
Dark Diazotrophy during the Late Summer in Surface Waters of Chile Bay, West Antarctic Peninsula

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
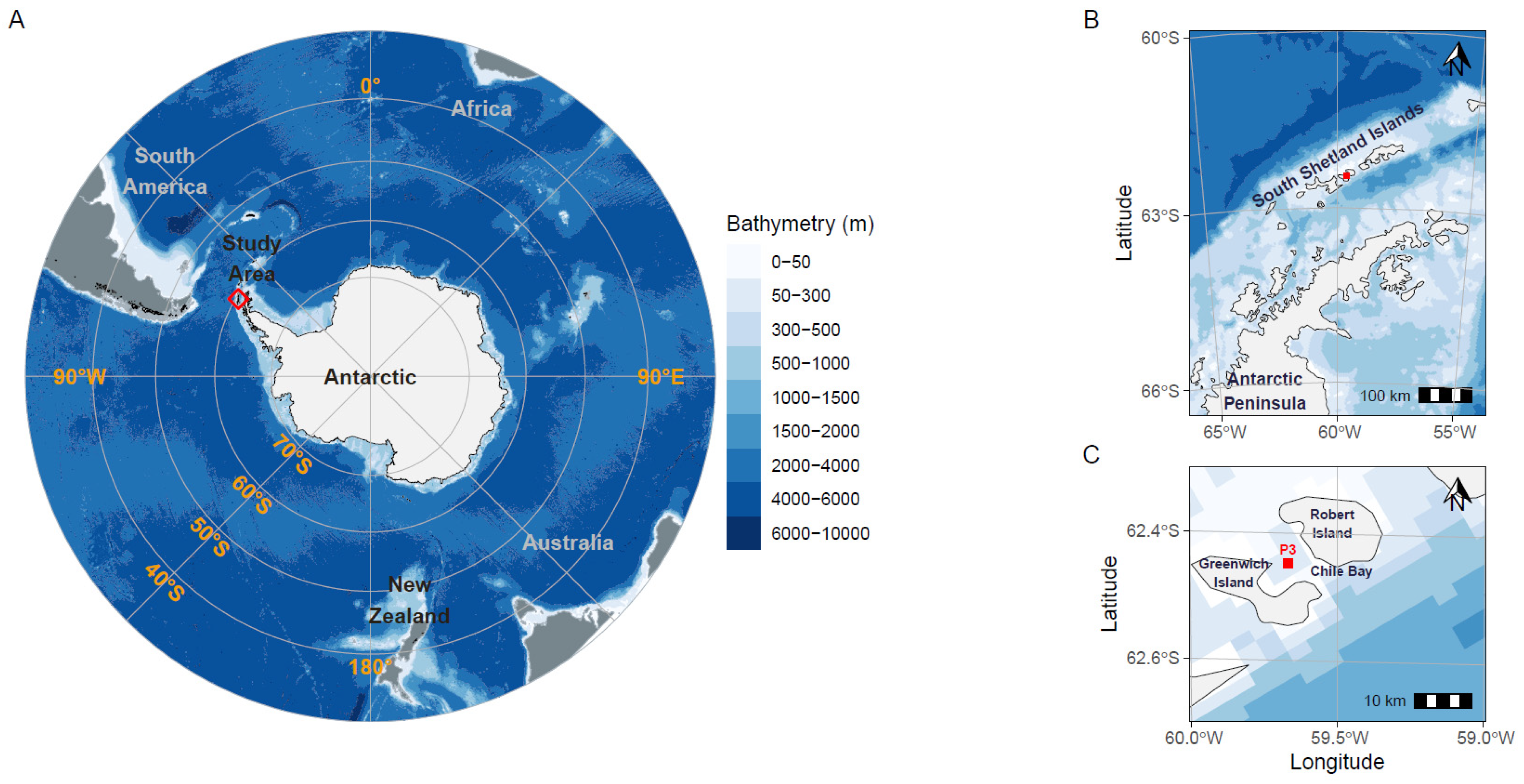
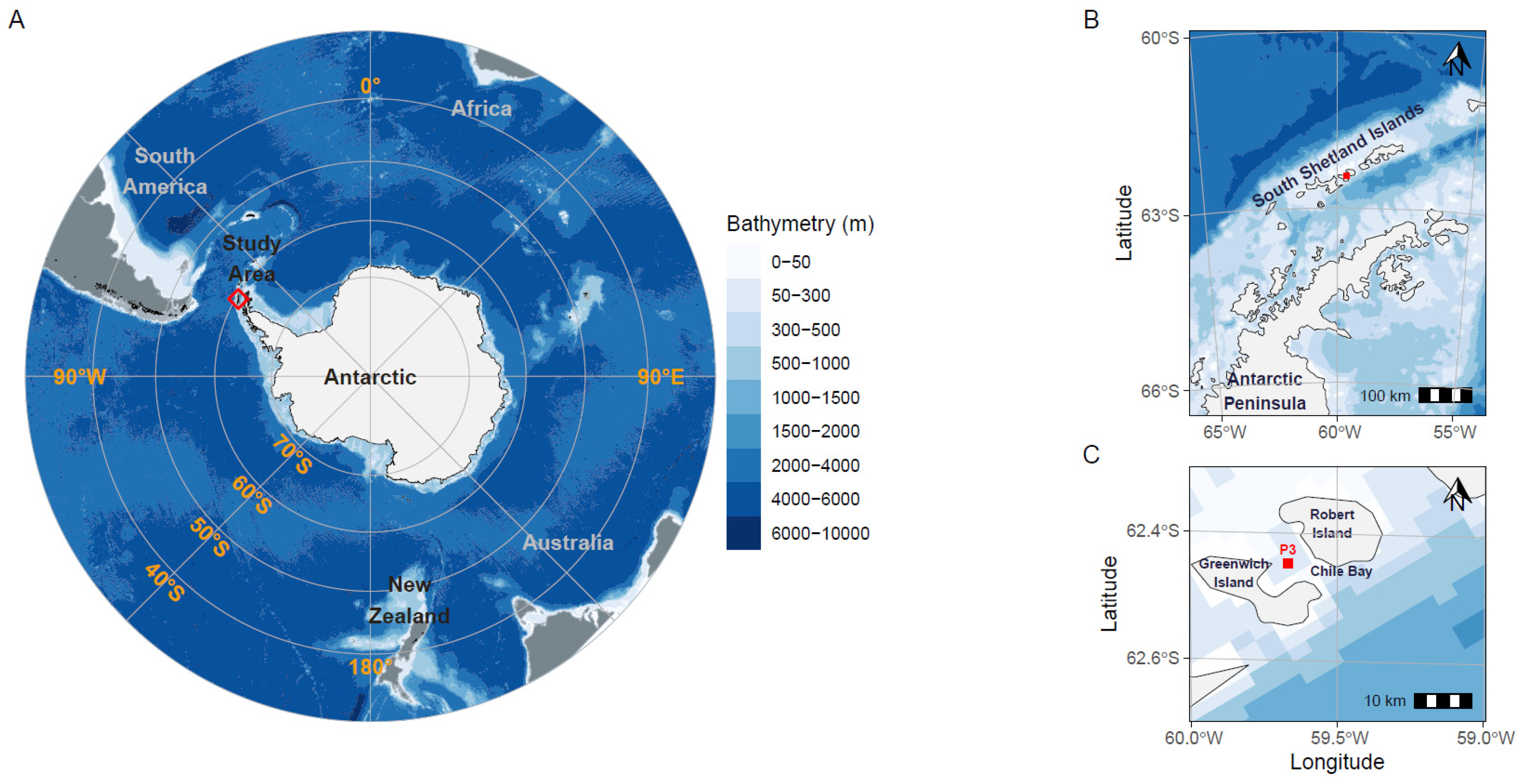
2.1. Sampling Site and Environmental Variable Measurements
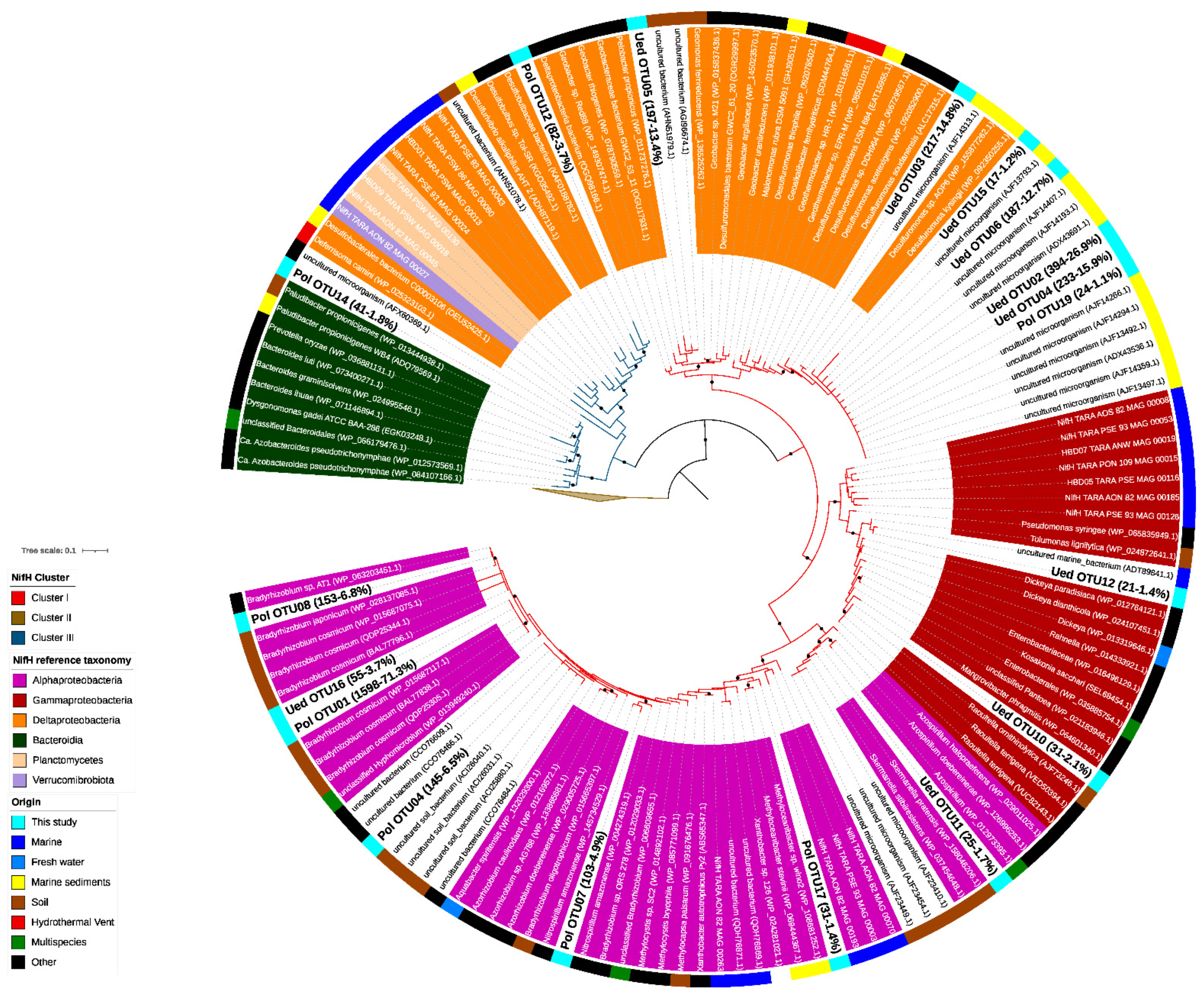
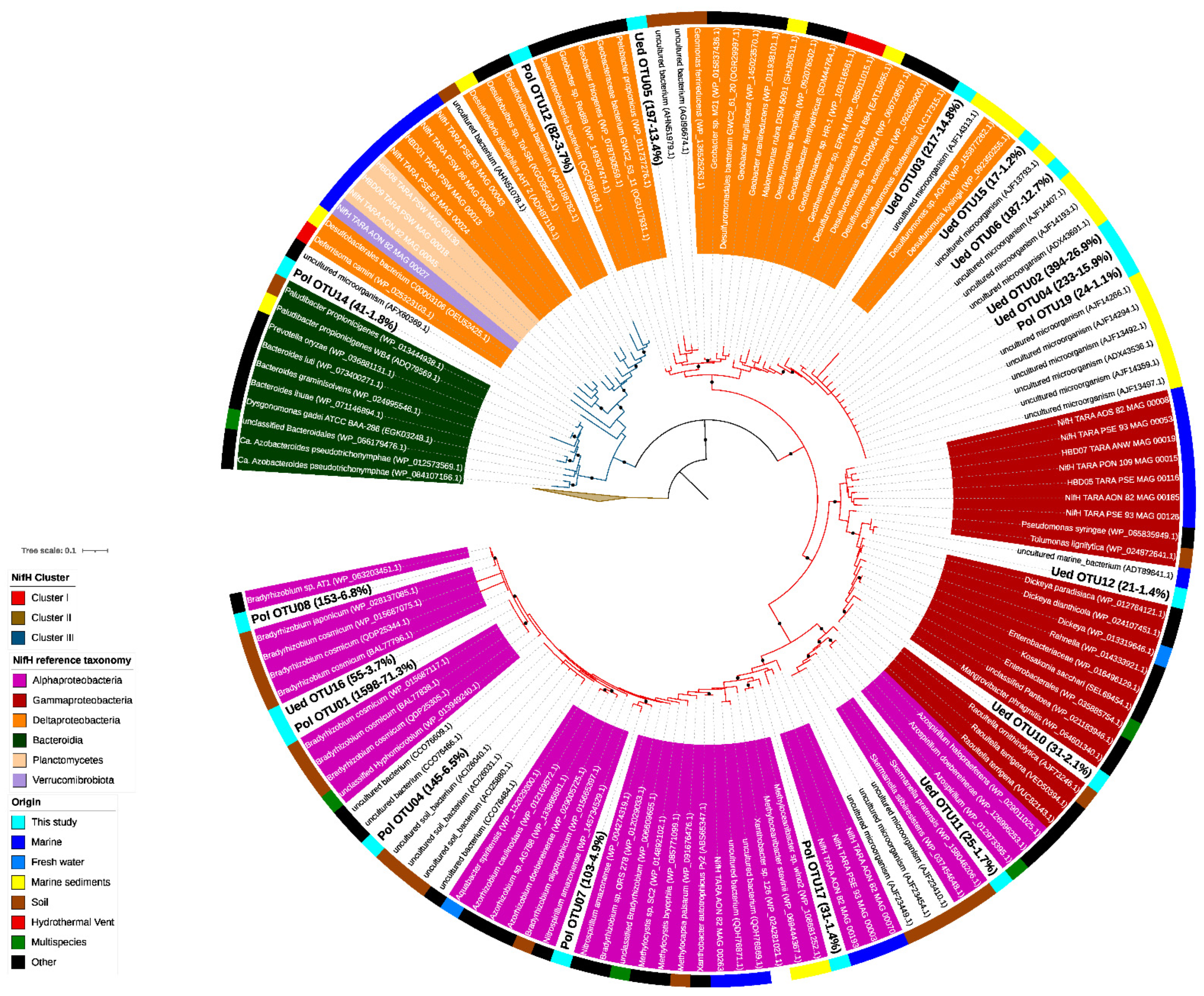
2.2. Phylogenetic Characterization of the Diazotrophic Community
2.3. Nitrogen Fixation Assays
2.4. N2 Fixation Contribution to Coastal Marine Productivity
2.5. Statistical Analysis
3. Results
3.1. Physical and Biogeochemical Variables of Late Summer Seawater in Chile Bay
3.2. Phylogenetic Characterization of the Diazotrophic Community of Chile Bay
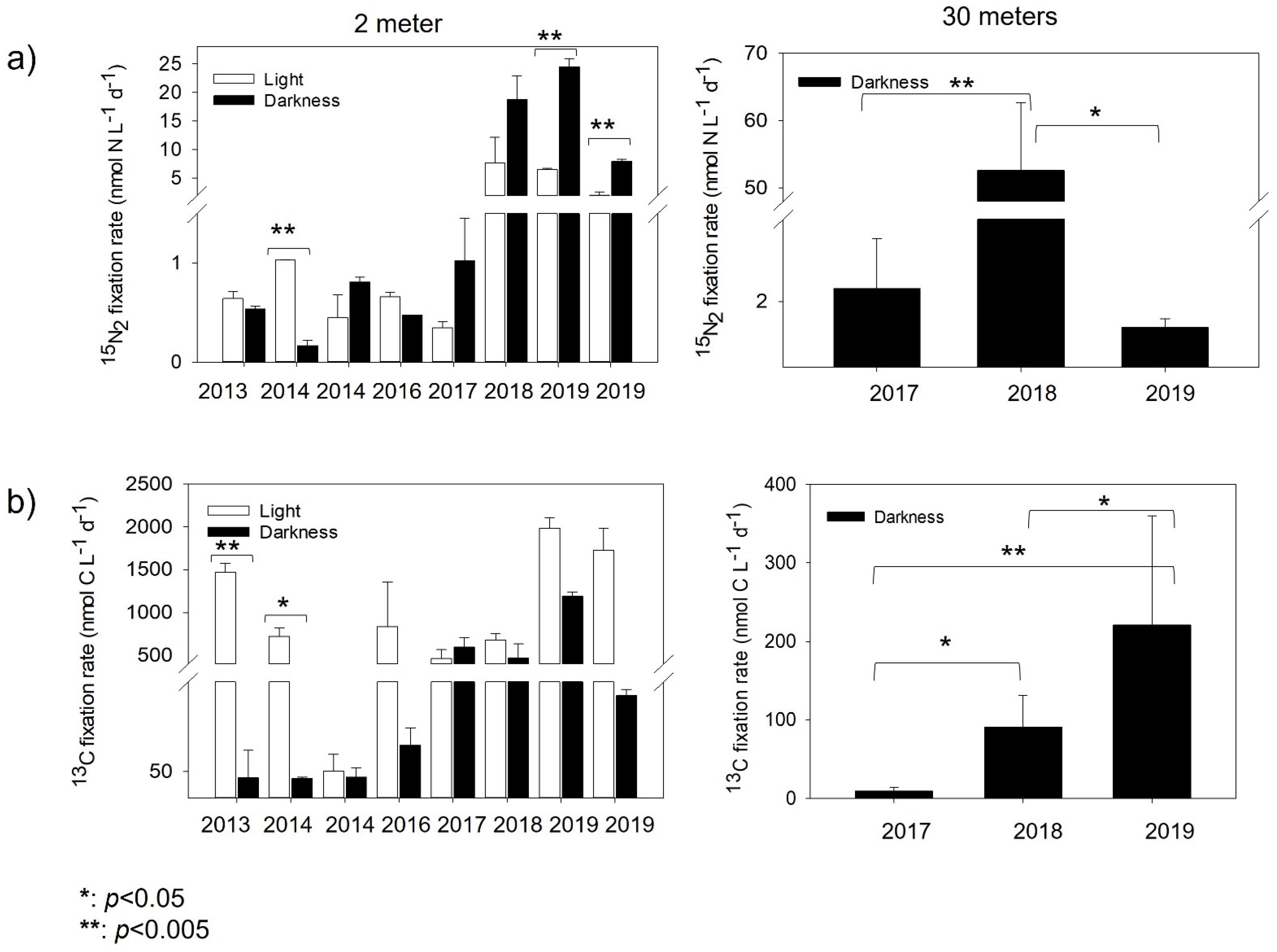
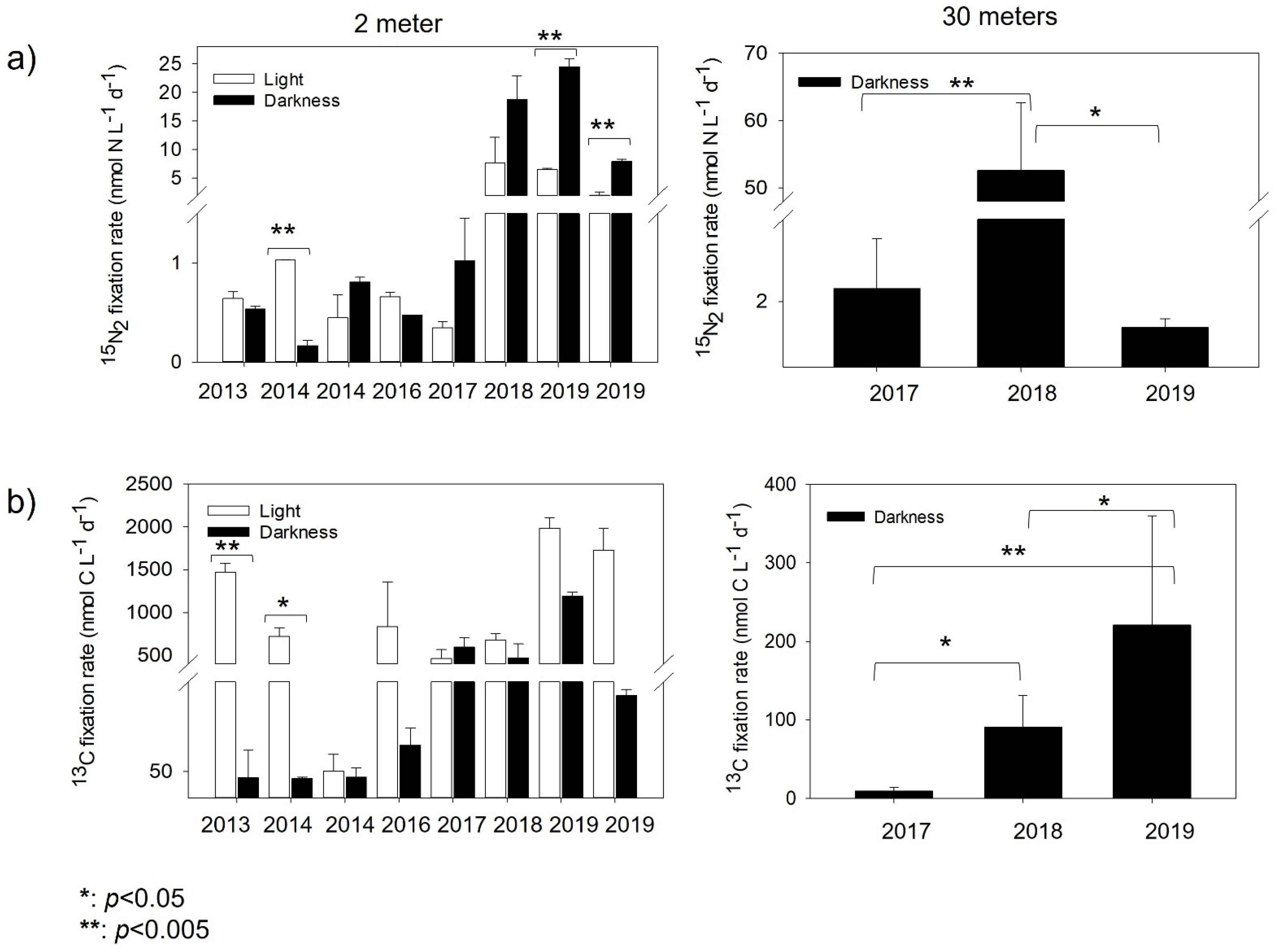
3.3. N2 Fixation and Inorganic Carbon Assimilation Rates
3.4. Contribution of N2 Fixation to New Nitrogen in Chile Bay
4. Discussion
4.1. Diazotrophs in Chile Bay
4.2. N2 Fixation Contribution to New Nitrogen in Chile Bay Seawater
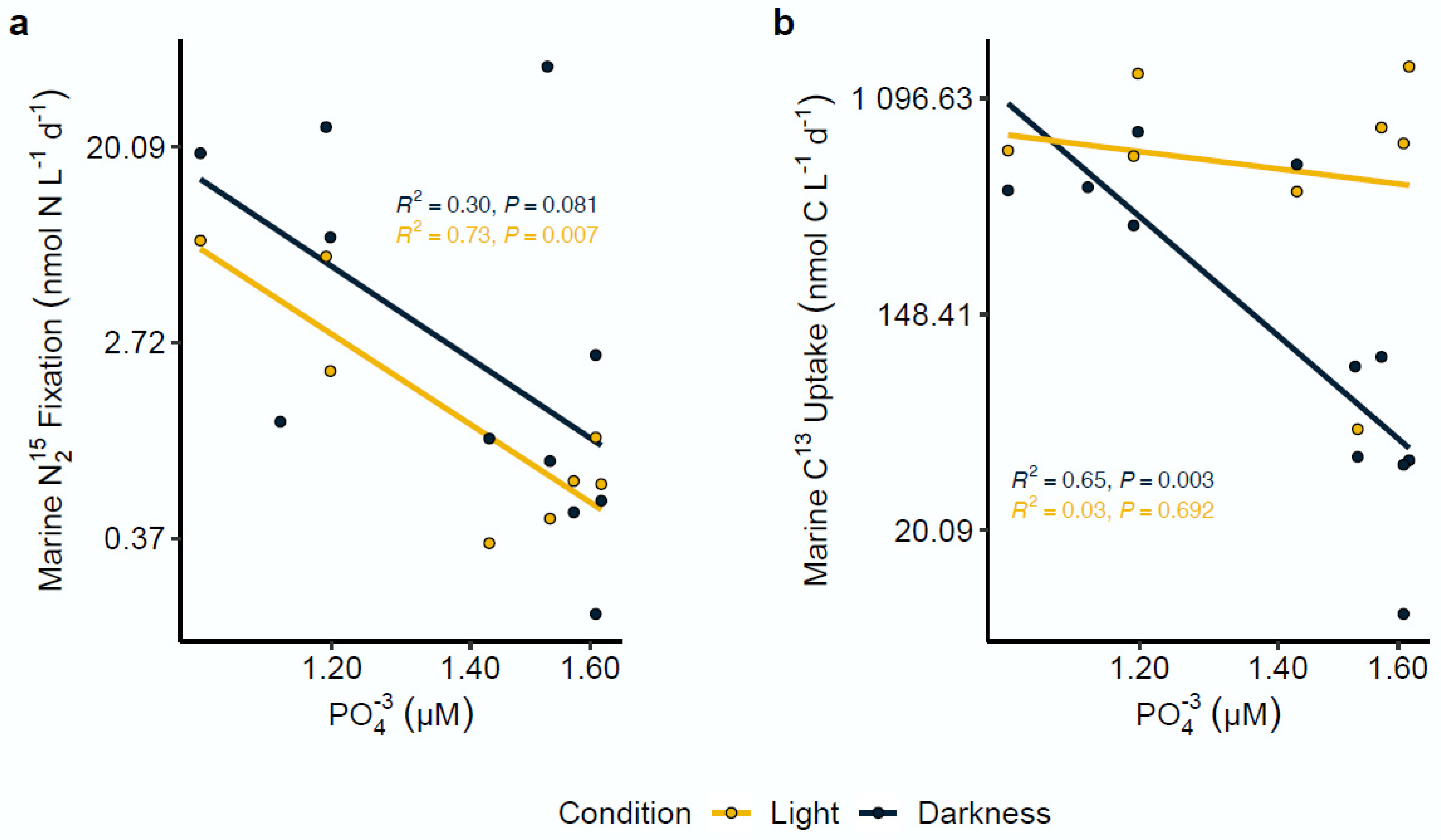
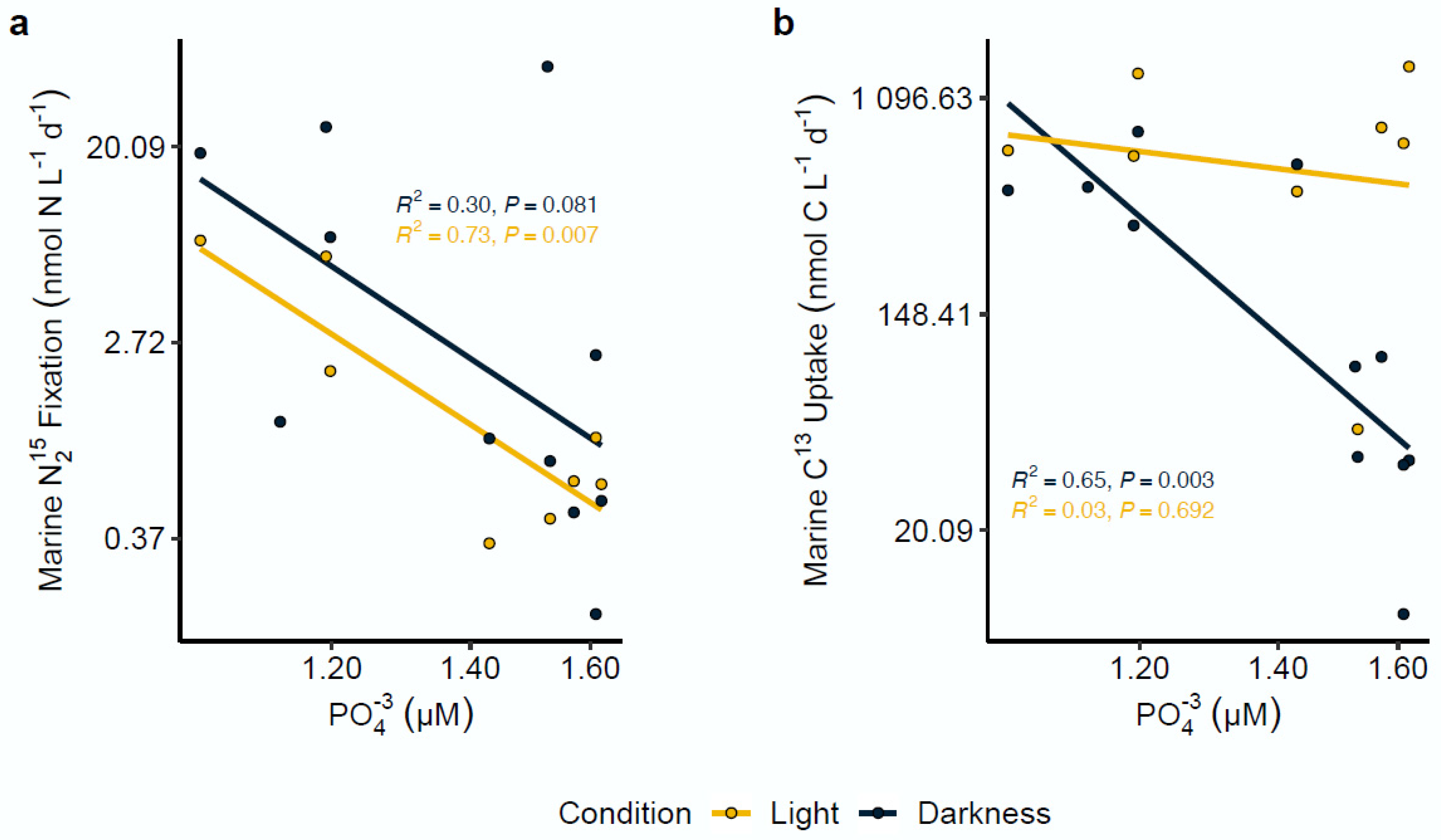
4.3. Effect of Environmental Variables on N2 Fixation Rates
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Levitan, O.; Rosenberg, G.; Setlik, I.; Setlikova, E.; Grigel, J.; Klepetar, J.; Prasil, O.; Berman-Frank, I. Elevated CO2 enhances nitrogen fixation and growth in the marine cyanobacterium Trichodesmium. Glob. Chang. Biol. 2007, 13, 531–538. [Google Scholar] [CrossRef]
- Zehr, J.; Capone, D.G. Changing perspectives in marine nitrogen fixation. Nature 2020, 368, 9514. [Google Scholar] [CrossRef] [PubMed]
- Montoya, J.P.; Holl, C.M.; Zehr, J.P.; Hansen, A.; Villareal, T.A.; Capone, D.G. High rates of N2 fixation by unicellular diazotrophs in the oligotrophic Pacific Ocean. Nature 2004, 430, 1027–1031. [Google Scholar] [CrossRef]
- Rees, A.; Gilbert, J.; Kelly-gerreyn, B. Nitrogen fixation in the western English Channel (NE Atlantic Ocean). Mar. Ecol. Prog. Ser. 2009, 374, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Sohm, J.A.; Webb, E.A.; Capone, D.G. Emerging patterns of marine nitrogen fixation. Nature 2011, 9, 499–508. [Google Scholar] [CrossRef]
- Landolfi, A.; Kähler, P.; Koeve, W.; Oschlies, A. Global marine N2 fixation estimates: From observations to models. Front. Microbiol. 2018, 9, 2112. [Google Scholar] [CrossRef]
- Mulholland, M.R.; Bernhardt, P.W.; Widner, B.N.; Selden, C.R.; Chappell, P.D.; Clayton, S.; Mannino, A.; Hyde, K. High Rates of N2 Fixation in Temperate, Western North Atlantic Coastal Waters Expand the Realm of Marine Diazotrophy. Glob. Biogeochem. Cycles 2019, 33, 826–840. [Google Scholar] [CrossRef] [Green Version]
- Pajares, S.; Ramos, R. Processes and Microorganisms Involved in the Marine Nitrogen Cycle: Knowledge and Gaps. Front. Mar. Sci. 2019, 6, 739. [Google Scholar] [CrossRef]
- Kumar, P.K.; Singh, A.; Ramesh, R.; Nallathambi, T. N2 fixation in the Eastern Arabian Sea: Probable role of heterotrophic diazotrophs. Front. Mar. Sci. 2017, 4, 80. [Google Scholar] [CrossRef] [Green Version]
- Sipler, R.E.; Gong, D.; Baer, S.E.; Sanderson, M.P.; Roberts, Q.N.; Mulholland, M.R.; Bronk, D.A. Preliminary estimates of the contribution of Arctic nitrogen fixation to the global nitrogen budget. Limnol. Oceanogr. Lett. 2017, 2, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Shiozaki, T.; Fujiwara, A.; Inomura, K.; Hirose, Y.; Hashihama, F.; Harada, N. Biological nitrogen fixation detected under Antarctic sea ice. Nat. Geosci. 2020, 13, 729–732. [Google Scholar] [CrossRef]
- Blais, M.; Tremblay, J.É.; Jungblut, A.D.; Gagnon, J.; Martin, J.; Thaler, M.; Lovejoy, C. Nitrogen fixation and identification of potential diazotrophs in the Canadian Arctic. Glob. Biogeochem. Cycles 2012, 26. [Google Scholar] [CrossRef]
- Fernández-Méndez, M.; Turk-Kubo, K.A.; Buttigieg, P.L.; Rapp, J.Z.; Krumpen, T.; Zehr, J.P.; Boetius, A. Diazotroph diversity in the sea ice, melt ponds, and surface waters of the eurasian basin of the Central Arctic Ocean. Front. Microbiol. 2016, 7, 1884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruber, N. Elusive marine nitrogen fixation. Proc. Natl. Acad. Sci. USA 2016, 113, 4246–4248. [Google Scholar] [CrossRef] [Green Version]
- Knapp, A.N. The sensitivity of marine N2 fixation to dissolved inorganic nitrogen. Front. Microbiol. 2012, 3, 374. [Google Scholar] [CrossRef] [Green Version]
- Turk-Kubo, K.A.; Connell, P.; Caron, D.; Hogan, M.E.; Farnelid, H.M.; Zehr, J.P. In situ diazotroph population dynamics under different resource ratios in the North Pacific subtropical gyre. Front. Microbiol. 2018, 9, 1616. [Google Scholar] [CrossRef] [PubMed]
- Karl, D.; Letelier, R.; Tupas, L.; Dore, J.; Christian, J.; Hebel, D. The role of nitrogen fixation in biogeochemical cycling in the subtropical North Pacific Ocean. Nature 1997, 388, 533–538. [Google Scholar] [CrossRef]
- Zehr, J.P.; Kudela, R.M. Nitrogen cycle of the open ocean: From genes to ecosystems. Ann. Rev. Mar. Sci. 2011, 3, 197–225. [Google Scholar] [CrossRef] [Green Version]
- Zehr, J.P.; Ward, B.B. Nitrogen cycling in the ocean: New perspectives on processes and paradigms. Appl. Environ. Microbiol. 2002, 68, 1015–1024. [Google Scholar] [CrossRef] [Green Version]
- Langlois, R.; Hummer, D.; LaRoche, J. Abundances and Distributions of the Dominant nifH Phylotypes in the Northern Atlantic Ocean. Appl. Environ. Microbiol. 2008, 74, 1922–1931. [Google Scholar] [CrossRef] [Green Version]
- Moisander, P.; Moisander, P.; Beinart, R.; Hewson, I.; White, A.; Johnson, K.; Carlson, C.; Montoya, J.; Zehr, J. Broaden the Oceanic N2 Fixation Domain. Science 2010, 327, 1512–1514. [Google Scholar] [CrossRef] [PubMed]
- Farnelid, H.; Andersson, A.; Bertilsson, S.; Al-soud, W.; Hansen, L. Nitrogenase Gene Amplicons from Global Marine Surface Waters Are Dominated by Genes of Non-Cyanobacteria. PLoS ONE 2011, 6, e19223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halm, H.; Lam, P.; Ferdelman, T.G.; Lavik, G.; Dittmar, T.; Laroche, J.; D’Hondt, S.; Kuypers, M.M.M. Heterotrophic organisms dominate nitrogen fixation in the south pacific gyre. ISME J. 2011, 6, 1238–1249. [Google Scholar] [CrossRef] [PubMed]
- Riemann, L.; Farnelid, H.; Steward, G. Nitrogenase genes in non-cyanobacterial plankton: Prevalence, diversity and regulation in marine waters. Aquat. Microb. Ecol. 2010, 61, 235–247. [Google Scholar] [CrossRef] [Green Version]
- Shiozaki, T.; Nagata, T.; Ijichi, M.; Furuya, K. Seasonal dynamics of nitrogen fixation and the diazotroph community in the temperate coastal region of the northwestern North Pacific. Biogeosciences Discuss. 2015, 12, 865–889. [Google Scholar] [CrossRef]
- Bombar, D.; Paerl, R.W.; Riemann, L. Marine Non-Cyanobacterial Diazotrophs: Moving beyond Molecular Detection. Trends Microbiol. 2016, 24, 916–927. [Google Scholar] [CrossRef]
- Gradoville, M.; Bombar, D.; Crump, B.; Letelier, R.; Zehr, J.; White, A. Diversity and activity of nitrogen-fixing communities across ocean basins. Limnol. Oceanogr. 2017, 62, 1895–1909. [Google Scholar] [CrossRef] [Green Version]
- Delmont, T.; Pierella, J.; Veseli, I.; Fuessel, J.; Eren, M.; Foster, R.; Bowler, C.; Wincker, P.; Pelletier, E. Heterotrophic bacterial diazotrophs outperform their cyanobacterial counterparts in metagenomes covering most of the sunlit ocean. ISME J. 2022, 16, 927–936. [Google Scholar] [CrossRef]
- Bowman, J.; Berthiaume, C.; Armbrust, E.; Deming, J. The genetic potential for key biogeochemical processes in Arctic frost flowers and young sea ice revealed by metagenomic analysis. FEMS Microbiol. Ecol. 2014, 89, 376–387. [Google Scholar] [CrossRef]
- Díez, B.; Bergman, B.; Pedrós-Alió, C.; Antó, M.; Snoeijs, P. High cyanobacterial nifH gene diversity in Arctic seawater and sea ice brine. Environ. Microbiol. Rep. 2012, 4, 360–366. [Google Scholar] [CrossRef]
- von Friesen, L.W.; Riemann, L. Nitrogen Fixation in a Changing Arctic Ocean: An Overlooked Source of Nitrogen? Front. Microbiol. 2020, 11, 3149. [Google Scholar] [CrossRef] [PubMed]
- Raes, E.J.; van de Kamp, J.; Bodrossy, L.; Fong, A.A.; Riekenberg, J.; Holmes, B.H.; Erler, D.V.; Eyre, B.D.; Weil, S.S.; Waite, A.M. N2 Fixation and New Insights Into Nitrification From the Ice-Edge to the Equator in the South Pacific Ocean. Front. Mar. Sci. 2020, 7, 389. [Google Scholar] [CrossRef]
- González, M.L.; Molina, V.; Florez-Leiva, L.; Oriol, L.; Cavagna, A.J.; Dehairs, F.; Farias, L.; Fernandez, C. Nitrogen fixation in the Southern Ocean: A case of study of the Fe-fertilized Kerguelen region (KEOPS II cruise). Biogeosciences Discuss. 2014, 11, 17151–17185. [Google Scholar] [CrossRef]
- Wang, W.L.; Moore, J.K.; Martiny, A.C.; Primeau, F.W. Convergent estimates of marine nitrogen fixation. Nature 2019, 566, 205–211. [Google Scholar] [CrossRef]
- Chorus, I.; Welker, M. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management, 2nd ed.; Taylor & Francis: Abingdon, UK, 2021; Available online: https://library.oapen.org/handle/20.500.12657/47047 (accessed on 1 July 2021).
- Strickland, J.D.H.; Parsons, T.R. A Practical Hand Book of Seawater Analysis, 2nd ed.; Fisheries Research Board of Canada Bulletin 157; Minister of Supply and Services Canada: Ottawa, ON, Canada, 1972; Available online: https://www.scirp.org/(S(lz5mqp453edsnp55rrgjct55))/reference/ReferencesPapers.aspx?ReferenceID=1916578 (accessed on 10 February 2021).
- Caspers, H. Internationale Revue der gesamten Hydrobiologie und Hydrographie. Int. Rev. Hydrobiol. 1985, 70, 302–303. Available online: http://doi.wiley.com/10.1002/iroh.19850700232 (accessed on 10 February 2021).
- Holmes, R.; Amonit, A.; Kerouel, R.; Hooker, B.; Peterson, B. A simple and precise method for measuring ammonium in marine and freshwater ecosystems. J. Fischeries Aquat. Sci. 1999, 56, 1801–1808. [Google Scholar] [CrossRef]
- Alcamán-Arias, M.E.; Farías, L.; Verdugo, J.; Alarcón-Schumacher, T.; Díez, B. Microbial activity during a coastal phytoplankton bloom on the Western Antarctic Peninsula in late summer. FEMS Microbiol. Lett. 2018, 365, fny090. [Google Scholar] [CrossRef]
- Poly, F.; Monrozier, L.; Bally, R. Improvement in the RFLP procedure for studying the diversity of nifH genes in communities of nitrogen fixers in soil. Res. Microbiol. 2001, 152, 95–103. [Google Scholar] [CrossRef]
- Angel, R.; Nepel, M.; Panhölzl, C.; Schmidt, H.; Herbold, C.W.; Eichorst, S.A.; Woebken, D. Evaluation of primers targeting the diazotroph functional gene and development of NifMAP—A bioinformatics pipeline for analyzing nifH amplicon data. Front. Microbiol. 2018, 9, 703. [Google Scholar] [CrossRef] [Green Version]
- Schmieder, R.; Edwards, R. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011, 27, 863–864. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyatt, D.; Chen, G.L.; LoCascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. Available online: https://academic.oup.com/mbe/article/30/4/772/1073398 (accessed on 10 February 2021). [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. bioRxiv 2017, 35, 518–522. [Google Scholar] [CrossRef]
- Barbera, P.; Kozlov, A.M.; Czech, L.; Morel, B.; Darriba, D.; Flouri, T.; Stamatakis, A. EPA-ng: Massively Parallel Evolutionary Placement of Genetic Sequences. Syst. Biol. 2018, 68, 365–369. [Google Scholar] [CrossRef]
- Czech, L.; Barbera, P.; Stamatakis, A. Genesis and Gappa: Processing, analyzing and visualizing phylogenetic (placement) data. Bioinformatics 2020, 36, 3263–3265. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, 256–259. [Google Scholar] [CrossRef] [Green Version]
- Alcamán-Arias, M.E.; Cifuentes-Anticevic, J.; Díez, B.; Testa, G.; Troncoso, M.; Bello, E.; Farías, L. Surface Ammonia-Oxidizer Abundance During the Late Summer in the West Antarctic Coastal System. Front. Microbiol. 2022, 13, 821902. [Google Scholar] [CrossRef]
- Dabundo, R.; Lehmann, M.F.; Treibergs, L.; Tobias, C.R.; Altabet, M.A.; Moisander, P.H.; Granger, J. The contamination of commercial 15N2 gas stocks with 15N-labeled nitrate and ammonium and consequences for nitrogen fixation measurements. PLoS ONE 2014, 9, e110335. [Google Scholar] [CrossRef] [PubMed]
- White, A.E.; Granger, J.; Selden, C.; Gradoville, M.R.; Potts, L.; Bourbonnais, A.; Fulweiler, R.W.; Knapp, A.N.; Mohr, W.; Moisander, P.H.; et al. A critical review of the 15N2 tracer method to measure diazotrophic production in pelagic ecosystems. Limnol. Oceanogr. Methods 2020, 18, 129–147. [Google Scholar] [CrossRef] [Green Version]
- Böttjer, D.; Dore, J.E.; Karl, D.M.; Letelier, R.M.; Mahaffey, C.; Wilson, S.T.; Zehr, J.; Church, M.J. Temporal variability of nitrogen fixation and particulate nitrogen export at Station ALOHA. Limnol. Oceanogr. 2017, 62, 200–216. [Google Scholar] [CrossRef] [Green Version]
- Hauri, C.; Doney, S.; Takahashi, T.; Erickson, M.; Jiang, G.; Ducklow, H. Two decades of inorganic carbon dynamics along the West Antarctic Peninsula. Biogeosciences 2015, 12, 6761–6779. [Google Scholar] [CrossRef] [Green Version]
- Montoya, J.; Voss, M.; Kahler, P.; Capone, D. A Simple, High-Precision, High-Sensitivity Tracer Assay for N ( inf2) Fixation. These include: A Simple, High-Precision, High-Sensitivity Tracer Assay for N2 Fixation. Appl. Environ. Microbiol. 1996, 62, 986–993. [Google Scholar] [CrossRef] [Green Version]
- Raimbault, P.; Garcia, N. Evidence for efficient regenerated production and dinitrogen fixation in nitrogen-deficient waters of the South Pacific Ocean: Impact on new and export production estimates. Biogeosciences 2008, 5, 323–338. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Wagner, H. Vegan: 2.5-5., Community Ecology Package; R Package Version; R Core Team: Vienna, Austria, 2019. [Google Scholar]
- Siebeneichler, S.C.; Barbosa, J.S.; Cruz, A.M.M.; Ramos, M.A.D.; Fernandes, H.E.; Nascimento, V.L. Comparison between extraction methods of photosynthetic pigments in Acacia mangium. Commun. Plant Sci. 2019, 9, 2237–4027. [Google Scholar] [CrossRef] [Green Version]
- Redfield, A.C. On the Proportions of Organic Derivatives in Sea Water and Their Relation to the Composition of Plankton; University Press of Liverpool: Liverpool, UK, 1934; pp. 176–192. [Google Scholar]
- Codispoti, L.A. An oceanic fixed nitrogen sink exceeding 400 Tg N a−1 vs the concept of homeostasis in the fixed-nitrogen inventory. Biogeosciences 2007, 4, 233–253. [Google Scholar] [CrossRef] [Green Version]
- Eugster, O.; Gruber, N.; Deutsch, C.; Jaccard, S.L.; Payne, M.R. The dynamics of the marine nitrogen cycle across the last deglaciation. Paleoceanography 2013, 28, 116–129. [Google Scholar] [CrossRef] [Green Version]
- Mohr, W.; Großkopf, T.; Wallace, D.W.R.; LaRoche, J. Methodological underestimation of oceanic nitrogen fixation rates. PLoS ONE 2010, 5, e12583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grokopf, T.; Mohr, W.; Baustian, T.; Schunck, H.; Gill, D.; Kuypers, M.M.M.; Lavik, G.; Schmitz, R.A.; Wallace, D.W.R.; Laroche, J. Doubling of marine dinitrogen-fixation rates based on direct measurements. Nature 2012, 488, 361–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biggs, T.E.G.; Alvarez-Fernandez, S.; Evans, C.; Mojica, K.D.A.; Rozema, P.D.; Venables, H.J.; Pond, D.W.; Brussaard, C.P.D. Antarctic phytoplankton community composition and size structure: Importance of ice type and temperature as regulatory factors. Polar Biol. 2019, 42, 1997–2015. [Google Scholar] [CrossRef] [Green Version]
- Höfer, J.; Giesecke, R.; Hopwood, M.J.; Carrera, V.; Alarcón, E.; González, H.E. The role of water column stability and wind mixing in the production/export dynamics of two bays in the Western Antarctic Peninsula. Prog. Oceanogr. 2019, 174, 105–116. [Google Scholar] [CrossRef]
- Alcamán-Arias, M.E.; Fuentes-Alburquenque, S.; Vergara-Barros, P.; Cifuentes-Anticevic, J.; Verdugo, J.; Polz, M.; Farías, L.; Pedrós-Alió, C.; Díez, B. Coastal Bacterial Community Response to Glacier Melting in the Western Antarctic Peninsula. Microorganisms 2021, 9, 88. [Google Scholar] [CrossRef]
- Fuentes, S.; Arroyo, J.I.; Rodríguez-Marconi, S.; Masotti, I.; Alarcón-Schumacher, T.; Polz, M.F.; Trefault, N.; De la Iglesia, R.; Díez, B. Summer phyto- and bacterioplankton communities during low and high productivity scenarios in the Western Antarctic Peninsula. Polar Biol. 2019, 42, 159–169. [Google Scholar] [CrossRef]
- Alarcón-Schumacher, T.; Guajardo-Leiva, S.; Antón, J.; Díez, B. Elucidating Viral Communities During a Phytoplankton Bloom on the West Antarctic Peninsula. Front. Mar. Sci. 2019, 10, 1014. [Google Scholar] [CrossRef]
- Farnelid, H.; Turk-Kubo, K.; Ploug, H.; Ossolinski, J.E.; Collins, J.R.; Van Mooy, B.A.S.; Zehr, J.P. Diverse diazotrophs are present on sinking particles in the North Pacific Subtropical Gyre. ISME J. 2019, 13, 170–182. [Google Scholar] [CrossRef]
- Riemann, L.; Rahav, E.; Passow, U.; Grossart, H.-P.; de Beer, D.; Klawonn, I.; Eichner, M.; Benavides, M.; Bar-Zeev, E. Planktonic Aggregates as Hotspots for Heterotrophic Diazotrophy: The Plot Thickens. Front. Microbiol. 2022, 13, 1092. [Google Scholar] [CrossRef]
- Bonnet, S.; Dekaezemacker, J.; Turk-Kubo, K.A.; Moutin, T.; Hamersley, R.M.; Grosso, O.; Zehr, J.P.; Capone, D.G. Aphotic N2 fixation in the eastern tropical South Pacific Ocean. PLoS ONE 2013, 8, e81265. [Google Scholar] [CrossRef]
- Rahav, E.; Bar-Zeev, E.; Ohayon, S.; Elifantz, H.; Belkin, N.; Herut, B.; Mulholland, M.R.; Berman-Frank, I.; Goodman, E. Dinitrogen fixation in aphotic oxygenated marine environments. Front. Microbiol. 2013, 4, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, C.; Farías, L.; Ulloa, O. Nitrogen fixation in denitrified marine waters. PLoS ONE 2011, 6, e20539. [Google Scholar] [CrossRef] [PubMed]
- Jayakumar, A.; Al-Rshaidat, M.M.D.; Ward, B.B.; Mulholland, M.R. Diversity, distribution, and expression of diazotroph nifH genes in oxygen-deficient waters of the Arabian Sea. FEMS Microbiol. Ecol. 2012, 82, 597–606. [Google Scholar] [CrossRef] [Green Version]
- Loescher, C.R.; Grokopf, T.; Desai, F.D.; Gill, D.; Schunck, H.; Croot, P.L.; Schlosser, C.; Neulinger, S.C.; Pinnow, N.; Lavik, G.; et al. Facets of diazotrophy in the oxygen minimum zone waters off Peru. ISME J. 2014, 8, 2180–2192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayakumar, A.; Ward, B.B. Diversity and distribution of nitrogen fixation genes in the oxygen minimum zones of the world oceans. Biogeosciences 2020, 17, 5953–5966. [Google Scholar] [CrossRef]
- Farnelid, H.; Bentzon-Tilia, M.; Andersson, A.F.; Bertilsson, S.; Nter Jost, G.; Labrenz, M.; Rgens, K.J.; Riemann, L. Active nitrogen-fixing heterotrophic bacteria at and below the chemocline of the central Baltic Sea. ISME J. 2013, 7, 1413–1423. [Google Scholar] [CrossRef]
- Benavides, M.; Bonnet, S.; Berman-Frank, I.; Riemann, L. Deep into oceanic N2 fixation. Front. Mar. Sci. 2018, 5, 3–6. [Google Scholar] [CrossRef] [Green Version]
- Bentzon-Tilia, M.; Traving, S.J.; Mantikci, M.; Knudsen-Leerbeck, H.; Hansen, J.L.; Markager, S.; Riemann, L. Significant N2 fixation by heterotrophs, photoheterotrophs and heterocystous cyanobacteria in two temperate estuaries. ISME J. 2015, 9, 273–285. [Google Scholar] [CrossRef] [Green Version]
- Severin, I.; Bentzon-Tilia, M.; Moisander, P.H.; Riemann, L. Nitrogenase expression in estuarine bacterioplankton influenced by organic carbon and availability of oxygen. FEMS Microbiol. Lett. 2015, 362, 105. [Google Scholar] [CrossRef]
- Moisander, P.H.; Benavides, M.; Bonnet, S.; Berman-Frank, I.; White, A.E.; Riemann, L. Chasing after Non-cyanobacterial Nitrogen Fixation in Marine Pelagic Environments. Front. Microbiol. 2017, 8, 1736. [Google Scholar] [CrossRef]
- Chakraborty, S.; Andersen, K.H.; Visser, A.W.; Inomura, K.; Follows, M.J.; Riemann, L. Quantifying nitrogen fixation by heterotrophic bacteria in sinking marine particles. Nat. Commun. 2021, 12, 4085. [Google Scholar] [CrossRef] [PubMed]
- Fulweiler, R.W.; Nixon, S.W.; Buckley, B.A.; Granger, S.L. Reversal of the net dinitrogen gas flux in coastal marine sediments. Nature 2007, 448, 180–182. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Lin, W.; Shen, R.; Hong, H.; Kao, S.J.; Shi, D. Nitrogen fixation in two coastal upwelling regions of the Taiwan Strait. Sci. Rep. 2017, 7, 17601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, W.; Wang, S.; Fonseca-Batista, D.; Dehairs, F.; Gifford, S.; Gonzalez, A.G.; Gallinari, M.; Planquette, H.; Sarthou, G.; Cassar, N. Revisiting the distribution of oceanic N2 fixation and estimating diazotrophic contribution to marine production. Nat. Commun. 2019, 10, 831. [Google Scholar] [CrossRef]
- Delmont, T.; Quince, C.; Shaiber, A.; Esen, Ö.C.; Lee, S.T.; Rappé, M.S.; MacLellan, S.L.; Lücker, S.; Eren, A.M. Nitrogen-fixing populations of Planctomycetes and Proteobacteria are abundant in surface ocean metagenomes. Nat. Microbiol. 2018, 3, 804–813. [Google Scholar] [CrossRef] [Green Version]
- Capone, D.G.; Subramaniam, A.; Montoya, J.P.; Voss, M.; Humborg, C.; Johansen, A.M.; Siefert, R.L.; Carpenter, E.J. An extensive bloom of the N2-fixing Cyanobacterium trichodesmium erythraeum in the central Arabian Sea. Mar. Ecol. Prog. Ser. 1998, 172, 281–292. [Google Scholar] [CrossRef] [Green Version]
- Zehr, J.; Waterburry, J.; Turner, P.; Montoya, J.; Omeregie, E.; Steward, G.; Hansen, A.; Karl, D.M. Unicellular cyanobacteria fix N2 in the subtropical North Pacic Ocean. Nature 2001, 715, 25–28. [Google Scholar]
- Wu, C.; Fu, F.X.; Sun, J.; Thangaraj, S.; Pujari, L. Nitrogen Fixation by Trichodesmium and unicellular diazotrophs in the northern South China Sea and the Kuroshio in summer. Sci. Rep. 2018, 8, 2415. [Google Scholar] [CrossRef] [Green Version]
- Herbert, R.A. Heterotrophic nitrogen fixation in shallow estuarine sediments. J. Exp. Mar. Bio. Ecol. 1975, 18, 215–225. [Google Scholar] [CrossRef]
- Nedwell, D.B.; Azni bin Abdul Aziz, S. Heterotrophic nitrogen fixation in an intertidal saltmarsh sediment. Estuar. Coast. Mar. Sci. 1980, 10, 699–702. [Google Scholar] [CrossRef]
- Inomura, K.; Bragg, J.; Follows, M.J. A quantitative analysis of the direct and indirect costs of nitrogen fixation: A model based on Azotobacter vinelandii. ISME J. 2017, 11, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Inomura, K.; Bragg, J.; Riemann, L.; Follows, M.J. A quantitative model of nitrogen fixation in the presence of ammonium. PLoS ONE 2018, 13, e0208282. [Google Scholar] [CrossRef] [PubMed]
- Harding, K.; Turk-kubo, K.A.; Sipler, R.E.; Mills, M.M.; Bronk, D.A. Symbiotic unicellular cyanobacteria fix nitrogen in the Arctic Ocean. Proc. Natl. Acad. Sci. USA 2018, 115, 13371–13375. [Google Scholar] [CrossRef] [Green Version]
- Shiozaki, T.; Inomura, K.; Fujiwara, A.; Hirose, Y.; Hashihama, F.; Harada, N. Reply to: Questioning High Nitrogen Fixation Rate Measurements in the Southern Ocean. Nat. Geosci. 2022, 15, 31–32. [Google Scholar] [CrossRef]
- White, A.E.; Granger, J.; Turk-Kubo, K. Questioning High Nitrogen Fixation Rate Measurements in the Southern Ocean. Nat. Geosci. 2022, 15, 29–30. [Google Scholar] [CrossRef]
- Capone, D.G.; Burns, J.A.; Montoya, J.P.; Subramaniam, A.; Mahaffey, C.; Gunderson, T.; Michaels, A.F.; Carpenter, E.J. Nitrogen fixation by Trichodesmium spp.: An important source of new nitrogen to the tropical and subtropical North Atlantic Ocean. Glob. Biogeochem. Cycles 2005, 19, 1–17. [Google Scholar] [CrossRef]
- Bonnet, S.; Caffin, M.; Berthelot, H.; Moutin, T. Hot spot of N2 fixation in the western tropical South Pacific pleads for a spatial decoupling between N2 fixation and denitrification. Proc. Natl. Acad. Sci. USA 2017, 114, E2800–E2801. [Google Scholar] [CrossRef] [Green Version]
- Holl, C.; Waite, A.; Pesant, S.; Thompson, P.; Montoya, J. Unicellular diazotrophy as a source of nitrogen to Leeuwin Current coastal eddies. Deep. Res. Part II Top. Stud. Oceanogr. 2007, 54, 1045–1054. [Google Scholar] [CrossRef]
- Dekaezemacker, J.; Bonnet, S. Sensitivity of N2 fixation to combined nitrogen forms (NO3− and NH4+) in two strains of the marine diazotroph Crocosphaera watsonii (Cyanobacteria). Mar. Ecol. Prog. Ser. 2011, 438, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Rabouille, S.; Randall, B.; Talec, A.; Raimbault, P.; Blasco, T.; Latifi, A.; Oschlies, A. Independence of a marine unicellular diazotroph to the presence of NO3−. Microorganisms 2021, 9, 2073. [Google Scholar] [CrossRef]
- Weber, T.S.; Deutsch, C. Ocean nutrient ratios governed by plankton biogeography. Nature 2010, 467, 550–554. [Google Scholar] [CrossRef] [PubMed]
- Arrigo, K.R.; Robinson, D.H.; Worthen, D.L.; Dunbar, R.B.; DiTullio, G.R.; VanWoert, M.; Lizotte, M.P. Phytoplankton community structure and the drawdown of nutrients and CO2 in the Southern Ocean. Science 1999, 283, 365–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sañudo-Wilhelmy, S.A.; Kustka, A.B.; Gobler, C.J.; Hutchins, D.A.; Yang, M.; Lwiza, K.; Burns, J.; Capone, D.G.; Raven, J.A.; Carpenter, E.J. Phosphorus limitation of nitrogen fixation by Trichodesmium in the central Atlantic Ocean. Nature 2001, 411, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Inomura, K.; Deutsch, C.; Masuda, T.; Prášil, O.; Follows, M.J. Quantitative models of nitrogen-fixing organisms. Comput. Struct. Biotechnol. J. 2020, 18, 3905–3924. [Google Scholar] [CrossRef]
- Inomura, K.; Deutsch, C.; Wilson, S.T.; Masuda, T.; Lawrenz, E.; Sobotka, R.; Gauglitz, J.M.; Saito, M.A.; Follows, M.J. Quantifying Oxygen Management and Temperature and Light Dependencies of Nitrogen Fixation by Crocosphaera watsonii. mSphere 2019, 4, e00531-19. [Google Scholar] [CrossRef] [Green Version]
- Inomura, K.; Wilson, S.T.; Deutsch, C. Mechanistic Model for the Coexistence of Nitrogen Fixation and Photosynthesis in Marine Trichodesmium. mSystems 2019, 4, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Dedysh, S.N.; Didriksen, A.; Danilova, O.V.; Belova, S.E.; Liebner, S.; Svenning, M.M. Methylocapsa palsarum sp. nov., a methanotroph isolated from a subarctic discontinuous permafrost ecosystem. Int. J. Syst. Evol. Microbiol. 2015, 65, 3618–3624. [Google Scholar] [CrossRef]
- Yasuda, S.; Suenaga, T.; Orschler, L.; Agrawal, S.; Lackner, S.; Terada, A. Identification of a Metagenome-Assembled Genome of an Uncultured Methyloceanibacter sp. Strain Acquired from an Activated Sludge System Used for Landfill Leachate Treatment. Microbiol. Resour. Announc. 2020, 9, e00771-20. [Google Scholar] [CrossRef]
- Vekeman, B.; Kerckhof, F.-M.; Cremers, G.; De Vos, P.; Vandamme, P.; Boon, N.; Op Den Camp, H.J.M.; Heylen, K. New Methyloceanibacter diversity from North Sea sediments includes methanotroph containing solely the soluble methane monooxygenase. Environ. Microbiol. 2016, 18, 4523–4536. [Google Scholar] [CrossRef] [Green Version]
- Han, D.; Dedysh, S.N.; Liesack, W. Unusual Genomic Traits Suggest Methylocystis bryophila S285 to Be Well Adapted for Life in Peatlands. Genome Biol. Evol. 2018, 10, 623–628. [Google Scholar] [CrossRef] [Green Version]
- Dam, B.; Dam, S.; Kube, M.; Reinhardt, R.; Liesack, W. Complete genome sequence of Methylocystis sp. strain SC2, an aerobic methanotroph with high-affinity methane oxidation potential. J. Bacteriol. 2012, 194, 6008–6009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oren, A. The Prokaryotes: Alphaproteobacteria and Betaproteobacteria; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1–1012. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Depth (m) | Temperature °C | Salinity | Chl-a (mg L−1) | NH4+ (µM) | NO2− (µM) | NO3− (µM) | PO43− (µM) | N:P |
|---|---|---|---|---|---|---|---|---|---|
| 8 February 2013 | 2 | 0.55 | 34.15 | 0.03 | NaN | 0.32 | 20.52 | 1.62 | 12.85 |
| 14 February 2014 | 2 | −0.11 | 33.20 | 0.36 | NaN | 0.16 | 19.52 | 1.61 | 12.22 |
| 22 February 2014 | 2 | 0.34 | 33.47 | 1.18 | NaN | 0.13 | 17.16 | 1.53 | 11.30 |
| 22 February 2016 | 2 | −0.25 | NaN | 1.01 | NaN | 0.29 ± 0.03 | 14.56 ± 0.16 | 1.57 ± 0.16 | 9.45 |
| 10 February 2017 | 2 | 2.23 | 34.03 | 0.69 ± 0.32 | 0.33 ± 0.07 | 0.22 ± 0.02 | 21.64 ± 2.12 | 1.43 ± 0.11 | 15.52 |
| 21 February 2017 | 30 | 1.42 | 34.16 | 0.327 ± 0.17 | 0.61 ± 0.09 | 0.204 ± 0.03 | 23.3 ± 4.55 | 1.61 ± 0.13 | 14.98 |
| 9 February 2018 | 30 | 1.33 | 34.05 | 1.55 ± 1.63 | 0.96 ± 0.29 | 0.26 ± 0.01 | 20.90 ± 3.73 | 1.53 ± 0.30 | 14.50 |
| 16 February 2018 | 2 | 1.61 | 34.01 | 3.58 ± 2.07 | 0.93 ± 0.09 | 0.26 ± 0.04 | 17.70 ± 5.50 | 1.04 ± 0.42 | 18.22 |
| 17 February 2019 | 2 | 1.54 | 34.10 | 2.14 ± 1.90 | 0.48 ± 0.18 | 0.17 ± 0.03 | 16.50 ± 3.97 | 1.19 ± 0.27 | 14.39 |
| 8 March 2019 | 2 | 1.11 | 34.36 | 4.17 ± 2.03 | 1.19 ± 0.54 | 0.16 ± 0.04 | 15.55 ± 4.31 | 1.20 ± 0.23 | 14.11 |
| 8 March 2019 | 30 | 1.16 | 34.21 | 2.83 ± 1.81 | 1.27 ± 0.78 | 0.17 ± 0.04 | 17.28 ± 4.06 | 1.13 ± 0.26 | 16.52 |
| 15N2 Contribution to | |||||||
|---|---|---|---|---|---|---|---|
| Date | Sample Depth (m) | Condition | molC:molN | Marine 15N2 Fixation | Total Marine 13C Uptake | Total N2LPP (C) * | Total N2P (N) ** |
| (nmol N L−1 d−1) | (nmol C L−1 d−1) | % | % | ||||
| 8 February 2013 | 2 | Light | 10.49 | 0.64 ± 0.06 | 1471 ± 101.51 | 0.46 | 0.76 |
| 14 February 2014 | 2 | Light | 7.47 | 1.03 ± 0.002 | 723 ± 97.91 | 1.07 | 4.29 |
| 22 February 2014 | 2 | Light | 5.95 | 0.45 ± 0.14 | 51.18 ± 31.50 | 5.23 | 0.66 |
| 22 February 2016 | 2 | Light | 6.80 | 0.66 ± 0.05 | 837 ± 5515.2 | 0.54 | 9.82 |
| 10 February 2017 | 2 | Light | 5.82 | 0.35 ± 0.09 | 463 ± 104 | 0.44 | 0.31 |
| 16 February 2018 | 2 | Light | 5.02 | 7.70 ± 4.42 | 677 ± 76.3 | 5.71 | 3.84 |
| 17 February 2019 | 2 | Light | 4.11 | 6.54 ± 0.20 | 643 | 4.18 | 4.35 |
| 8 March 2019 | 2 | Light | 3.11 | 2.03 ± 0.57 | 1380 | 0.46 | 1.39 |
| 8 February 2013 | 2 | Darkness | 0.18 | 0.54 ± 0.03 | 38.37 ± 22.04 | 0.25 | 4.66 |
| 14 February 2014 | 2 | Darkness | 7.65 | 0.17 ± 0.05 | 36.80 ± 2.93 | 3.50 | 0.60 |
| 22 February 2014 | 2 | Darkness | 6.43 | 0.81 ± 0.05 | 39.60 ± 17.53 | 13.15 | 0.82 |
| 22 February 2016 | 2 | Darkness | 6.31 | 0.48 | 100 ± 32.51 | 3.00 | 37.59 |
| 10 February 2017 | 2 | Darkness | 4.72 | 1.02 ± 0.73 | 595 ± 109 | 0.81 | 2.26 |
| 16 February 2018 | 2 | Darkness | 4.22 | 18.78 ± 2.19 | 468 ± 165 | 16.93 | 5.76 |
| 17 February 2019 | 2 | Darkness | 4.69 | 24.51 ± 1.40 | 338 ± 167 | 34.01 | 36.96 |
| 8 March 2019 | 2 | Darkness | 3.29 | 7.96 ± 0.38 | 805 ± 345 | 3.25 | 15.99 |
| 21 February 2017 | 30 | Darkness | 4.96 | 2.39 ± 1.53 | 9.24 ± 4.70 | 128 | 5.13 |
| 9 February 2018 | 30 | Darkness | 7.10 | 45.40 ± 10.09 | 91.47 ± 40.09 | 354 | 12.88 |
| 8 March 2019 | 30 | Darkness | 3.55 | 1.21 ± 0.27 | 482 ± 123 | 0.89 | 2.80 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alcamán-Arias, M.E.; Cifuentes-Anticevic, J.; Castillo-Inaipil, W.; Farías, L.; Sanhueza, C.; Fernández-Gómez, B.; Verdugo, J.; Abarzua, L.; Ridley, C.; Tamayo-Leiva, J.; et al. Dark Diazotrophy during the Late Summer in Surface Waters of Chile Bay, West Antarctic Peninsula. Microorganisms 2022, 10, 1140. https://doi.org/10.3390/microorganisms10061140
Alcamán-Arias ME, Cifuentes-Anticevic J, Castillo-Inaipil W, Farías L, Sanhueza C, Fernández-Gómez B, Verdugo J, Abarzua L, Ridley C, Tamayo-Leiva J, et al. Dark Diazotrophy during the Late Summer in Surface Waters of Chile Bay, West Antarctic Peninsula. Microorganisms. 2022; 10(6):1140. https://doi.org/10.3390/microorganisms10061140
Chicago/Turabian StyleAlcamán-Arias, María E., Jerónimo Cifuentes-Anticevic, Wilson Castillo-Inaipil, Laura Farías, Cynthia Sanhueza, Beatriz Fernández-Gómez, Josefa Verdugo, Leslie Abarzua, Christina Ridley, Javier Tamayo-Leiva, and et al. 2022. "Dark Diazotrophy during the Late Summer in Surface Waters of Chile Bay, West Antarctic Peninsula" Microorganisms 10, no. 6: 1140. https://doi.org/10.3390/microorganisms10061140
APA StyleAlcamán-Arias, M. E., Cifuentes-Anticevic, J., Castillo-Inaipil, W., Farías, L., Sanhueza, C., Fernández-Gómez, B., Verdugo, J., Abarzua, L., Ridley, C., Tamayo-Leiva, J., & Díez, B. (2022). Dark Diazotrophy during the Late Summer in Surface Waters of Chile Bay, West Antarctic Peninsula. Microorganisms, 10(6), 1140. https://doi.org/10.3390/microorganisms10061140