
Insights on Gut and Skin Wound Microbiome in Stranded Indo-Pacific Finless Porpoise (Neophocaena phocaenoides)

 , ,
, ,  , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Genomic DNA Extraction
2.3. Library Construction and Sequencing
2.4. Sequencing Data Processing
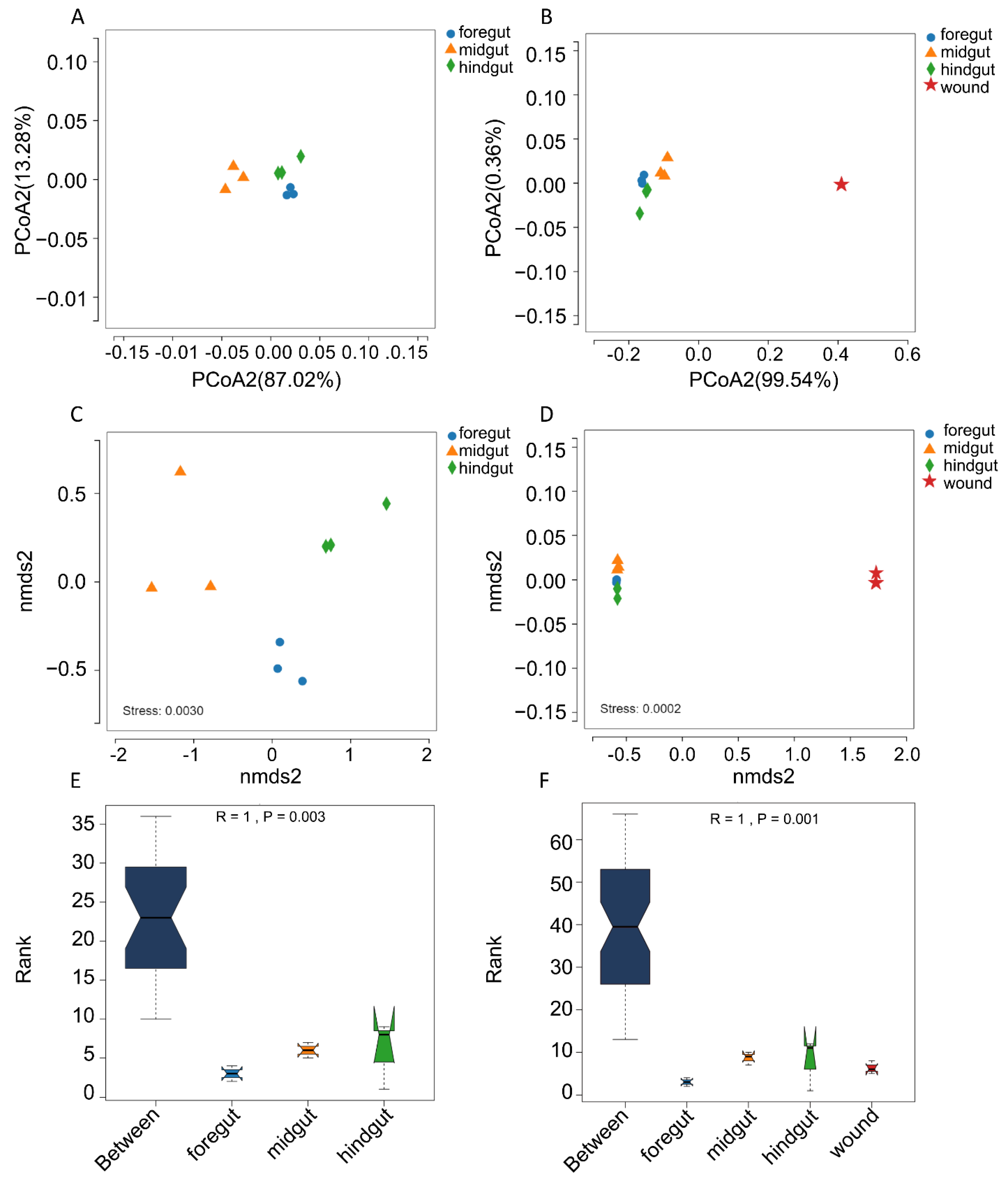
2.5. Microbiome Diversity and Structure Comparison
2.6. Statistical Analyses
3. Results
3.1. Subsection
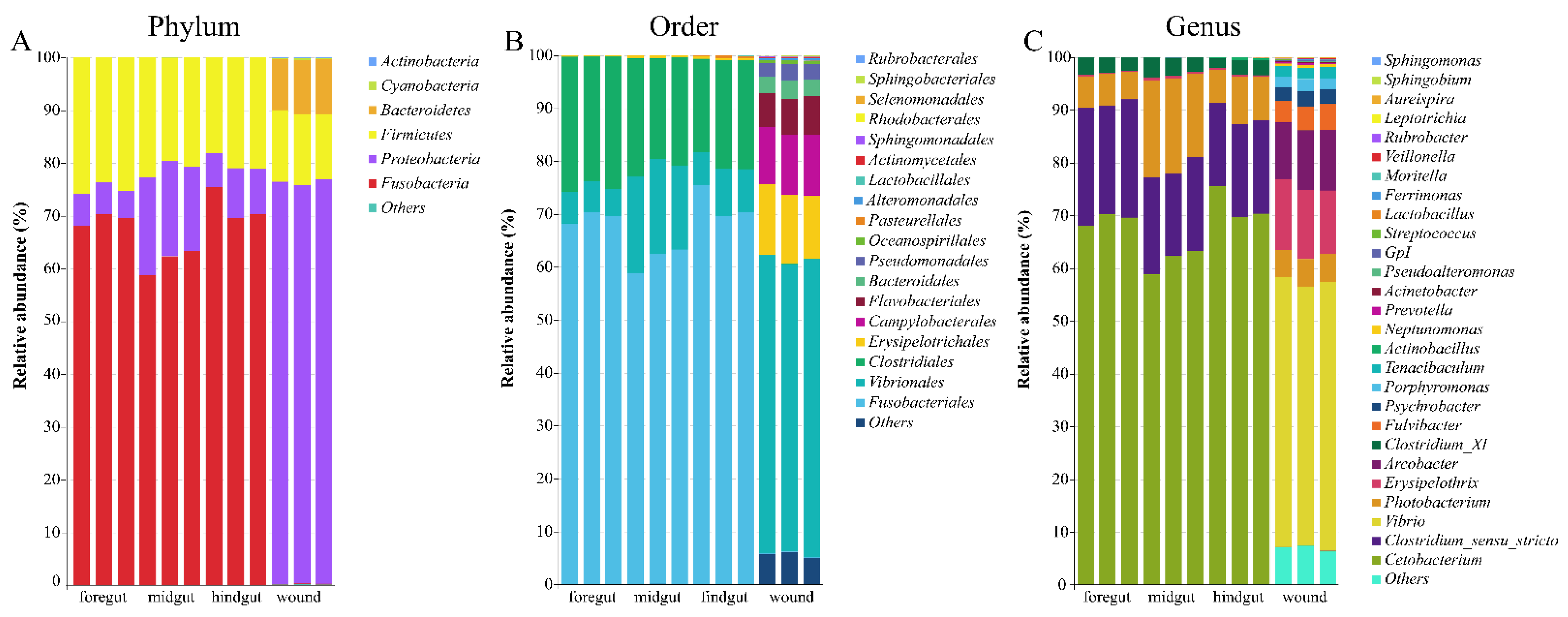
3.2. Microbial Taxonomic Profiles
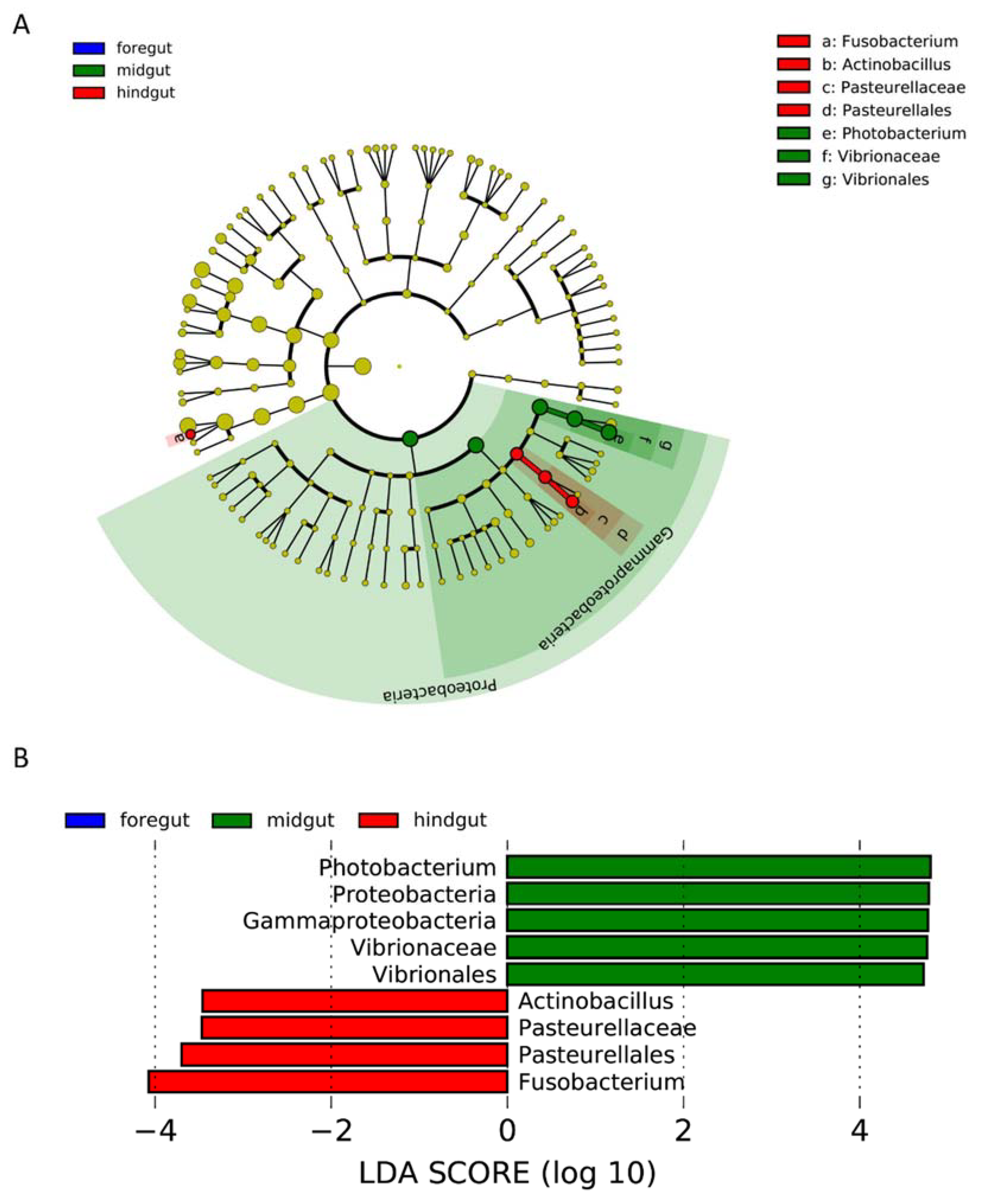
3.3. Identification of Key Microbes in Each Anatomical Region
4. Discussion
4.1. Microbiome of Gastrointestinal Tract of Indo-Pacific Finless Porpoise
4.2. Microbiome on the Skin Wounds of the Indo-Pacific Finless Porpoise
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cho, I.; Blaser, M.J. The human microbiome: At the interface of health and disease. Nat. Rev. Genet. 2012, 13, 260–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundin, O.H.; Mendoza-Ladd, A.; Zeng, M.; Diaz-Arevalo, D.; Morales, E.; Fagan, B.M.; Ordonez, J.; Velez, P.; Antony, N.; McCallum, R.W. The human jejunum has an endogenous microbiota that differs from those in the oral cavity and colon. BMC Microbiol. 2017, 17, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez-Repiso, C.; Moreno-Indias, I.; Martin-Nunez, G.M.; Ho-Plagaro, A.; Rodriguez-Canete, A.; Gonzalo, M.; Garcia-Fuentes, E.; Tinahones, F.J. Mucosa-associated microbiota in the jejunum of patients with morbid obesity: Alterations in states of insulin resistance and metformin treatment. Surg. Obes. Related Dis. Off. J. Am. Soc. Bariat. Surg. 2020, 16, 1575–1585. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Zhang, P.; Zhang, C.; Du, J.; Du, X.; Zhu, C.; Liu, J.; Xie, P.; Li, S. Comparative Study of the Gut Microbiota Among Four Different Marine Mammals in an Aquarium. Front. Microbiol. 2021, 12, 769012. [Google Scholar] [CrossRef]
- Bai, S.; Zhang, P.; Lin, M.; Lin, W.; Yang, Z.; Li, S. Microbial diversity and structure in the gastrointestinal tracts of two stranded short-finned pilot whales (Globicephala macrorhynchus) and a pygmy sperm whale (Kogia breviceps). Integr. Zool. 2021, 16, 324–335. [Google Scholar] [CrossRef]
- Henry, L.P.; Bruijning, M.; Forsberg, S.K.G.; Ayroles, J.F. The microbiome extends host evolutionary potential. Nat. Commun. 2021, 12, 5141. [Google Scholar] [CrossRef]
- Suzuki, A.; Segawa, T.; Sawa, S.; Nishitani, C.; Ueda, K.; Itou, T.; Asahina, K.; Suzuki, M. Comparison of the gut microbiota of captive common bottlenose dolphins Tursiops truncatus in three aquaria. J. Appl. Microbiol. 2019, 126, 31–39. [Google Scholar] [CrossRef]
- Salazar, N.; Valdes-Varela, L.; Gonzalez, S.; Gueimonde, M.; de Los Reyes-Gavilan, C.G. Nutrition and the gut microbiome in the elderly. Gut Microb. 2017, 8, 82–97. [Google Scholar] [CrossRef] [Green Version]
- Russo, C.D.; Weller, D.W.; Nelson, K.E.; Chivers, S.J.; Torralba, M.; Grimes, D.J. Bacterial Species Identified on the Skin of Bottlenose Dolphins Off Southern California via Next Generation Sequencing Techniques. Microb. Ecol. 2018, 75, 303–309. [Google Scholar] [CrossRef]
- Sanders, J.G.; Beichman, A.C.; Roman, J.; Scott, J.J.; Emerson, D.; McCarthy, J.J.; Girguis, P.R. Baleen whales host a unique gut microbiome with similarities to both carnivores and herbivores. Nat. Commun. 2015, 6, 8285. [Google Scholar] [CrossRef]
- Wan, X.; Li, J.; Cheng, Z.; Ao, M.; Tian, R.; McLaughlin, R.W.; Zheng, J.; Wang, D. The intestinal microbiome of an Indo-Pacific humpback dolphin (Sousa chinensis) stranded near the Pearl River Estuary, China. Integr. Zool. 2021, 16, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.L.; McLaughlin, R.W.; Zheng, J.S.; Hao, Y.J.; Fan, F.; Tian, R.M.; Wang, D. Microbial communities in different regions of the gastrointestinal tract in East Asian finless porpoises (Neophocaena asiaeorientalis sunameri). Sci. Rep. 2018, 8, 14142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, A.; Akuzawa, K.; Kogi, K.; Ueda, K.; Suzuki, M. Captive environment influences the composition and diversity of fecal microbiota in Indo-Pacific bottlenose dolphins, Tursiops aduncus. Marine Mammal Sci. 2021, 37, 207–219. [Google Scholar] [CrossRef]
- Callewaert, C.; Ravard Helffer, K.; Lebaron, P. Skin Microbiome and its Interplay with the Environment. Am. J. Clin. Dermatol. 2020, 21, 4–11. [Google Scholar] [CrossRef]
- Zhuang, M.; Sanganyado, E.; Li, P.; Liu, W. Distribution of microbial communities in metal-contaminated nearshore sediment from Eastern Guangdong, China. Environ. Pollut. 2019, 250, 482–492. [Google Scholar] [CrossRef]
- Hong, H.; Qiu, J.; Liang, Y. Environmental factors influencing the distribution of total and fecal coliform bacteria in six water storage reservoirs in the Pearl River Delta Region, China. J. Environ. Sci. 2010, 22, 663–668. [Google Scholar] [CrossRef]
- Chen, L.; Tsui, M.M.P.; Lam, J.C.W.; Hu, C.; Wang, Q.; Zhou, B.; Lam, P.K.S. Variation in microbial community structure in surface seawater from Pearl River Delta: Discerning the influencing factors. Sci. Total Environ. 2019, 660, 136–144. [Google Scholar] [CrossRef]
- Bik, E.M.; Costello, E.K.; Switzer, A.D.; Callahan, B.J.; Holmes, S.P.; Wells, R.S.; Carlin, K.P.; Jensen, E.D.; Venn-Watson, S.; Relman, D.A. Marine mammals harbor unique microbiotas shaped by and yet distinct from the sea. Nat. Commun. 2016, 7, 10516. [Google Scholar] [CrossRef] [Green Version]
- Apprill, A.; Miller, C.A.; Van Cise, A.M.; U’Ren, J.M.; Leslie, M.S.; Weber, L.; Baird, R.W.; Robbins, J.; Landry, S.; Bogomolni, A.; et al. Marine mammal skin microbiotas are influenced by host phylogeny. R. Soc. Open. Sci. 2020, 7, 192046. [Google Scholar] [CrossRef]
- Chiarello, M.; Villeger, S.; Bouvier, C.; Auguet, J.C.; Bouvier, T. Captive bottlenose dolphins and killer whales harbor a species-specific skin microbiota that varies among individuals. Sci. Rep. 2017, 7, 15269. [Google Scholar] [CrossRef] [Green Version]
- Khazaei, T.; Williams, R.L.; Bogatyrev, S.R.; Doyle, J.C.; Henry, C.S.; Ismagilov, R.F. Metabolic multistability and hysteresis in a model aerobe-anaerobe microbiome community. Sci. Adv. 2020, 6, eaba0353. [Google Scholar] [CrossRef] [PubMed]
- Tomic-Canic, M.; Burgess, J.L.; O’Neill, K.E.; Strbo, N.; Pastar, I. Skin Microbiota and its Interplay with Wound Healing. Am. J. Clin. Dermatol. 2020, 21, 36–43. [Google Scholar] [CrossRef]
- Johnson, T.R.; Gomez, B.I.; McIntyre, M.K.; Dubick, M.A.; Christy, R.J.; Nicholson, S.E.; Burmeister, D.M. The Cutaneous Microbiome and Wounds: New Molecular Targets to Promote Wound Healing. Int. J. Mol. Sci. 2018, 19, 2699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Dunaway, S.; Champer, J.; Kim, J.; Alikhan, A. Changing our microbiome: Probiotics in dermatology. Br. J. Dermatol. 2020, 182, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Gardner, S.E.; Hillis, S.L.; Heilmann, K.; Segre, J.A.; Grice, E.A. The neuropathic diabetic foot ulcer microbiome is associated with clinical factors. Diabetes 2013, 62, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Price, L.B.; Liu, C.M.; Frankel, Y.M.; Melendez, J.H.; Aziz, M.; Buchhagen, J.; Contente-Cuomo, T.; Engelthaler, D.M.; Keim, P.S.; Ravel, J.; et al. Macroscale spatial variation in chronic wound microbiota: A cross-sectional study. Wound Rep. Regen. 2011, 19, 80–88. [Google Scholar] [CrossRef] [Green Version]
- Wolcott, R.D.; Hanson, J.D.; Rees, E.J.; Koenig, L.D.; Phillips, C.D.; Wolcott, R.A.; Cox, S.B.; White, J.S. Analysis of the chronic wound microbiota of 2,963 patients by 16S rDNA pyrosequencing. Wound Rep. Regen. 2016, 24, 163–174. [Google Scholar] [CrossRef]
- Bierlich, K.C.; Miller, C.; DeForce, E.; Friedlaender, A.S.; Johnston, D.W.; Apprill, A. Temporal and Regional Variability in the Skin Microbiome of Humpback Whales along the Western Antarctic Peninsula. Appl. Environ. Microbiol. 2018, 84, e02574-02517. [Google Scholar] [CrossRef] [Green Version]
- Apprill, A.; Robbins, J.; Eren, A.M.; Pack, A.A.; Reveillaud, J.; Mattila, D.; Moore, M.; Niemeyer, M.; Moore, K.M.; Mincer, T.J. Humpback whale populations share a core skin bacterial community: Towards a health index for marine mammals? PLoS ONE 2014, 9, e90785. [Google Scholar] [CrossRef]
- Jefferson, T.A.; Webber, M.A.; Pitman, R.L. Marine Mammals of the World a Comprehensive Guide to Their Identification; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Naik, S.; Bouladoux, N.; Linehan, J.L.; Han, S.J.; Harrison, O.J.; Wilhelm, C.; Conlan, S.; Himmelfarb, S.; Byrd, A.L.; Deming, C.; et al. Commensal-dendritic-cell interaction specifies a unique protective skin immune signature. Nature 2015, 520, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Linehan, J.L.; Harrison, O.J.; Han, S.J.; Byrd, A.L.; Vujkovic-Cvijin, I.; Villarino, A.V.; Sen, S.K.; Shaik, J.; Smelkinson, M.; Tamoutounour, S.; et al. Non-classical Immunity Controls Microbiota Impact on Skin Immunity and Tissue Repair. Cell 2018, 172, 784–796.e718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christaki, E.; Giamarellos-Bourboulis, E.J. The complex pathogenesis of bacteremia: From antimicrobial clearance mechanisms to the genetic background of the host. Virulence 2014, 5, 57–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Cise, A.M.; Wade, P.R.; Goertz, C.E.C.; Burek-Huntington, K.; Parsons, K.M.; Clauss, T.; Hobbs, R.C.; Apprill, A. Skin microbiome of beluga whales: Spatial, temporal, and health-related dynamics. Anim. Microb. 2020, 2, 39. [Google Scholar] [CrossRef] [PubMed]
- Siebert, U.; Wünschmann, A.; Weiss, R.; Frank, H.; Benke, H.; Frese, K. Post-mortem Findings in Harbour Porpoises (Phocoena phocoena) from the German North and Baltic Seas. J. Comp. Pathol. 2001, 124, 102–114. [Google Scholar] [CrossRef]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; Du, J.; Lu, Z.; Han, J.; Wang, Z.; Li, D.; Guan, X.; Wang, Z. Distribution of microbiota across different intestinal tract segments of a stranded dwarf minke whale, Balaenoptera acutorostrata. Microbiologyopen 2020, 9, e1108. [Google Scholar] [CrossRef] [PubMed]
- You, L.; Ying, C.; Liu, K.; Zhang, X.; Lin, D.; Yin, D.; Zhang, J.; Xu, P. Changes in the fecal microbiome of the Yangtze finless porpoise during a short-term therapeutic treatment. Open. Life Sci. 2020, 15, 296–310. [Google Scholar] [CrossRef]
- Cockburn, D.W.; Koropatkin, N.M. Polysaccharide Degradation by the Intestinal Microbiota and Its Influence on Human Health and Disease. J. Mol. Biol. 2016, 428, 3230–3252. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Wang, X.; Yang, S.; Meng, F.; Wang, X.; Wei, H.; Chen, T. Evaluation of the Microbial Diversity in Amyotrophic Lateral Sclerosis Using High-Throughput Sequencing. Front. Microbiol. 2016, 7, 1479. [Google Scholar] [CrossRef] [Green Version]
- Labella, A.M.; Arahal, D.R.; Castro, D.; Lemos, M.L.; Borrego, J.J. Revisiting the genus Photobacterium: Taxonomy, ecology and pathogenesis. Int. Microbiol. Off. J. Span. Soc. Microbiol. 2017, 20, 1–10. [Google Scholar] [CrossRef]
- Labella, A.M.; Castro, M.D.; Manchado, M.; Lucena, T.; Arahal, D.R.; Borrego, J.J. Photobacterium malacitanum sp. nov., and Photobacterium andalusiense sp. nov., two new bacteria isolated from diseased farmed fish in Southern Spain. Syst. Appl. Microbiol. 2018, 41, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Altekruse, S.F.; Bishop, R.D.; Baldy, L.M.; Thompson, S.G.; Wilson, S.A.; Ray, B.J.; Griffin, P.M. Vibrio gastroenteritis in the US Gulf of Mexico region: The role of raw oysters. Epidemiol. Infect. 2000, 124, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Godoy-Vitorino, F.; Rodriguez-Hilario, A.; Alves, A.L.; Gonçalves, F.; Cabrera-Colon, B.; Mesquita, C.S.; Soares-Castro, P.; Ferreira, M.; Marçalo, A.; Vingada, J.; et al. The microbiome of a striped dolphin (Stenella coeruleoalba) stranded in Portugal. Res. Microbiol. 2017, 168, 85–93. [Google Scholar] [CrossRef]
- Zhang, X.; Sun, Z.; Zhang, X.; Zhang, M.; Li, S. Hemolymph Microbiomes of Three Aquatic Invertebrates as Revealed by a New Cell Extraction Method. Appl. Environ. Microbiol. 2018, 84, e02824-17. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.J.; Rangdale, R.E. 22-Tracing pathogens in molluscan shellfish production chains. In Tracing Pathogens in the Food Chain; Brul, S., Fratamico, P.M., McMeekin, T.A., Eds.; Woodhead Publishing: Sawston, UK, 2011; pp. 527–547. [Google Scholar] [CrossRef]
- Acevedo-Whitehouse, K.; Rocha-Gosselin, A.; Gendron, D. A novel non-invasive tool for disease surveillance of free-ranging whales and its relevance to conservation programs. Anim. Conserv. 2010, 13, 217–225. [Google Scholar] [CrossRef]
- Hansen, M.J.; Bertelsen, M.F.; Christensen, H.; Bisgaard, M.; Bojesen, A.M. Occurrence of Pasteurellaceae bacteria in the oral cavity of selected marine mammal species. J. Zoo Wild. Med. 2012, 43, 828–835. [Google Scholar] [CrossRef] [PubMed]
- Tobias, S.; Lee, J.H.; Tomford, J.W. Rare Actinobacillus infection of the cavernous sinus causing painful ophthalmoplegia: Case report. Neurosurgery 2002, 51, 807–809, discussion 809–810. [Google Scholar] [CrossRef] [PubMed]
- Liven, E.; Larsen, H.J.; Lium, B. Infection with actinobacillus suis in pigs. Acta Veter. Scand. 1978, 19, 313–315. [Google Scholar] [CrossRef]
- Beineke, A.; Siebert, U.; Baumgartner, W. The immune system of marine mammals. Part 1: Immune response, cytokines and immunotoxicity. Tieraerztliche Prax. Ausg. Kleintiere Heimtiere 2006, 34, 118–126. [Google Scholar]
- Apprill, A.; Mooney, T.A.; Lyman, E.; Stimpert, A.K.; Rappe, M.S. Humpback whales harbour a combination of specific and variable skin bacteria. Environ. Microbiol. Rep. 2011, 3, 223–232. [Google Scholar] [CrossRef]
- Grice, E.A.; Kong, H.H.; Conlan, S.; Deming, C.B.; Davis, J.; Young, A.C.; Program, N.C.S.; Bouffard, G.G.; Blakesley, R.W.; Murray, P.R.; et al. Topographical and temporal diversity of the human skin microbiome. Science 2009, 324, 1190–1192. [Google Scholar] [CrossRef] [Green Version]
- Wattrang, E.; Eriksson, H.; Jinnerot, T.; Persson, M.; Bagge, E.; Söderlund, R.; Naghizadeh, M.; Dalgaard, T.S. Immune responses upon experimental Erysipelothrix rhusiopathiae infection of naïve and vaccinated chickens. Veter. Res. 2020, 51, 114. [Google Scholar] [CrossRef]
- Liu, L.; Xiao, J.; Zhang, M.; Zhu, W.; Xia, X.; Dai, X.; Pan, Y.; Yan, S.; Wang, Y. A Vibrio owensii strain as the causative agent of AHPND in cultured shrimp, Litopenaeus vannamei. J. Invert. Pathol. 2018, 153, 156–164. [Google Scholar] [CrossRef]
- Goulden, E.F.; Hall, M.R.; Bourne, D.G.; Pereg, L.L.; Høj, L. Pathogenicity and infection cycle of Vibrio owensii in larviculture of the ornate spiny lobster (Panulirus ornatus). Appl. Environ. Microbiol. 2012, 78, 2841–2849. [Google Scholar] [CrossRef] [Green Version]
- Reboli, A.C.; Farrar, W.E. Erysipelothrix rhusiopathiae: An occupational pathogen. Clin. Microbiol. Rev. 1989, 2, 354–359. [Google Scholar] [CrossRef]
- Ceccolini, M.E.; Wessels, M.; Macgregor, S.K.; Deaville, R.; Perkins, M.; Jepson, P.D.; John, S.K.; Guthrie, A. Systemic Erysipelothrix rhusiopathiae in seven free-ranging delphinids stranded in England and Wales. Dis. Aquat. Organ. 2021, 145, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Pogoreutz, C.; Gore, M.A.; Perna, G.; Millar, C.; Nestler, R.; Ormond, R.F.; Clarke, C.R.; Voolstra, C.R. Similar bacterial communities on healthy and injured skin of black tip reef sharks. Anim. Microb. 2019, 1, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faílde, L.D.; Losada, A.P.; Bermúdez, R.; Santos, Y.; Quiroga, M.I. Evaluation of immune response in turbot (Psetta maxima L.) tenacibaculosis: Haematological and immunohistochemical studies. Microb. Pathog. 2014, 76, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hundenborn, J.; Thurig, S.; Kommerell, M.; Haag, H.; Nolte, O. Severe Wound Infection with Photobacterium damselae ssp. damselae and Vibrio harveyi, following a Laceration Injury in Marine Environment: A Case Report and Review of the Literature. Case Rep. Med. 2013, 2013, 610632. [Google Scholar] [CrossRef] [Green Version]
- DeBruyn, J.M.; Hauther, K.A. Postmortem succession of gut microbial communities in deceased human subjects. PeerJ 2017, 5, e3437. [Google Scholar] [CrossRef] [Green Version]
- Hauther, K.A.; Cobaugh, K.L.; Jantz, L.M.; Sparer, T.E.; DeBruyn, J.M. Estimating Time Since Death from Postmortem Human Gut Microbial Communities. J. Forensic Sci. 2015, 60, 1234–1240. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Xie, H.; Sun, Y.; Zeng, Y.; Tian, Z.; Chen, X.; Sanganyado, E.; Lin, J.; Yang, L.; Li, P.; et al. Insights on Gut and Skin Wound Microbiome in Stranded Indo-Pacific Finless Porpoise (Neophocaena phocaenoides). Microorganisms 2022, 10, 1295. https://doi.org/10.3390/microorganisms10071295
Li C, Xie H, Sun Y, Zeng Y, Tian Z, Chen X, Sanganyado E, Lin J, Yang L, Li P, et al. Insights on Gut and Skin Wound Microbiome in Stranded Indo-Pacific Finless Porpoise (Neophocaena phocaenoides). Microorganisms. 2022; 10(7):1295. https://doi.org/10.3390/microorganisms10071295
Chicago/Turabian StyleLi, Chengzhang, Huiying Xie, Yajing Sun, Ying Zeng, Ziyao Tian, Xiaohan Chen, Edmond Sanganyado, Jianqing Lin, Liangliang Yang, Ping Li, and et al. 2022. "Insights on Gut and Skin Wound Microbiome in Stranded Indo-Pacific Finless Porpoise (Neophocaena phocaenoides)" Microorganisms 10, no. 7: 1295. https://doi.org/10.3390/microorganisms10071295
APA StyleLi, C., Xie, H., Sun, Y., Zeng, Y., Tian, Z., Chen, X., Sanganyado, E., Lin, J., Yang, L., Li, P., Liang, B., & Liu, W. (2022). Insights on Gut and Skin Wound Microbiome in Stranded Indo-Pacific Finless Porpoise (Neophocaena phocaenoides). Microorganisms, 10(7), 1295. https://doi.org/10.3390/microorganisms10071295