
Assessing the Tsetse Fly Microbiome Composition and the Potential Association of Some Bacteria Taxa with Trypanosome Establishment

 ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Tsetse Collection and Preservation
2.3. DNA Extraction
2.4. PCR Detection of Trypanosomes
2.5. Determination of Flies’ Microbiome Composition
2.5.1. Library Preparation and Sequencing
2.5.2. Sequence Data Processing
2.5.3. Statistical Analyses
3. Results
3.1. Tsetse Fly Collection and Trypanosome-Infection Status
3.2. Bacterial 16S rRNA Sequencing Quality and Rarefaction
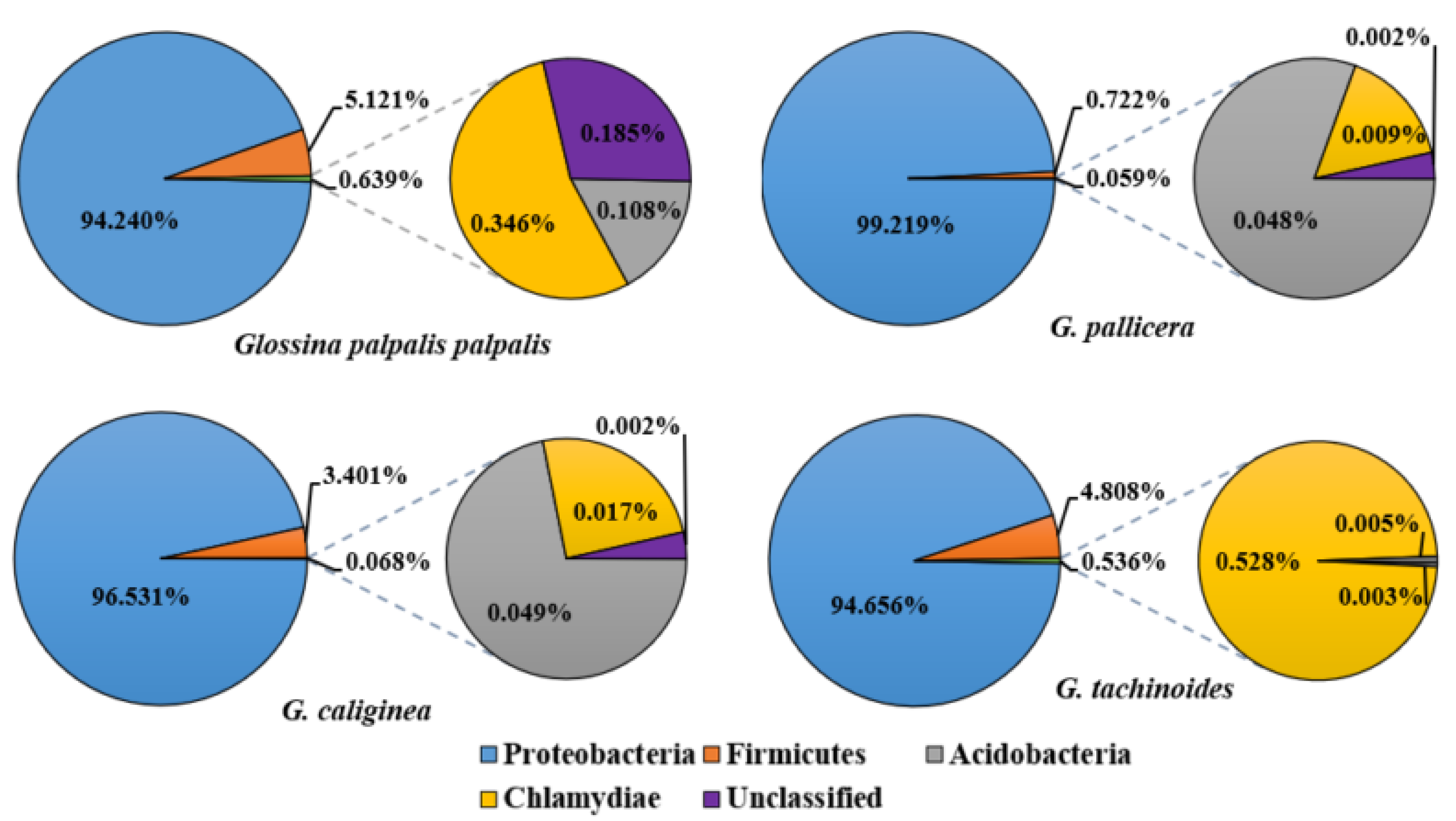
3.3. General Characterization of Bacterial Phyla and Abundances in Tsetse
3.4. Tsetse Microbiome Composition at Genus Level
3.5. Bacterial Genera Diversity in Tsetse Flies (Alpha-Diversity)
3.5.1. Diversity in Tsetse Species
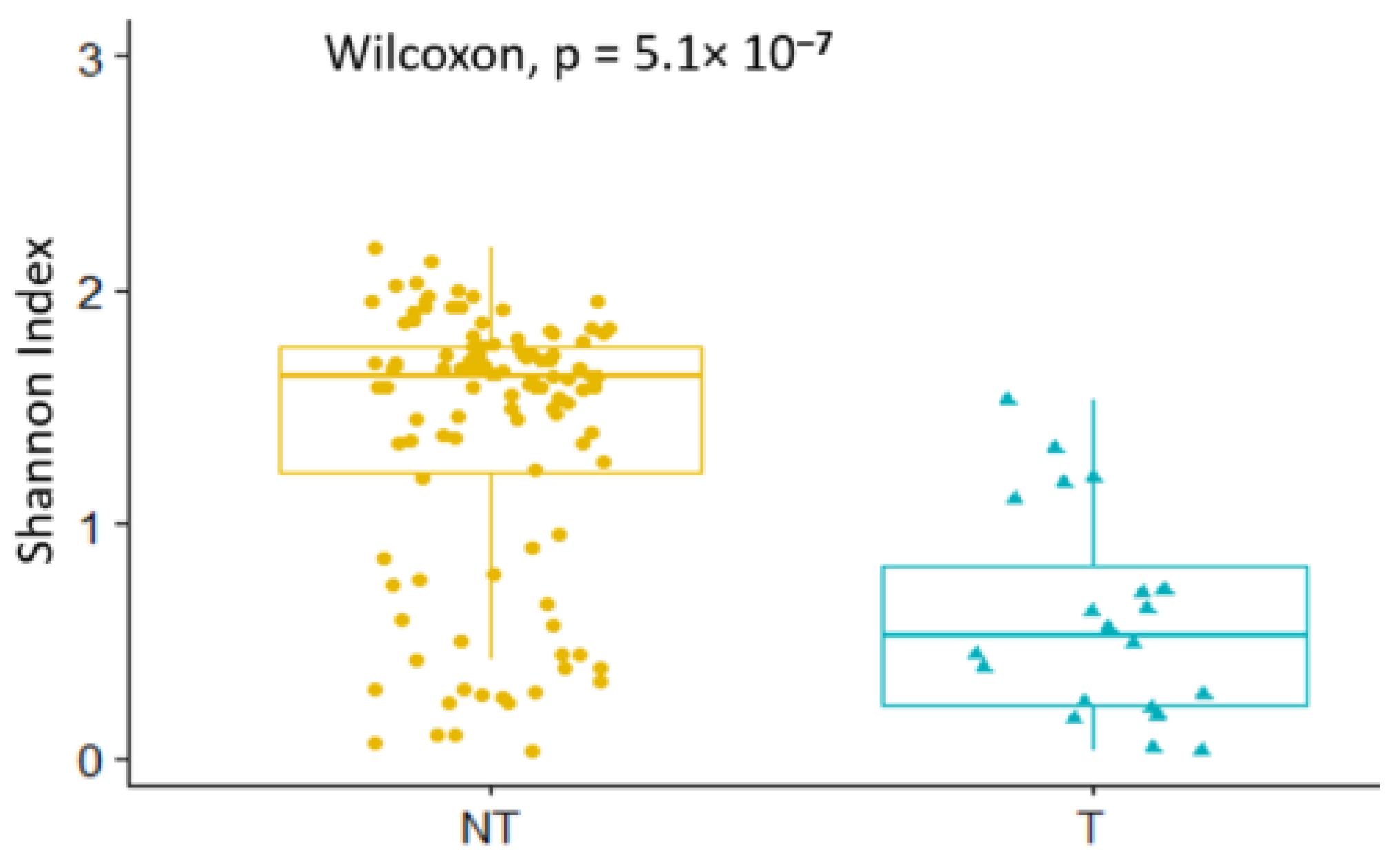
3.5.2. Diversity in Non-Teneral and Teneral Glossina palpalis palpalis
3.6. Multivariate Analysis (Beta Diversity)
3.7. Bacterial Communities in Glossina palpalis palpalis Infected and Non-Infected Flies
4. Discussion
4.1. Abundances of Bacterial Taxa in Whole Tsetse Fly Bodies
4.2. General Microbiome Diversity in Tsetse Flies
4.3. Microbiome of Trypanosome Infected and Non-Infected Glossina Palpalis Palpalis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kennedy, P.G.E. Clinical features, diagnosis, and treatment of human African trypanosomiasis (sleeping sickness). Lancet Neurol. 2013, 12, 186–194. [Google Scholar] [CrossRef]
- Cecchi, R.; Mattioli, G. Geospatial datasets and analyses for an environmental approach to African trypanosomiasis. In Programme against African Trypanosomiasis; FAO: Rome, Italy, 2009; pp. 1–80. [Google Scholar]
- Morrison, L.J.; Vezza, L.; Rowan, T.; Hope, J.C. Animal African Trypanosomiasis: Time to Increase Focus on Clinically Relevant Parasite and Host Species. Trends Parasitol. 2016, 32, 599–607. [Google Scholar] [CrossRef] [PubMed]
- WHO. Number of New Cases of Human African Trypanosomiases (T. b. Gambiense): 2020 [Internet]. 2021. Available online: https://apps.who.int/neglected_diseases/ntddata/hat/hat.html (accessed on 10 November 2021).
- Lumbala, C.; Biéler, S.; Kayembe, S.; Makabuza, J.; Ongarello, S.; Ndung’u, J.M. Prospective evaluation of a rapid diagnostic test for Trypanosoma brucei gambiense infection developed using recombinant antigens. PLoS Negl. Trop. Dis. 2018, 12, e0006386. [Google Scholar] [CrossRef] [PubMed]
- Tirados, I.; Esterhuizen, J.; Kovacic, V.; Mangwiro, T.N.C.; Vale, G.A.; Hastings, I.; Solano, P.; Lehane, M.J.; Torr, S.J. Tsetse control and Gambian sleeping sickness; implications for control strategy. PLoS Negl. Trop. Dis. 2015, 9, e0003822. [Google Scholar] [CrossRef]
- Nerima, B.; Matovu, E.; Lubega, G.W.; Enyaru, J.C.K.K. Detection of mutant P2 adenosine transporter (TbAT1) gene in Trypanosoma brucei gambiense isolates from northwest Uganda using allele-specific polymerase chain reaction. Trop. Med. Int. Health 2007, 12, 1361–1368. [Google Scholar] [CrossRef]
- Chitanga, S.; Marcotty, T.; Namangala, B.; van den Bossche, P.; van den Abbeele, J.; Delespaux, V. High prevalence of drug resistance in animal trypanosomes without a history of drug exposure. PLoS Negl. Trop. Dis. 2011, 5, e1454. [Google Scholar] [CrossRef]
- Van Den Abbeele, J.; Claes, Y.; Van Bockstaele, D.; Le Ray, D.; Coosemans, M. Trypanosoma brucei spp. development in the tsetse fly: Characterization of the post-mesocyclic stages in the foregut and proboscis. Parasitology 1999, 118, 469–478. [Google Scholar] [CrossRef]
- Aksoy, S. Wigglesworthia and Wigglesworthia glossinidia sp. taxa consisting of the mycetocyte-associated, primary endosymbionts of tsetse flies. Infect. J. Syst. Bacteriol. 1995, 45, 848–851. [Google Scholar] [CrossRef]
- Alam, U.; Medlock, J.; Brelsfoard, C.; Pais, R.; Lohs, C.; Balmand, S.; Carnogursky, J.; Heddi, A.; Takac, P.; Galvani, A.; et al. Wolbachia symbiont infections induce strong cytoplasmic incompatibility in the Tsetse fly Glossina morsitans. PLoS Pathog. 2011, 7, e1002415. [Google Scholar] [CrossRef]
- Balmand, S.; Lohs, C.; Aksoy, S.; Heddi, A. Tissue distribution and transmission routes for the tsetse fly endosymbionts. J. Invertebr. Pathol. 2013, 112 (Suppl. 1), S116–S122. [Google Scholar] [CrossRef]
- Geiger, A.; Ravel, S.; Mateille, T.; Janelle, J.; Patrel, D.; Cuny, G.; Frutos, R. Vector competence of Glossina palpalis gambiensis for Trypanosoma brucei s.l. and genetic diversity of the symbiont Sodalis glossinidius. Mol. Biol. Evol. 2007, 24, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Welburn, S.C.; Maudlin, I. Tsetse—Trypanosome Interactions: Rites of Passage. Parasitol. Today 1999, 4758, 399–403. [Google Scholar] [CrossRef]
- Dale, C.; Young, S.A.; Haydon, D.T.; Welburn, S.C. The insect endosymbiont Sodalis glossinidius utilizes a type III secretion system for cell invasion. Proc. Natl. Acad. Sci. USA 2001, 98, 1883–1888. [Google Scholar] [CrossRef] [PubMed]
- Weiss, B.L.; Maltz, M.A.; Vigneron, A.; Wu, Y.; Walter, K.S.; O’Neill, M.B.; Wang, J.; Aksoy, S. Colonization of the tsetse fly midgut with commensal Kosakonia cowanii Zambiae inhibits trypanosome infection establishment. PLoS Pathog. 2019, 15, e1007470. [Google Scholar] [CrossRef]
- Jacob, F.; Melachio, T.T.; Njitchouang, G.R.; Gimonneau, G.; Njiokou, F.; Abate, L.; Christen, R.; Reveillaud, J.; Geiger, A. Intestinal Bacterial Communities of Trypanosome-Infected and Uninfected Glossina palpalis palpalis from Three Human African Trypanomiasis Foci in Cameroon. Front. Microbiol. 2017, 8, 1464. [Google Scholar] [CrossRef]
- Kame-Ngasse, G.I.; Geiger, A.; Simo, G.; Melachio-Tanekou, T.T.; Farikou, O.; Njiokou, F. Prevalence of symbionts and trypanosome infections in tsetse flies of two villages of the “Faro and Déo” division of the Adamawa region of Cameroon. BMC Microbiol. 2018, 18, 83–91. [Google Scholar] [CrossRef]
- Kanté Tagueu, S.; Farikou, O.; Njiokou, F.; Simo, G. Prevalence of Sodalis glossinidius and different trypanosome species in Glossina palpalis palpalis caught in the Fontem sleeping sickness focus of the southern Cameroon. Parasite 2018, 25, 44. [Google Scholar] [CrossRef]
- Tsagmo Ngoune, J.M.; Reveillaud, J.; Sempere, G.; Njiokou, F.; Melachio, T.T.; Abate, L.; Tchioffo, M.T.; Geiger, A. The composition and abundance of bacterial communities residing in the gut of Glossina palpalis palpalis captured in two sites of southern Cameroon. Parasites Vectors 2019, 12, 151. [Google Scholar] [CrossRef]
- Ngambia Freitas, F.S.; Njiokou, F.; Tsagmo Ngoune, J.M.; Sempere, G.; Berthier, D.; Geiger, A. Modulation of trypanosome establishment in Glossina palpalis palpalis by its microbiome in the Campo sleeping sickness focus, Cameroon. Infect. Genet. Evol. 2021, 90, 104763. [Google Scholar] [CrossRef]
- Hamidou Soumana, I.; Simo, G.; Njiokou, F.; Tchicaya, B.; Abd-Alla, A.M.M.; Cuny, G.; Geiger, A. The bacterial flora of tsetse fly midgut and its effect on trypanosome transmission. J. Invertebr. Pathol. 2013, 112 (Suppl. 1), S89–S93. [Google Scholar] [CrossRef]
- Geiger, A.; Fardeau, M.-L.; Njiokou, F.; Ollivier, B.; Runyen-Janecky, L.; Rio, R. Glossina spp. gut bacterial flora and their putative role in fly-hosted trypanosome development. Front. Cell. Infect. Microbiol. 2013, 3, 34. [Google Scholar] [CrossRef] [PubMed]
- De Vooght, L.; Caljon, G.; De Ridder, K.; Van Den Abbeele, J. Delivery of a functional anti-trypanosome Nanobody in different tsetse fly tissues via a bacterial symbiont, Sodalis glossinidius. Microb. Cell Fact. 2014, 13, 156. [Google Scholar] [CrossRef]
- Romoli, O.; Gendrin, M. The tripartite interactions between the mosquito, its microbiota and Plasmodium. Parasites Vectors 2018, 11, 200. [Google Scholar] [CrossRef] [PubMed]
- Njiokou, F.; Laveissière, C.; Simo, G.; Nkinin, S.; Grébaut, P.; Cuny, G.; Herder, S. Wild fauna as a probable animal reservoir for Trypanosoma brucei gambiense in Cameroon. Infect. Genet. Evol. 2006, 6, 147–153. [Google Scholar] [CrossRef]
- Simo, G.; Njiokou, F.; Mbida Mbida, J.A.; Njitchouang, G.R.; Herder, S.; Asonganyi, T.; Cuny, G. Tsetse fly host preference from sleeping sickness foci in Cameroon: Epidemiological implications. Infect. Genet. Evol. 2008, 8, 34–39. [Google Scholar] [CrossRef]
- Grébaut, P.; Melachio, T.; Nyangmang, S.; Eyenga, V.E.; Njitchouang, G.-R.; Ofon, E.; Njiokou, F.; Simo, G. Xenomonitoring of sleeping sickness transmission in Campo (Cameroon). Parasit. Vectors 2016, 9, 201. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gouteux, P.; Lancien, J. Le piege pyramidal a tsé-tsé (Diptera: Glossinidae) pour la capture et la lutte Essais comparatifs et description de nouveaux systèmes de capture. Trop. Med. Parasitol. 1986, 37, 61–66. [Google Scholar]
- Livak, K.J. Organization and mapping of a sequence on the Drosophila melanogaster X and Y chromosomes that is transcribed during spermatogenesis. Genetics 1984, 107, 611–634. [Google Scholar] [CrossRef]
- Desquesnes, M.; McLaughlin, G.; Zoungrana, A.; Dávila, A.M. Detection and identification of Trypanosoma of African livestock through a single PCR based on internal transcribed spacer 1 of rDNA. Int. J. Parasitol. 2001, 31, 610–614. [Google Scholar] [CrossRef]
- Klindworth, A.; Quast, C.; Pruesse, E.; Schweer, T.; Horn, M.; Glo, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Illumina. 16S Metagenomic Sequencing Library Preparation; Illumina: San Diego, CA, USA, 2013; Available online: https://support.illumina.com/documents/documentation/chemistry_documentation/16s/16s-metagenomic-library-prep-guide-15044223-b.pdf (accessed on 10 December 2020).
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a Dual-Index Sequencing Strategy and Curation Pipeline for Analyzing Amplicon Sequence Data on the MiSeq Illumina Sequencing Platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2020; Available online: https://www.r-project.org/ (accessed on 10 January 2021).
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Callahan, B.J.; Sankaran, K.; Fukuyama, J.A.; McMurdie, P.J.; Holmes, S.P. Bioconductor workflow for microbiome data analysis: From raw reads to community analyses [version 1; referees: 3 approved]. F1000Research 2016, 5, 1492. [Google Scholar] [CrossRef]
- Wickham, H. Elegant Graphics for Data Analysis: Ggplot2. In Applied Spatial Data Analysis with R; Springer: New York, NY, USA, 2008; pp. 21–54. [Google Scholar]
- Kassambara, A. Ggpubr: “Ggplot2” Based Publication Ready Plots [Internet]. 2020. Available online: https://rpkgs.datanovia.com/ggpubr/ (accessed on 12 February 2021).
- Oksanen, A.J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; Hara, R.B.O.; Simpson, G.L.; Solymos, P.; et al. Vegan. In Encyclopedia of Food and Agricultural Ethics; Springer: Dordrecht, The Netherlands, 2019; pp. 2395–2396. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Paradis, E.; Schliep, K. Ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- Galili, T. Dendextend: An R package for visualizing, adjusting and comparing trees of hierarchical clustering. Bioinformatics 2015, 31, 3718–3720. [Google Scholar] [CrossRef]
- Xie, Y. Knitr: A General-Purpose Package for Dynamic Report Generation in R. R Package Version 1.36 [Internet]. 2021. Available online: https://yihui.org/knitr/ (accessed on 12 February 2021).
- Geiger, A.; Fardeau, M.L.; Njiokou, F.; Joseph, M.; Asonganyi, T.; Ollivier, B.; Cuny, G. Bacterial Diversity Associated with Populations of Glossina spp. from Cameroon and Distribution within the Campo Sleeping Sickness Focus. Microb. Ecol. 2011, 62, 632–643. [Google Scholar] [CrossRef]
- Yabu, Y.; Takayanagi, T. Trypsin-stimulated transformation of Trypanosoma brucei gambiense bloodstream forms to procyclic forms in vitro. Parasitol. Res. 1988, 74, 501–506. [Google Scholar] [CrossRef]
- Compant, S.; Nowak, J.; Coenye, T.; Clément, C.; Ait Barka, E. Diversity and occurrence of Burkholderia spp. in the natural environment. FEMS Microbiol. Rev. 2008, 32, 607–626. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Meng, X.Y.; Fukatsu, T. Gut symbiotic bacteria of the genus Burkholderia in the broad-headed bugs Riptortus clavatus and Leptocorisa chinensis (Heteroptera: Alydidae). Appl. Environ. Microbiol. 2005, 71, 4035–4043. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Hosokawa, T.; Fukatsu, T. Insect-microbe mutualism without vertical transmission: A stinkbug acquires a beneficial gut symbiont from the environment every generation. Appl. Environ. Microbiol. 2007, 73, 4308–4316. [Google Scholar] [CrossRef]
- Solano, P.; Salou, E.; Rayaisse, J.B.; Ravel, S.; Gimonneau, G.; Traore, I.; Bouyer, J. Do tsetse flies only feed on blood? Infect. Genet. Evol. 2015, 36, 184–189. [Google Scholar] [CrossRef]
- Colman, D.R.; Toolson, E.C.; Takacs-Vesbach, C.D. Do diet and taxonomy influence insect gut bacterial communities? Mol. Ecol. 2012, 21, 5124–5137. [Google Scholar] [CrossRef]
- Boissière, A.; Tchioffo, M.T.; Bachar, D.; Abate, L.; Marie, A.; Nsango, S.E.; Shahbazkia, H.R.; Awono-Ambene, P.H.; Levashina, E.A.; Christen, R.; et al. Midgut microbiota of the malaria mosquito vector Anopheles gambiae and interactions with Plasmodium falciparum infection. PLoS Pathog. 2012, 8, e1002742. [Google Scholar] [CrossRef]
- Santos, A.V.; Dillon, R.J.; Dillon, V.M.; Reynolds, S.E.; Samuels, R.I. Ocurrence of the antibiotic producing bacterium Burkholderia sp. in colonies of the leaf-cutting ant Atta sexdens rubropilosa. FEMS Microbiol. Lett. 2004, 239, 319–323. [Google Scholar] [CrossRef]
- Dinparast Djadid, N.; Jazayeri, H.; Raz, A.; Favia, G.; Ricci, I.; Zakeri, S. Identification of the midgut microbiota of An. stephensi and An. maculipennis for their application as a paratransgenic tool against malaria. PLoS ONE 2011, 6, 6–12. [Google Scholar] [CrossRef]
- Aksoy, E.; Telleria, E.L.; Echodu, R.; Wu, Y.; Okedi, L.M.; Weiss, B.L.; Aksoy, S.; Caccone, A. Analysis of multiple tsetse fly populations in Uganda reveals limited diversity and species-specific gut microbiota. Appl. Environ. Microbiol. 2014, 80, 4301–4312. [Google Scholar] [CrossRef]
- Muturi, E.J.; Ramirez, J.L.; Rooney, A.P.; Kim, C.H. Comparative analysis of gut microbiota of mosquito communities in central Illinois. PLoS Negl. Trop. Dis. 2017, 11, e0005377. [Google Scholar] [CrossRef]
- Heise, S.R.; Elshahed, M.S.; Little, S.E. Bacterial diversity in Amblyomma americanum (Acari: Ixodidae) with a focus on members of the genus Rickettsia. J. Med. Entomol. 2010, 47, 258–268. [Google Scholar] [CrossRef]
- Coburn, P.S.; Gilmore, M.S. The Enterococcus faecalis cytolysin: A novel toxin active against eukaryotic and prokaryotic cells. Cell. Microbiol. 2003, 5, 661–669. [Google Scholar] [CrossRef]
- Lazaro, J.E.H.; Nitcheu, J.; Predicala, R.Z.; Mangalindan, G.C.; Nesslany, F.; Marzin, D.; Concepcion, G.P.; Diquet, B. Heptyl prodigiosin, a bacterial metabolite, is antimalarial in vivo and non-mutagenic in vitro. J. Nat. Toxins 2002, 11, 367–377. [Google Scholar]
- Azambuja, P.; Feder, D.; Garcia, E.S. Isolation of Serratia marcescens in the midgut of Rhodnius prolixus: Impact on the establishment of the parasite Trypanosoma cruzi in the vector. Exp. Parasitol. 2004, 107, 89–96. [Google Scholar] [CrossRef]
- Apte-Deshpande, A.D.; Paingankar, M.S.; Gokhale, M.D.; Deobagkar, D.N. Serratia odorifera mediated enhancement in susceptibility of Aedes aegypti for chikungunya virus. Indian J. Med. Res. 2014, 139, 762–768. [Google Scholar]
- Apte-Deshpande, A.; Paingankar, M.; Gokhale, M.D.; Deobagkar, D.N. Serratia odorifera a midgut inhabitant of aedes aegypti mosquito enhances its susceptibility to dengue-2 virus. PLoS ONE 2012, 7, e40401. [Google Scholar] [CrossRef]
- Fouda, M.A.; Hassan, M.I.; Al-Daly, A.G.; Hammad, K.M. Effect of midgut bacteria of Culex pipiens L. on digestion and reproduction. J. Egypt. Soc. Parasitol. 2001, 31, 767–780. [Google Scholar]
- König, H. Bacillus species in the intestine of termites and other soil invertebrates. J. Appl. Microbiol. 2006, 101, 620–627. [Google Scholar] [CrossRef]
- Peck, G.W.; Walton, W.E. Effect of bacterial quality and density on growth and whole body stoichiometry of Culex quinquefasciatus and Culex tarsalis (Diptera: Culicidae). J. Med. Entomol. 2006, 43, 25–33. [Google Scholar] [CrossRef]
- Doudoumis, V.; Tsiamis, G.; Wamwiri, F.; Brelsfoard, C.; Alam, U.; Aksoy, E.; Dalaperas, S.; Abd-Alla, A.; Ouma, J.; Takac, P.; et al. Detection and characterization of Wolbachia infections in laboratory and natural populations of different species of tsetse flies (genus Glossina). BMC Microbiol. 2012, 12 (Suppl. 1), S3. [Google Scholar] [CrossRef]
- Farikou, O.; Njiokou, F.; Mbida Mbida, J.A.; Njitchouang, G.R.; Djeunga, H.N.; Asonganyi, T.; Simarro, P.P.; Cuny, G.; Geiger, A. Tripartite interactions between tsetse flies, Sodalis glossinidius and trypanosomes—An epidemiological approach in two historical human African trypanosomiasis foci in Cameroon. Infect. Genet. Evol. 2010, 10, 115–121. [Google Scholar] [CrossRef]
- Hamidou Soumana, I.; Loriod, B.; Ravel, S.; Tchicaya, B.; Simo, G.; Rihet, P.; Geiger, A. The transcriptional signatures of Sodalis glossinidius in the Glossina palpalis gambiensis flies negative for Trypanosoma brucei gambiense contrast with those of this symbiont in tsetse flies positive for the parasite: Possible involvement of a Sodalis-hosted prophage in fly Trypanosoma refractoriness? Infect. Genet. Evol. 2014, 24, 41–56. [Google Scholar] [CrossRef]
- Stouthamer, R.; Breeuwer, J.A.J.; Hurst, G.D.D. Wolbachia pipientis: Microbial manipulator of arthropod reproduction. Annu. Rev. Microbiol. 1999, 53, 71–102. [Google Scholar] [CrossRef]
- Blagrove, M.S.C.; Arias-Goeta, C.; Failloux, A.B.; Sinkins, S.P. Wolbachia strain wMel induces cytoplasmic incompatibility and blocks dengue transmission in Aedes albopictus. Proc. Natl. Acad. Sci. USA 2012, 109, 255–260. [Google Scholar] [CrossRef]
- Chen, X.; Li, S.; Aksoy, S. Concordant evolution of a symbiont with its host insect species: Molecular phylogeny of genus Glossina and its bacteriome-associated endosymbiont, Wigglesworthia glossinidia. J. Mol. Evol. 1999, 48, 49–58. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | G. cal. % | G. pal. % | G. p. p. % | G. tach % | Total Abundance (%) |
|---|---|---|---|---|---|
| Wigglesworthia | 47.61 | 65.64 | 42.13 | 62.61 | 47.29 |
| Serratia | 21.70 | 8.95 | 18.82 | 4.33 | 16.58 |
| Pantoae_Klebsiella_ Enterobacter_Kluyvera | 8.10 | 14.81 | 19.45 | 5.18 | 16.30 |
| Pseudomonas | 11.95 | 4.80 | 6.16 | 1.69 | 6.16 |
| Staphylococcus | 2.96 | 0.69 | 3.96 | 2.43 | 3.36 |
| Acinetobacter | 1.78 | 2.35 | 2.85 | 3.62 | 2.77 |
| Stenotrophomonas | 3.97 | 1.58 | 2.06 | 0.39 | 2.03 |
| Burkholderia | 0.01 | 0 | 0.03 | 13.02 | 1.38 |
| Others | 1.87 | 1.14 | 4.49 | 6.69 | 4.10 |
| OTUs | Genus (Percentage/Num OTUs in the Genus) * | Base Mean | log2 Fold Change | lfcSE | Statistic | p-Value |
|---|---|---|---|---|---|---|
| OTU37 | Dechloromonas (100/1) | 21.44 | −25.44 | 2.3 | −11.06 | 0.0000 |
| OTU49 | Ralstonia (100/1) | 19.88 | −10.58 | 1.67 | −6.33 | 0.0000 |
| OTU14 | Listeria (100/1) | 283.11 | −5.41 | 1.08 | −5.01 | 0.0000 |
| OTU28 | Aquabacterium (79.12/2) | 46.11 | −7.76 | 1.99 | −3.9 | 0.0001 |
| OTU25 | Bacillus (53.62/3) | 38 | −2.98 | 0.9 | −3.31 | 0.0009 |
| OTU24 | Methylophilus (24.65/2) | 11.6 | −6.3 | 2.25 | −2.8 | 0.0051 |
| OTU70 | Kinneretia_ Roseateles_ Pelomonas_Mitsuaria (100/1) | 11.36 | −4.37 | 1.59 | −2.74 | 0.0061 |
| OTU33 | Pseudomonas_ Escherichia (100/1) | 9.31 | −2.98 | 1.19 | −2.51 | 0.0120 |
| OTU45 | Pseudomonas (0.36/3) | 4.66 | −5 | 2.11 | −2.37 | 0.0179 |
| OTU63 | Cupriavidus (40.19/2) | 3.25 | −4.82 | 2.18 | −2.21 | 0.0273 |
| OTU27 | Enterococcus (70.41/3) | 5.99 | −5.36 | 2.52 | −2.13 | 0.0334 |
| OTU18 | Wigglesworthia (0.25/2) | 15.97 | 4.57 | 0.84 | 5.43 | 0.0000 |
| OTU55 | Serratia (0.14/2) | 10.16 | 3.39 | 0.96 | 3.51 | 0.0004 |
| OTU40 | Staphylococcus (1.85/7) | 44.63 | 5.48 | 1.89 | 2.9 | 0.0037 |
| OTUs | Genus (Percentage/Num OTUs in the Genus) * | Base Mean | log2 Fold Change | lfcSE | Statistic | p-Value |
|---|---|---|---|---|---|---|
| OTU29 | Staphylococcus (6.19/7) | 7.85 | −24.84 | 2.92 | -8.51 | 0.0000 |
| OTU55 | Serratia (0.14/2) | 1.59 | −3.02 | 1.32 | -2.28 | 0.0224 |
| OTU49 | Ralstonia (100/1) | 21.57 | −3.88 | 1.77 | -2.19 | 0.0286 |
| OTU16 | Enhydrobacter (100/1) | 238.65 | 2.66 | 0.84 | 3.15 | 0.0016 |
| OTU25 | Bacillus (53.62/3) | 37.31 | 2.35 | 1.07 | 2.19 | 0.0287 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsakeng, C.U.B.; Tanekou, T.T.M.; Soffack, S.F.; Tirados, I.; Noutchih, C.; Njiokou, F.; Bigoga, J.D.; Wondji, C.S. Assessing the Tsetse Fly Microbiome Composition and the Potential Association of Some Bacteria Taxa with Trypanosome Establishment. Microorganisms 2022, 10, 1141. https://doi.org/10.3390/microorganisms10061141
Tsakeng CUB, Tanekou TTM, Soffack SF, Tirados I, Noutchih C, Njiokou F, Bigoga JD, Wondji CS. Assessing the Tsetse Fly Microbiome Composition and the Potential Association of Some Bacteria Taxa with Trypanosome Establishment. Microorganisms. 2022; 10(6):1141. https://doi.org/10.3390/microorganisms10061141
Chicago/Turabian StyleTsakeng, Calmes Ursain Bouaka, Tito Tresor Melachio Tanekou, Steve Feudjio Soffack, Inaki Tirados, Cedrique Noutchih, Flobert Njiokou, Jude Daiga Bigoga, and Charles Sinclair Wondji. 2022. "Assessing the Tsetse Fly Microbiome Composition and the Potential Association of Some Bacteria Taxa with Trypanosome Establishment" Microorganisms 10, no. 6: 1141. https://doi.org/10.3390/microorganisms10061141
APA StyleTsakeng, C. U. B., Tanekou, T. T. M., Soffack, S. F., Tirados, I., Noutchih, C., Njiokou, F., Bigoga, J. D., & Wondji, C. S. (2022). Assessing the Tsetse Fly Microbiome Composition and the Potential Association of Some Bacteria Taxa with Trypanosome Establishment. Microorganisms, 10(6), 1141. https://doi.org/10.3390/microorganisms10061141