP8 (apoER2) in human blood platelets. J. Lipid Res. 1999, 40, 1925–1930. [Google Scholar] [CrossRef] [PubMed]
- Koči, J.; Bista, S.; Chirania, P.; Yang, X.; Kitsou, C.; Rana, V.S.; Yas, O.B.; Sonenshine, D.E.; Pal, U. Antibodies against EGF-like domains in Ixodes scapularis BM86 orthologs impact tick feeding and survival of Borrelia burgdorferi. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Kaewmongkol, S.; Kaewmongkol, G.; Inthong, N.; Lakkitjaroen, N.; Sirinarumitr, T.; Berry, C.M.; Jonsson, N.N.; Stich, R.W.; Jittapalapong, S. Variation among Bm86 sequences in Rhipicephalus (Boophilus) microplus ticks collected from cattle across Thailand. Exp. Appl. Acarol. 2015, 66, 247–256. [Google Scholar] [CrossRef]
- Popara, M.; Villar, M.; Mateos-Hernández, L.; de Mera, I.G.F.; Marina, A.; del Valle, M.; Almazán, C.; Domingos, A.; de la Fuente, J. Lesser protein degradation machinery correlates with higher BM86 tick vaccine efficacy in Rhipicephalus annulatus when compared to Rhipicephalus microplus. Vaccine 2013, 31, 4728–4735. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Primer Name | Nucleotide Sequence (5′-3′) | At the Position of the Sequence a |
|---|---|---|---|
| Sequencing | |||
| pGCAP 1–2 | ACTGCTCCTCAGTGGATGTT | ||
| pGCAP-F | CTCAGTGGATGTTGCCTTTAC | ||
| M13 (-21) | TGTAAAACGACGGCCAGT | ||
| HlATAQ-1 | GAGGAATGTGCGAATGGTGG | 343–362 | |
| HlATAQ-2 | GAGGTTGAAGGTGTCTGCTCA | 856–876 | |
| HlATAQ-3 | ACAAACGAAACCGCAGGAGA | 1483–1502 | |
| T3 primer | ATTAACCCTCACTAAAGGGA | ||
| pGCAP-R | GCATTCTAGTTGTGGTTTGTCC | ||
| Real-time PCR | |||
| HlP0 F | CTCCATTGTCAACGGTCTCA | ||
| HlP0 R | TCAGCCTCCTTGAAGGTGAT | ||
| HlqPCR ATAQ-1 F | TCGGAAGATCCAATGCAACCAG | 1738–1759 | |
| HlqPCR ATAQ-1 R | TTAGGAGCACGACGACCACAAC | 1861–1882 | |
| dsRNA synthesis | |||
| HlATAQ-4 dsRNA | T7-HlATAQ-4 F | TAATACGACTCACTATAGGTGTGACTGTGGAGTGAACCG | 157–176 |
| T7-HlATAQ-4R | TAATACGACTCACTATAGGGAGCTCTTGATCTCCTCCGC | 704–723 | |
| HlATAQ-2 dsRNA | T7-HlATAQ-2 F | TAATACGACTCACTATAGGGAGGTTGAAGGTGTCTGCTCA | 856–876 |
| T7-HlATAQ-2 R | TAATACGACTCACTATAGGCGCCGTCTTGATGGCATAGA | 1410–1429 | |
| Recombinant protein expression | |||
| rHlATAQ | rHlATAQF | CGCGGATCCATGCCCTTCGAGATCGTG | |
| rHlATAQR | GGTGCTCGAGTCAGCATTCTTCCTCAGCG | ||
| T7 promoter | TAATACGACTCACTATAGGG | ||
| rHlATAQ middle | CGCCAAGGATGACCTTTGCAC | ||
| T7 terminator | ATGCTAGTTATTGCTCAGCGG | ||
| rtHlATAQ | rtHlATAQF | ACCCTCGAGATTTGCACAGCAGTTGGAC | |
| rtHlATAQR | TTCGGATCCTCAAGTACACGTCAATCCGTCAGC | ||
| Protein | Tick Species | Accession No. | AA No. | MW | pI | Glycosylation (N-Linked/O-Linked) | EGF Domains (Full/ Partial) | Anchor |
|---|---|---|---|---|---|---|---|---|
| HlATAQ | H. longicornis | ON210133 | 654 | 70.6 | 4.6 | 8/12 | 6/1 | TM |
| BmATAQ | R. (B.) microplus | GU144589 | 605 | 66.6 | 4.82 | 8/2 | 6/1 | TM |
| BdATAQ | R. (B.) decoloratus | GU144591 | 605 | 66.5 | 5.16 | 8/4 | 6/1 | TM |
| BaATAQ | R. (B.) annulatus | GU144590 | 605 | 66.4 | 5.05 | 8/2 | 6/1 | TM |
| ReeATAQ | R. evertsi evertsi | GU144592 | 605 | 66.4 | 4.95 | 8/3 | 6/1 | TM |
| RaATAQ-1 | R. appendiculatus | GU144594 | 605 | 66.7 | 5.42 | 7/3 | 6/1 | TM |
| RaATAQ-2 | R. appendiculatus | GU144593 | 561 | 61.6 | 5.33 | 8/1 | 6/1 | TM |
| HmATAQ | Hy. m. marginatum | GU144595 | 601 | 65.5 | 5.18 | 5/1 | 6/1 | TM |
| DrATAQ | D. reticulatus | GU144596 | 596 | 64.7 | 4.79 | 9/2 | 6/1 | TM |
| DvATAQ | D. variabilis | GU144597 | 598 | 65.0 | 4.84 | 6/4 | 6/1 | TM |
| HeATAQ | H. elliptica | GU144598 | 597 | 65.6 | 5.47 | 3/17 | 6/1 | GPI |
| AvATAQ | A. variegatum | GU144599 | 522 | 57.5 | 5.04 | 4/1 | 6/1 | GPI |
| Hl86 | H. longicornis | AB251858 | 594 | 66.6 | 5.6 | 5/10 | 6/1 | GPI |
| Bm86 | R. (B.) microplus | M29321 | 650 | 71.72 | 5.6 | 4/2 | 8/1 | GPI |
| ATAQ Sequences | Homology to HlATAQ (%) | ||
|---|---|---|---|
| Tick species | Country | Identity | Similarity |
| D. reticulatus | Netherlands | 48.61 | 53.43 |
| D. variabilis | USA | 47.59 | 51.38 |
| Hy. m. marginatum | France | 46.86 | 51.82 |
| R. (B.) decoloratus | South Africa | 44.23 | 49.34 |
| R. sanguineus | China | 44.08 | 49.63 |
| R. (B.) microplus | Mozambique | 43.94 | 49.34 |
| R. (B.) microplus | Mexico (Media Joya) | 43.94 | 49.19 |
| R. (B.) microplus | Mexico (Moyahua) | 43.94 | 49.34 |
| R. (B.) microplus | Mexico (Campeche) | 43.94 | 49.19 |
| R. (B.) microplus | Mexico (Hybrid) | 43.94 | 49.34 |
| R. (B.) microplus | Mexico (Verdineno) | 43.94 | 49.34 |
| R. evertsi evertsi | South Africa | 43.79 | 49.05 |
| R. (B.) microplus | Mexico (Vargas) | 43.64 | 49.05 |
| R. (B.) microplus | Mexico (Huastecas) | 43.50 | 49.05 |
| R. (B.) annulatus | Israel | 43.35 | 48.46 |
| R. appendiculatus | South Africa | 43.35 | 49.63 |
| R. appendiculatus | South Africa | 40.72 | 46.27 |
| D. silvarum | China | 40.72 | 42.91 |
| A. variegatum | Gambia | 34.16 | 38.68 |
| H. elliptica | South Africa | 32.70 | 37.51 |
| Groups | No. of Ticks | Blood Feeding Period (Days) | Body Weight of Engorged Ticks (mg) | Egg Mass/Body Weight (%) | Pre- Oviposition Period (Days) | Egg Hatching Period (Days) |
|---|---|---|---|---|---|---|
| Control-1 | 22 | 7.2 ± 0.2 | 183.4 ± 8.9 | 41.2 ± 1.8 | 6.1 ± 0.2 | 38.9 ± 0.5 |
| RNAi/HlATAQ-2 dsRNA | 19 | 6.9 ± 0.2 | 182.8 ± 10.2 | 45.0±2.0 | 5.9 ± 0.2 | 39.4 ± 0.5 |
| Control-2 | 22 | 7.2 ± 0.2 | 231.8 ± 8.8 | 56.1 ± 0.8 | 5.9 ± 0.2 | 36.4 ± 0.3 |
| RNAi/HlATAQ-4 dsRNA | 22 | 7.5 ± 0.2 | 232.5 ± 7.5 | 54.7 ± 0.6 | 5.8 ± 0.2 | 36.5 ± 0.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adjou Moumouni, P.F.; Naomasa, S.; Tuvshintulga, B.; Sato, N.; Okado, K.; Zheng, W.; Lee, S.-H.; Mosqueda, J.; Suzuki, H.; Xuan, X.; et al. Identification and Characterization of Rhipicephalus microplus ATAQ Homolog from Haemaphysalis longicornis Ticks and Its Immunogenic Potential as an Anti-Tick Vaccine Candidate Molecule. Microorganisms 2023, 11, 822. https://doi.org/10.3390/microorganisms11040822
Adjou Moumouni PF, Naomasa S, Tuvshintulga B, Sato N, Okado K, Zheng W, Lee S-H, Mosqueda J, Suzuki H, Xuan X, et al. Identification and Characterization of Rhipicephalus microplus ATAQ Homolog from Haemaphysalis longicornis Ticks and Its Immunogenic Potential as an Anti-Tick Vaccine Candidate Molecule. Microorganisms. 2023; 11(4):822. https://doi.org/10.3390/microorganisms11040822
Chicago/Turabian StyleAdjou Moumouni, Paul Franck, Souichirou Naomasa, Bumduuren Tuvshintulga, Nariko Sato, Kiyoshi Okado, Weiqing Zheng, Seung-Hun Lee, Juan Mosqueda, Hiroshi Suzuki, Xuenan Xuan, and et al. 2023. "Identification and Characterization of Rhipicephalus microplus ATAQ Homolog from Haemaphysalis longicornis Ticks and Its Immunogenic Potential as an Anti-Tick Vaccine Candidate Molecule" Microorganisms 11, no. 4: 822. https://doi.org/10.3390/microorganisms11040822
APA StyleAdjou Moumouni, P. F., Naomasa, S., Tuvshintulga, B., Sato, N., Okado, K., Zheng, W., Lee, S.-H., Mosqueda, J., Suzuki, H., Xuan, X., & Umemiya-Shirafuji, R. (2023). Identification and Characterization of Rhipicephalus microplus ATAQ Homolog from Haemaphysalis longicornis Ticks and Its Immunogenic Potential as an Anti-Tick Vaccine Candidate Molecule. Microorganisms, 11(4), 822. https://doi.org/10.3390/microorganisms11040822