
Variations in the Sporulation Efficiency of Pathogenic Freshwater Oomycetes in Relation to the Physico-Chemical Properties of Natural Waters

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
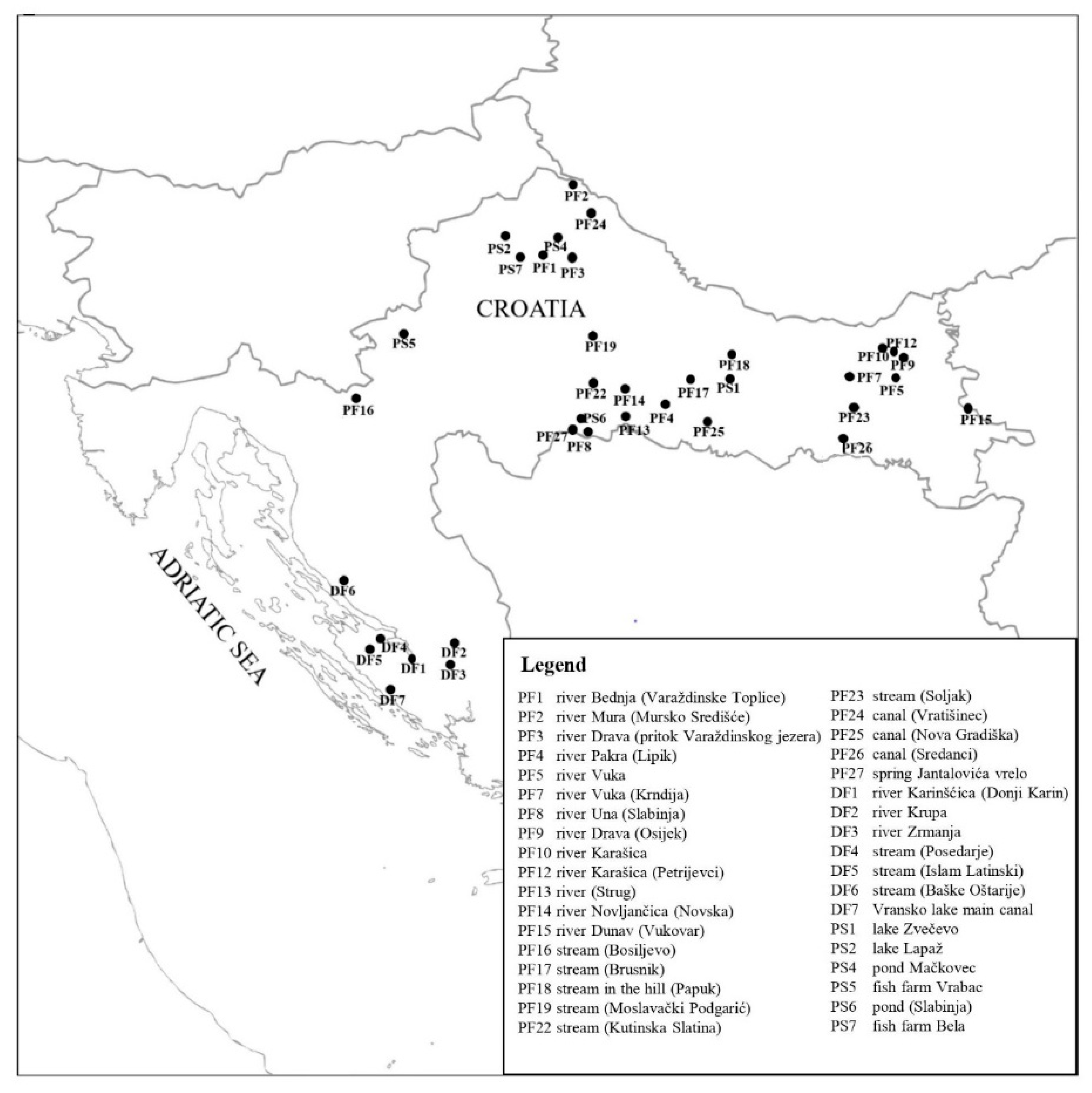
2.1. Water Sampling
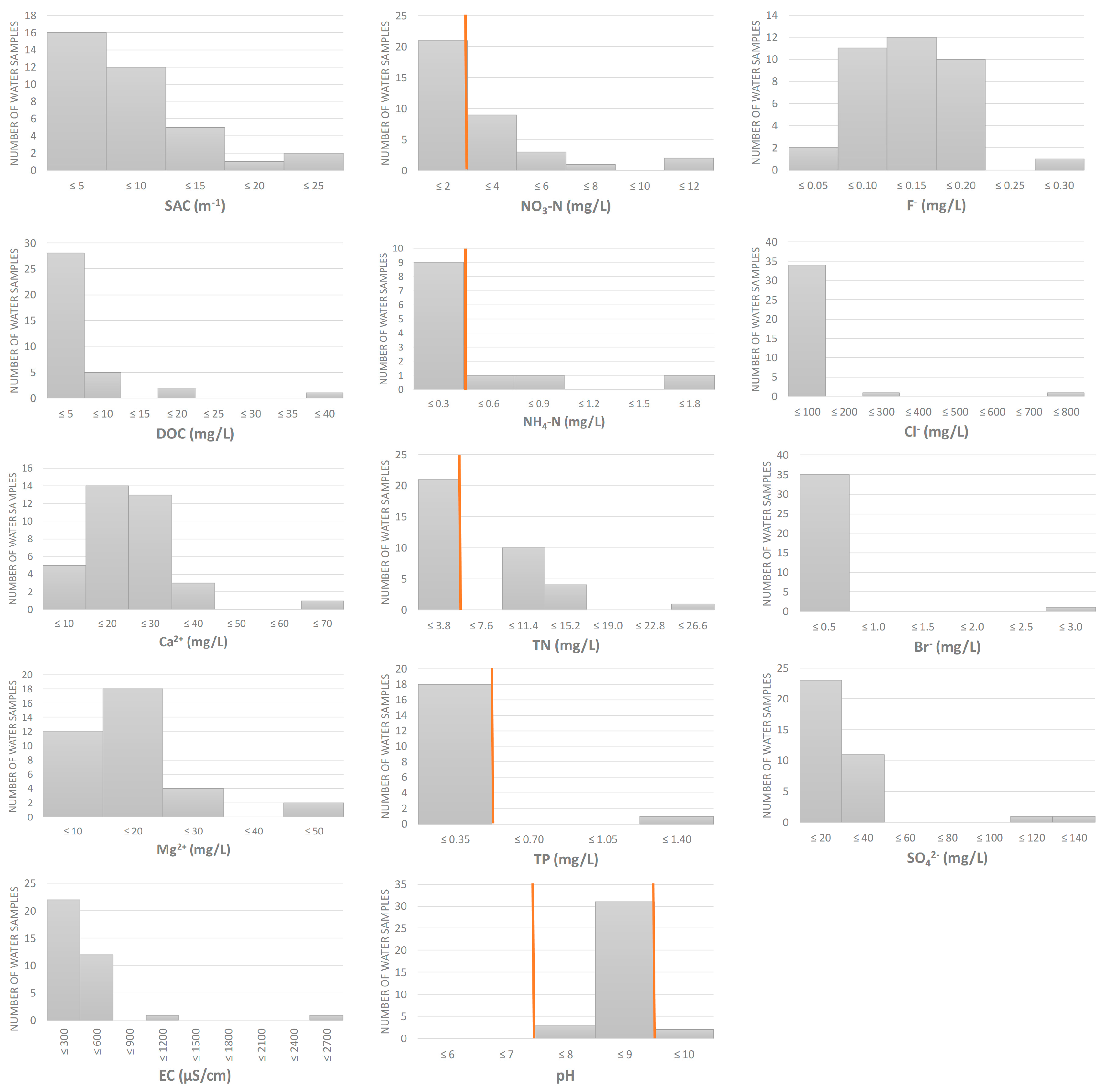
2.2. Physico-Chemical Analyses of Water
2.3. Sporulation
2.4. Data Analysis
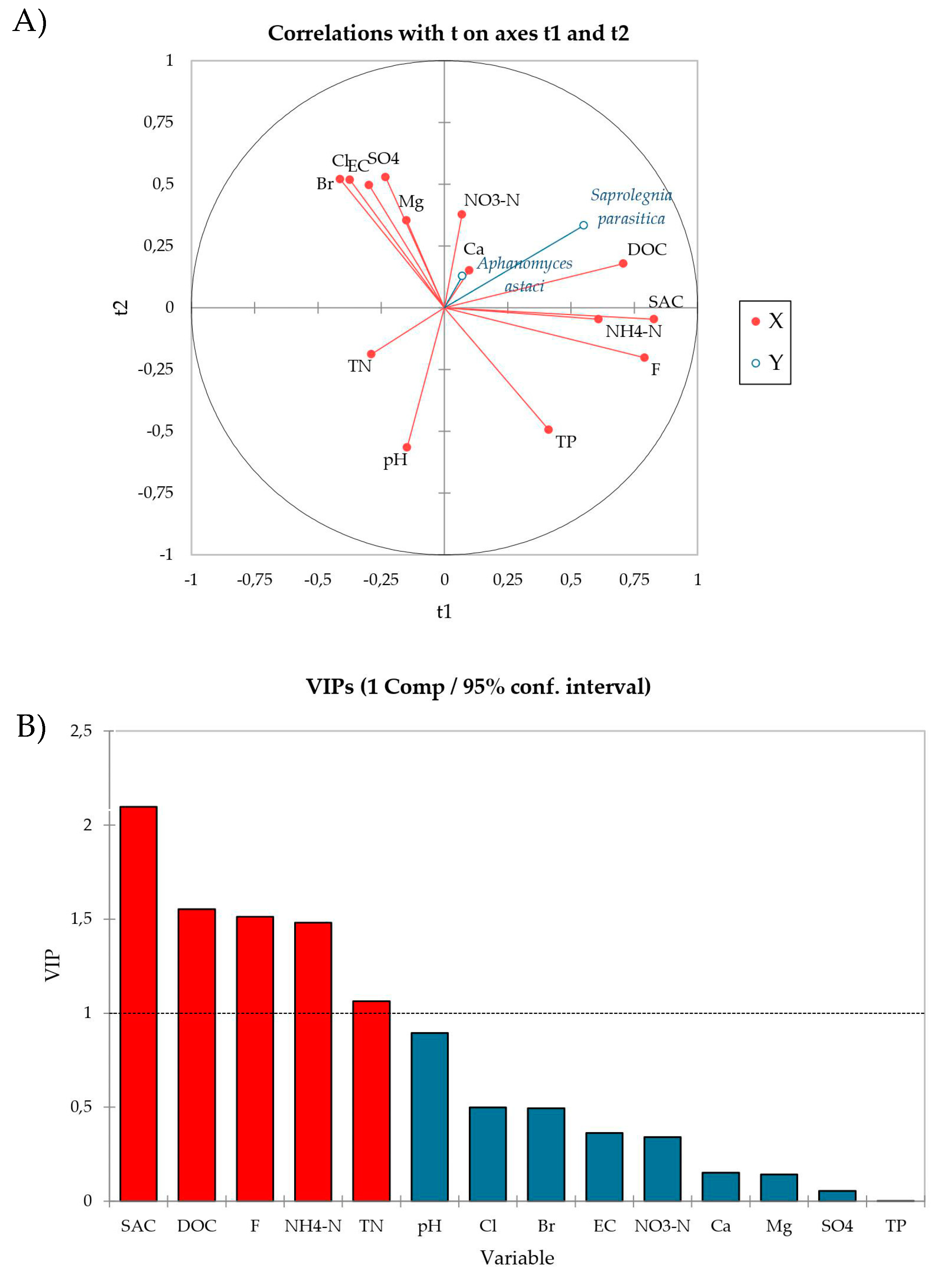
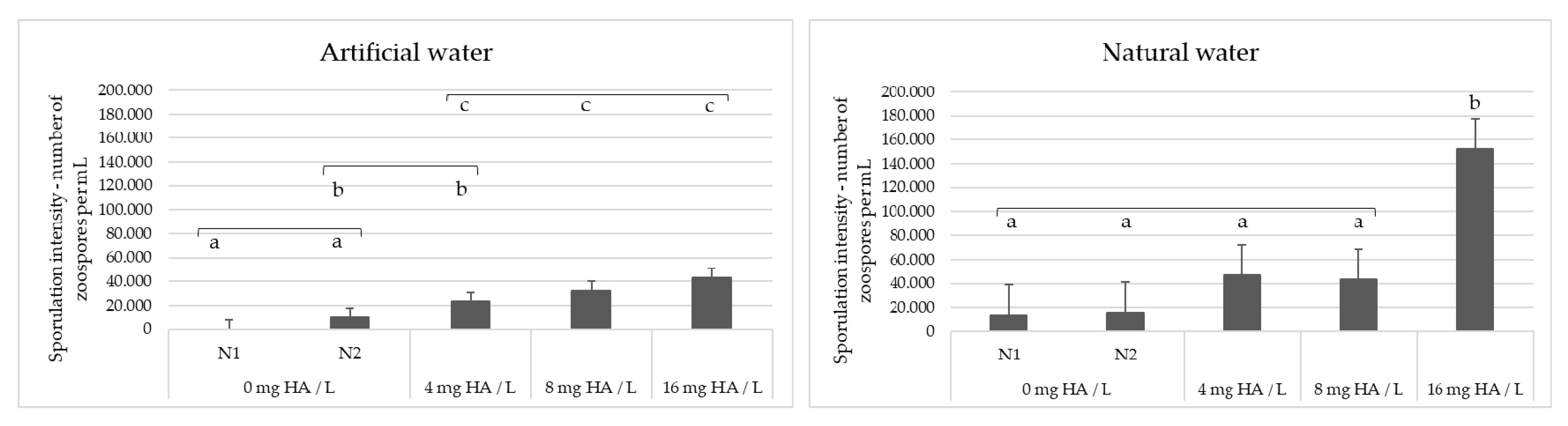
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Derevnina, L.; Petre, B.; Kellner, R.; Dagdas, Y.F.; Sarowar, M.N.; Giannakopoulou, A.; de la Concepcion, J.C.; Chaparro-Garcia, A.; Pennington, H.G.; van West, P.; et al. Emerging oomycete threats to plants and animals. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150459. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.C.; Henk, D.A.; Briggs, C.J.; Brownstein, J.S.; Madoff, L.C.; McCraw, S.L.; Gurr, S.J. Emerging fungal threats to animal, plant and ecosystem health. Nature 2012, 484, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Gozlan, R.E.; Marshall, W.; Lilje, O.; Jessop, C.; Gleason, F.H.; Andreou, D. Current ecological understanding of fungal-like pathogens of fish: What lies beneath? Front. Microbiol. 2014, 5, 1–16. [Google Scholar] [CrossRef] [PubMed]
- van den Berg, A.H.; McLaggan, D.; Diéguez-Uribeondo, J.; van West, P. The impact of the water moulds Saprolegnia diclina and Saprolegnia parasitica on natural ecosystems and the aquaculture industry. Fungal Biol. Rev. 2013, 27, 33–42. [Google Scholar] [CrossRef]
- Diéguez-Uribeondo, J.; Cerenius, L.; Dyková, I.; Gelder, S.; Henttonen, P.; Jiravanichpaisal, P.; Lom, J.; Söderhäll, K. Pathogens, parasites and ectocommensals. In Atlas of Crayfish in European; Souty-Grosset, C., Holdich, D.M., Noël, P.Y., Reynolds, J.D., Haffner, P., Eds.; Muséum National d’Histoire Naturelle: Paris, France, 2006; pp. 131–150. [Google Scholar]
- Söderhäll, K.; Cerenius, L. The crayfish plague fungus: History and recent advances. Freshw. Crayfish 1999, 12, 11–35. [Google Scholar]
- van West, P. Saprolegnia parasitica, an oomycete pathogen with a fishy appetite: New challenges for an old problem. Mycologist 2006, 20, 99–104. [Google Scholar] [CrossRef]
- Holdich, D.M.; Reynolds, J.D.; Souty-Grosset, C.; Sibley, P.J. A review of the ever increasing threat to European crayfish from non-indigenous crayfish species. Knowl. Manag. Aquat. Ecosyst. 2009, 11, 1–46. [Google Scholar] [CrossRef]
- Martín-Torrijos, L.; Kawai, T.; Makkonen, J.; Jussila, J.; Kokko, H.; Diéguez-Uribeondo, J. Crayfish plague in Japan: A real threat to the endemic Cambaroides japonicus. PLoS ONE 2018, 1903, e0195353. [Google Scholar] [CrossRef]
- Peiró, D.F.; Almerão, M.P.; Delaunay, C.; Jussila, J.; Makkonen, J.; Bouchon, D.; Araujo, P.B.; Souty-Grosset, C. First detection of the crayfish plague pathogen Aphanomyces astaci in South America: A high potential risk to native crayfish. Hydrobiologia 2016, 781, 181–190. [Google Scholar] [CrossRef]
- Putra, M.D.; Bláha, M.; Wardiatno, Y.; Krisanti, M.; Jerikho, R.; Kamal, M.M.; Mojžišová, M.; Bystřický, P.K.; Kouba, A.; Kalous, L.; et al. Procambarus clarkii (Girard, 1852) and crayfish plague as new threats for biodiversity in Indonesia. Aquat. Conserv. Mar. Freshw. Ecosyst. 2018, 28, 1434–1440. [Google Scholar] [CrossRef]
- Martín-Torrijos, L.; Jose Correa-Villalona, A.; Juan Carlos Azofeifa-Solano, J.C.; Villalobos-Rojas, F.; Wehrtmann, I.S.; Diéguez-Uribeondo, J. First detection of the crayfish plague pathogen Aphanomyces astaci in Costa Rica: European mistakes should not be repeated. Front. Ecol. Evol. 2021, 9, 376. [Google Scholar] [CrossRef]
- Alderman, D.J. Geographical spread of bacterial and fungal diseases of crustaceans. Rev. Sci. Tech. Int. Epizoot. 1996, 15, 603–632. [Google Scholar] [CrossRef] [PubMed]
- Cerenius, L.; Bangyeekhun, E.; Keyser, P.; Söderhäll, I.; Söderhäll, K. Host prophenoloxidase expression in freshwater crayfish is linked to increased resistance to the crayfish plague fungus, Aphanomyces astaci. Cell. Microbiol. 2003, 5, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Zheng, W.; Xu, J.; Ou, R.; He, S.; Yang, X. Identification of an isolate of Saprolegnia ferax as the causal agent of saprolegniosis of Yellow catfish (Pelteobagrus fulvidraco) eggs. Vet. Res. Commun. 2012, 36, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Thoen, E.; Evensen, Ø.; Skaar, I. Pathogenicity of Saprolegnia spp. to Atlantic salmon, Salmo salar L., eggs. J. Fish Dis. 2011, 34, 601–608. [Google Scholar] [CrossRef]
- Rezinciuc, S.; Sandoval-Sierra, J.V.; Diéguez-Uribeondo, J. Molecular identification of a bronopol tolerant strain of Saprolegnia australis causing egg and fry mortality in farmed brown trout, Salmo trutta. Fungal Biol. 2014, 118, 591–600. [Google Scholar] [CrossRef]
- Hatai, K.; Hoshiai, G. Mass mortality in cultured coho salmon (Oncorhynchus kisutch) due to Saprolegnia parasitica Coker. J. Wildl. Dis. 1992, 28, 532–536. [Google Scholar] [CrossRef]
- Bruno, D.; West, V.P.; Beakes, G. Saprolegnia and other oomycetes. In Fish Diseases and Disorders: Viral, Bacterial and Fungal Infections Volume 3; Woo, P.T.K., Bruno, D.W., Eds.; CABI International: Wallingford, UK, 2011; Volume 3, pp. 669–720. [Google Scholar]
- Ravasi, D.; De Respinis, S.; Wahli, T. Multilocus sequence typing reveals clonality in Saprolegnia parasitica outbreaks. J. Fish Dis. 2018, 41, 1653–1665. [Google Scholar] [CrossRef]
- Pavić, D.; Grbin, D.; Šver, L.; Vladušić, T.; Galuppi, R.; Tedesco, P.; Bielen, A. Identification and molecular characterization of oomycete isolates from trout farms in Croatia, and their upstream and downstream water environments. Aquaculture 2021, 540, 736652. [Google Scholar] [CrossRef]
- Tedesco, P.; Saraiva, M.; Sandoval-Sierra, J.V.; Fioravanti, M.L.; Morandi, B.; Dieguez-Uribeondo, J.; Van West, P.; Galuppi, R. Evaluation of potential transfer of the pathogen Saprolegnia parasitica between farmed salmonids and wild fish. Pathogens 2021, 10, 926. [Google Scholar] [CrossRef]
- Diéguez-Uribeondo, J.; García, M.A.; Cerenius, L.; Kozubíková, E.; Ballesteros, I.; Windels, C.; Weiland, J.; Kator, H.; Söderhäll, K.; Martín, M.P. Phylogenetic relationships among plant and animal parasites, and saprotrophs in Aphanomyces (Oomycetes). Fungal Genet. Biol. 2009, 46, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Phillips, A.J.; Anderson, V.L.; Robertson, E.J.; Secombes, C.J.; van West, P. New insights into animal pathogenic oomycetes. Trends Microbiol. 2008, 16, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Oidtmann, B.; Heitz, E.; Rogers, D.; Hoffmann, R.W. Transmission of crayfish plague. Dis. Aquat. Organ. 2002, 52, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Unestam, T. On the adaptation of Aphanomyces astaci as a parasite. Physiol. Plant. 1969, 22, 221–235. [Google Scholar] [CrossRef]
- Cerenius, L.; Söderhäll, K. Chemotaxis in Aphanomyces astaci, an arthropod-parasitic fungus. J. Invertebr. Pathol. 1984, 43, 278–281. [Google Scholar] [CrossRef]
- Cerenius, L.; Söderhäll, K.; Persson, M.; Ajaxon, R. The crayfish plague fungus Aphanomyces astaci—Diagnosis, isolation and pathobiology. Freshw. Crayfish 1988, 7, 131–144. [Google Scholar]
- Willoughby, L.G. Fungi and Fish Diseases; Pisces Press: Stirling, UK, 1994. [Google Scholar]
- Hatai, K.; Hoshiai, G. Pathogenicity of Saprolegnia parasitica Coker. In Salmon Saprolegniasis; Mueller, G.J., Ed.; U.S. Department of Energy, Bonneville Power Administration: Portland, OR, USA, 1994; pp. 87–98. [Google Scholar]
- Cerenius, L.; Söderhäll, K. Repeated zoospore emergence from isolated spore cysts of Aphanomyces astaci. Exp. Mycol. 1984, 8, 370–377. [Google Scholar] [CrossRef]
- Diéguez-Uribeondo, J.; Cerenius, L.; Söderhäll, K. Repeated zoospore emergence in Saprolegnia parasitica. Mycol. Res. 1994, 98, 810–815. [Google Scholar] [CrossRef]
- Walker, C.A.; van West, P. Zoospore development in the oomycetes. Fungal Biol. Rev. 2007, 21, 10–18. [Google Scholar] [CrossRef]
- Pavić, D.; Bielen, A.; Hudina, S.; Špoljarić, I.; Grandjean, F.; Jussila, J.; Maguire, I. Distribution of Aphanomyces astaci Schikora, 1906, the causative agent of crayfish plague, in the Plitvice Lakes National Park, Croatia. BioInvasions Rec. 2021, 10, 654–668. [Google Scholar] [CrossRef]
- Bly, J.E.; Lawson, L.A.; Dale, D.J.; Szalai, A.J.; Durburow, R.M.; Clem, L.W. Winter saprolegniosis in channel catfish. Dis. Aquat. Organ. 1992, 13, 155–164. [Google Scholar] [CrossRef]
- Unestam, T. On the physiology of zoospore production in Aphanomyces astaci. Physiol. Plant. 1969, 22, 236–246. [Google Scholar] [CrossRef]
- Makkonen, J.; Jussila, J.; Kortet, R.; Vainikka, A.; Kokko, H. Differing virulence of Aphanomyces astaci isolates and elevated resistance of noble crayfish Astacus astacus against crayfish plague. Dis. Aquat. Organ. 2012, 102, 129–136. [Google Scholar] [CrossRef]
- Heungens, K.; Parke, J.L. Zoospore homing and infection events: Effects of the biocontrol bacterium Burkholderia cepacia AMMDR1 on two Oomycete pathogens of pea (Pisum sativum L.). Appl. Environ. Microbiol. 2000, 66, 5192–5200. [Google Scholar] [CrossRef] [PubMed]
- Appiah, A.A.; Van West, P.; Osborne, M.C.; Gow, N.A.R. Potassium homeostasis influences the locomotion and encystment of zoospores of plant pathogenic oomycetes. Fungal Genet. Biol. 2005, 42, 213–223. [Google Scholar] [CrossRef]
- Rantamäki, J.; Cerenius, L.; Söderhäll, K. Prevention of transmission of the crayfish plague fungus (Aphanomyces astaci) to the freshwater crayfish Astacus astacus by treatment with MgCl2. Aquaculture 1992, 104, 11–18. [Google Scholar] [CrossRef]
- Weyhenmeyer, G.A.; Hartmann, J.; Hessen, D.O.; Kopáček, J.; Hejzlar, J.; Jacquet, S.; Hamilton, S.K.; Verburg, P.; Leach, T.H.; Schmid, M.; et al. Widespread diminishing anthropogenic effects on calcium in freshwaters. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Preuett, J.A.; Collins, D.J.; Luster, D.G.; Widmer, T.L. The effect of salinity on the survival, growth, sporulation and infection of Phytophthora ramorum. Fungal Ecol. 2016, 23, 123–130. [Google Scholar] [CrossRef]
- Unestam, T. Studies on the crayfish plague fungus Aphanomyces astaci I. Some factors affecting growth in vitro. Physiol. Plant. 1965, 18, 483–506. [Google Scholar] [CrossRef]
- Min, H.; Hatai, K.; Bai, S. Some inhibitory effects of chitosan on fish-pathogenic oomycete, Saprolegnia parasitica. Fish Pathol. 1998, 29, 73–77. [Google Scholar] [CrossRef]
- Croatian Community (2019) NN 96/19 (Official Gazette 96/19); Status Regulation on the Water Quality. Narodne novine d.d.: Zagreb, Croatia, 9 October 2019.
- Jain, P.; Vineis, P.; Liquet, B.; Vlaanderen, J.; Bodinier, B.; van Veldhoven, K.; Kogevinas, M.; Athersuch, T.J.; Font-Ribera, L.; Villanueva, C.M.; et al. A multivariate approach to investigate the combined biological effects of multiple exposures. J. Epidemiol. Community Health 2018, 72, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Lew, S.; Glińska-Lewczuk, K.; Lew, M. The effects of environmental parameters on the microbial activity in peat-bog lakes. PLoS ONE 2019, 14, e0224441. [Google Scholar] [CrossRef]
- De la Bastide, P.Y.; Leung, W.L.; Hintz, W.E. Species composition of the genus Saprolegnia in fin fish aquaculture environments, as determined by nucleotide sequence analysis of the nuclear rDNA ITS regions. Fungal Biol. 2015, 119, 27–43. [Google Scholar] [CrossRef] [PubMed]
- Neish, G.A.; Hughes, G.C. Diseases of fish. In Fungal Diseases of Fishes; T.W.F. Publications: Neptune, NJ, USA, 1980; p. 159. [Google Scholar]
- Stueland, S.; Hatai, K.; Skaar, I. Morphological and physiological characteristics of Saprolegnia spp. strains pathogenic to Atlantic salmon, Salmo salar L. J. Fish Dis. 2005, 28, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Whisler, H.C. Identification od Saprolegnia spp. Pathogenic in Chinook Salmon; Final Report, DE-AC79-90BP02836; University of Washington: Washington, DC, USA, 1996. [Google Scholar]
- Willoughby, L.G.; Pickering, A.D. Viable Saprolegniaceae spores on the epidermis of the salmonid fish Salmo trutta and Salvelinus alpinus. Trans. Br. Mycol. Soc. 1977, 68, 91–95. [Google Scholar] [CrossRef]
- Bittar, T.B.; Vieira, A.A.H.; Stubbins, A.; Mopper, K. Competition between photochemical and biological degradation of dissolved organic matter from the cyanobacteria microcystis aeruginosa. Limnol. Oceanogr. 2015, 60, 1172–1194. [Google Scholar] [CrossRef]
- Sobek, S.; Tranvik, L.J.; Prairie, Y.T.; Kortelainen, P.; Cole, J.J. Patterns and regulation of dissolved organic carbon: An analysis of 7,500 widely distributed lakes. Limnol. Oceanogr. 2007, 52, 1208–1219. [Google Scholar] [CrossRef]
- Meinelt, T.; Paul, A.; My, T.; Zwirnmann, E.; Kr, A.; Wienke, A.; Steinberg, C.E.W. Reduction in vegetative growth of the water mold Saprolegnia parasitica (Coker) by humic substance of different qualities. Aquat. Toxicol. 2007, 83, 93–103. [Google Scholar] [CrossRef]
- Masigol, H.; Khodaparast, S.A.; Woodhouse, J.N.; Rojas-Jimenez, K.; Fonvielle, J.; Rezakhani, F.; Mostowfizadeh-Ghalamfarsa, R.; Neubauer, D.; Goldhammer, T.; Grossart, H.P. The contrasting roles of aquatic fungi and oomycetes in the degradation and transformation of polymeric organic matter. Limnol. Oceanogr. 2019, 64, 2662–2678. [Google Scholar] [CrossRef]
- Camargo, J.A. Fluoride toxicity to aquatic organisms: A review. Chemosphere 2003, 50, 251–264. [Google Scholar] [CrossRef]
- Marquis, R.E.; Clock, S.A.; Mota-Meira, M. Fluoride and organic weak acids as modulators of microbial physiology. FEMS Microbiol. Rev. 2003, 26, 493–510. [Google Scholar] [CrossRef] [PubMed]
- Mendes, G.; Zafra, D.L.; Vassilev, N.B.; Silva, I.R.; Ribeiro, J.I.J.; Costa, M.D. Biochar enhances Aspergillus niger rock phosphate solubilization by increasing organic acid production and alleviating fluoride toxicity. Appl. Environ. Microbiol. 2014, 80, 3081–3085. [Google Scholar] [CrossRef] [PubMed]
- Montagnolli, R.N.; Lopes, P.R.M.; Cruz, J.M.; Claro, E.M.T.; Quiterio, G.M.; Bidoia, E.D. The effects of fluoride based fire-fighting foams on soil microbiota activity and plant growth during natural attenuation of perfluorinated compounds. Environ. Toxicol. Pharmacol. 2017, 50, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Gao, X.; Li, C.; Luo, X.; Wang, Y. Fluoride contributes to the shaping of microbial community in high fluoride groundwater in Qiji County, Yuncheng City, China. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Carballo, M.; Munoz, M.J.; Cuellar, M.; Tarazona, J.V. Effects of waterborne copper, cyanide, ammonia, and nitrite on stress parameters and changes in susceptibility to saprolegniosis in rainbow trout (Oncorhynchus mykiss). Appl. Environ. Microbiol. 1995, 61, 2108–2112. [Google Scholar] [CrossRef]
- Carballo, M.; Munoz, M.J. Effect of sublethal concentrations of four chemicals on susceptibility of juvenile rainbow trout (Oncorhynchus mykiss) to saprolegniosis. Appl. Environ. Microbiol. 1991, 57, 1813–1816. [Google Scholar] [CrossRef]
- Yan, L.; Mu, X.; Han, B.; Zhang, S.; Qiu, C.; Ohore, O.E. Ammonium loading disturbed the microbial food webs in biofilms attached to submersed macrophyte Vallisneria natans. Sci. Total Environ. 2019, 659, 691–698. [Google Scholar] [CrossRef]
- Diéguez-Uribeondo, J.; Huang, T.-S.; Cerenius, L.; Söderhäll, K. Physiological adaptation of an Aphanomyces astaci strain isolated from the freshwater crayfish Procambarus clarkii. Mycol. Res. 1995, 99, 574–578. [Google Scholar] [CrossRef]
- Diéguez-Uribeondo, J.; Fregeneda-Grandes, J.M.; Cerenius, L.; Pérez-Iniesta, E.; Aller-Gancedo, J.M.; Tellería, M.T.; Söderhäll, K.; Martín, M.P. Re-evaluation of the enigmatic species complex Saprolegnia diclina-Saprolegnia parasitica based on morphological, physiological and molecular data. Fungal Genet. Biol. 2007, 44, 585–601. [Google Scholar] [CrossRef]
- Masigol, H.; Khodaparast, S.A.; Mostowfizadeh-Ghalamfarsa, R.; Rojas-Jimenez, K.; Woodhouse, J.N.; Neubauer, D.; Grossart, H.P. Taxonomical and functional diversity of Saprolegniales in Anzali lagoon, Iran. Aquat. Ecol. 2020, 51, 323–336. [Google Scholar] [CrossRef]
- Bangyeekhun, E.; Quiniou, S.M.A.; Bly, J.E.; Cerenius, L. Characterisation of Saprolegnia sp. isolates from channel catfish. Dis. Aquat. Organ. 2001, 45, 53–59. [Google Scholar] [CrossRef] [PubMed][Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavić, D.; Grbin, D.; Gregov, M.; Ćurko, J.; Vladušić, T.; Šver, L.; Miljanović, A.; Bielen, A. Variations in the Sporulation Efficiency of Pathogenic Freshwater Oomycetes in Relation to the Physico-Chemical Properties of Natural Waters. Microorganisms 2022, 10, 520. https://doi.org/10.3390/microorganisms10030520
Pavić D, Grbin D, Gregov M, Ćurko J, Vladušić T, Šver L, Miljanović A, Bielen A. Variations in the Sporulation Efficiency of Pathogenic Freshwater Oomycetes in Relation to the Physico-Chemical Properties of Natural Waters. Microorganisms. 2022; 10(3):520. https://doi.org/10.3390/microorganisms10030520
Chicago/Turabian StylePavić, Dora, Dorotea Grbin, Marija Gregov, Josip Ćurko, Tomislav Vladušić, Lidija Šver, Anđela Miljanović, and Ana Bielen. 2022. "Variations in the Sporulation Efficiency of Pathogenic Freshwater Oomycetes in Relation to the Physico-Chemical Properties of Natural Waters" Microorganisms 10, no. 3: 520. https://doi.org/10.3390/microorganisms10030520
APA StylePavić, D., Grbin, D., Gregov, M., Ćurko, J., Vladušić, T., Šver, L., Miljanović, A., & Bielen, A. (2022). Variations in the Sporulation Efficiency of Pathogenic Freshwater Oomycetes in Relation to the Physico-Chemical Properties of Natural Waters. Microorganisms, 10(3), 520. https://doi.org/10.3390/microorganisms10030520