
Clinical Infections, Antibiotic Resistance, and Pathogenesis of Staphylococcus haemolyticus

 , and
, and
Abstract
1. Introduction
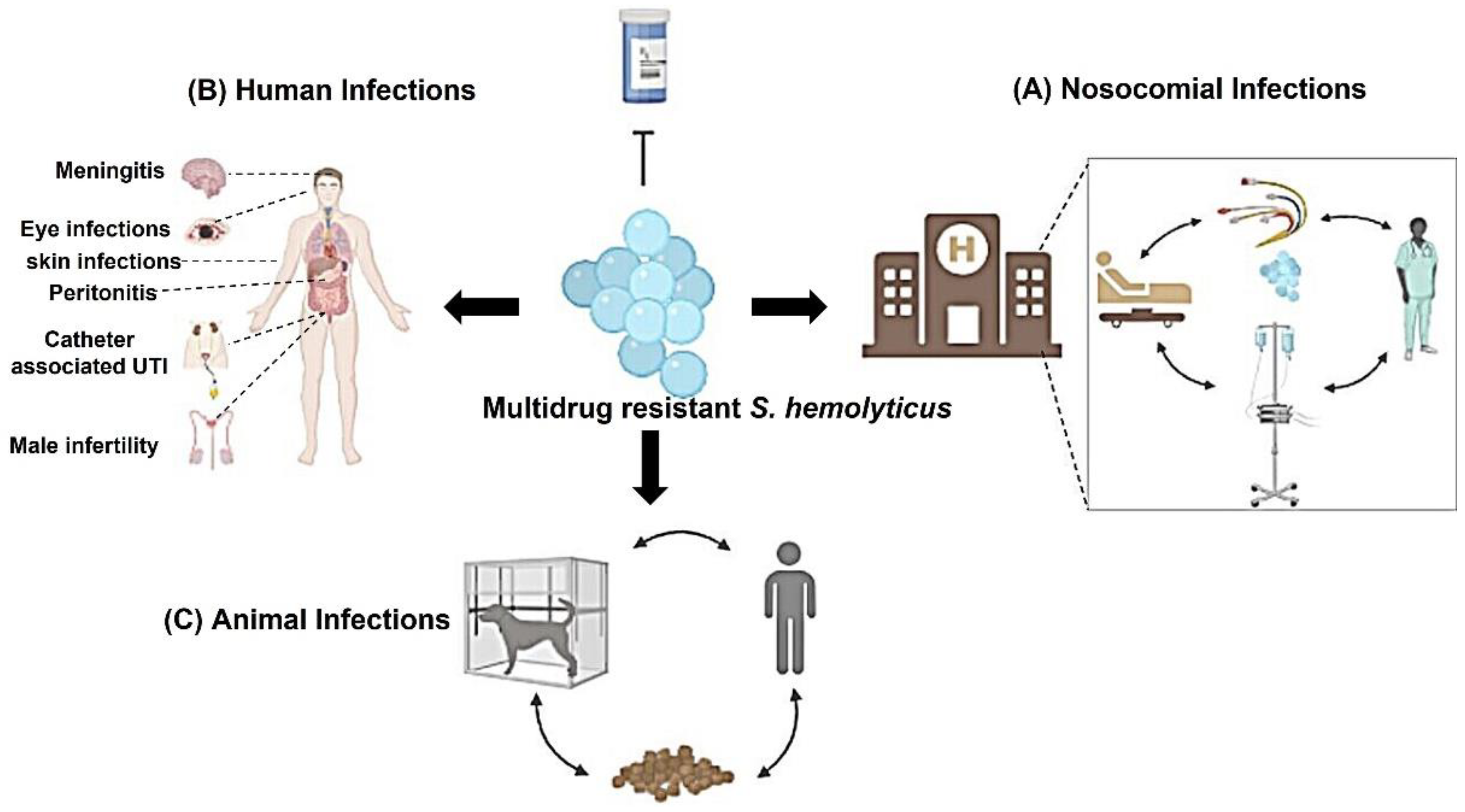
2. Clinical Infections Associated with S. haemolyticus
2.1. Bloodstream Infections (BSIs)
2.2. Eyes Infections
2.3. Nosocomial Infection
2.4. Male Infertility
2.5. Other Human Diseases
2.6. Animal Disease
3. Antibiotic Resistance in S. heamolyticus
3.1. β-Lactam
3.2. Methicillin
3.3. Glycopeptides
3.3.1. Vancomycin
3.3.2. Teicoplanin
3.4. Linezolid
3.5. Lincosamides
3.6. Mupirocin
4. Virulence Factors of S. haemolyticus
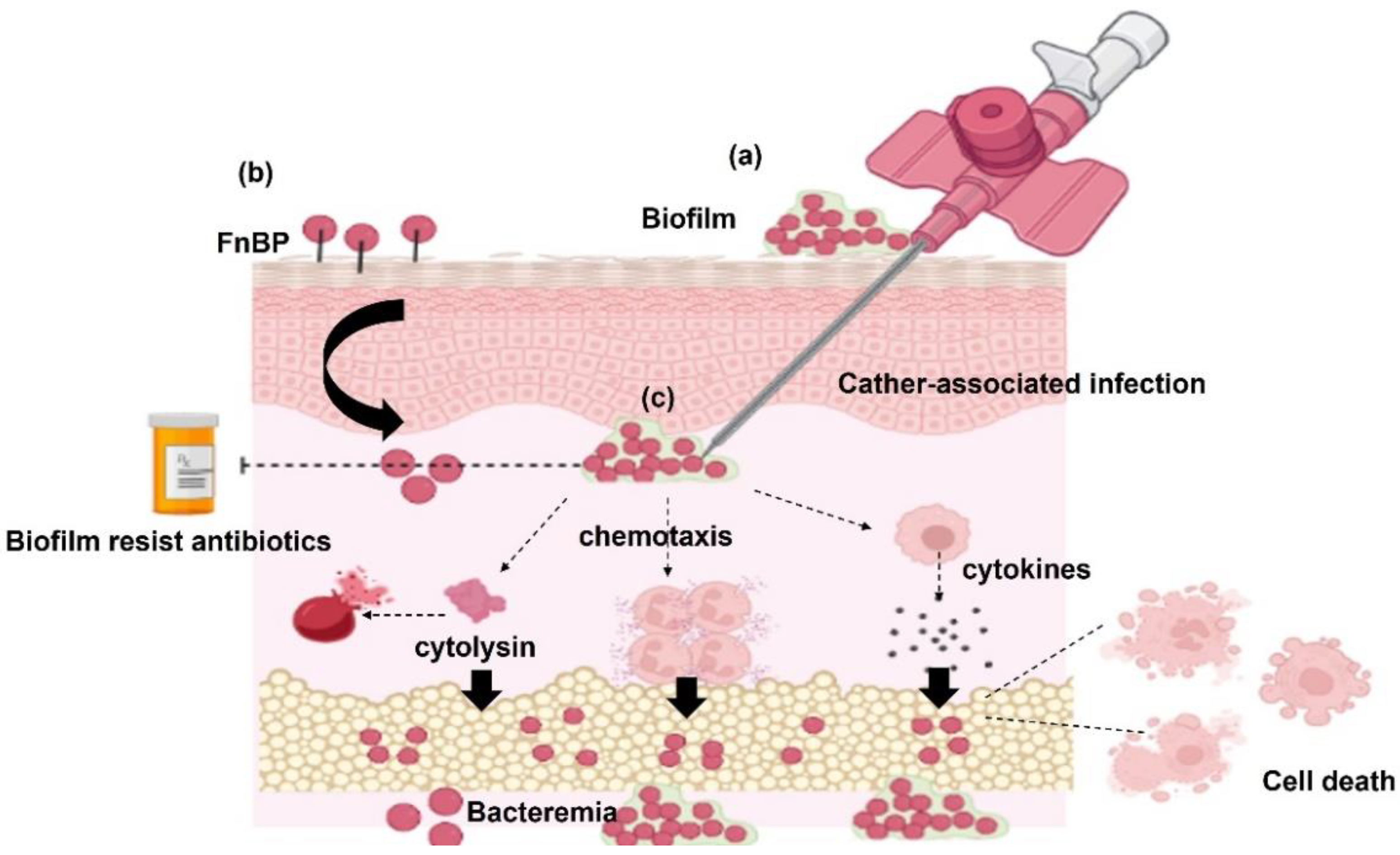
4.1. Biofilm Formation
4.2. S. haemolyticus Surface Proteins Required for Bacteria Adherence
4.3. Toxins and Enzymes
4.4. Cytotoxicity and Apoptosis of the Host Cells
5. Conclusions and Future Perspectives
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Aap | accumulation-associated protein |
| Ala | alanine amino acid |
| Bhp | biofilm-associated protein |
| BSIs | Bloodstream infections |
| CC1 | clonal complex 1 |
| CAPD | continuous ambulatory peritoneal dialysis |
| CoNS | Coagulase-negative Staph |
| CRS | catheter-related sepsis |
| eDNA | extracellular DNA |
| FnBP | fibronectin-binding proteins |
| GraS | glycopeptide resistance-associated histidine kinase |
| ica | intercellular adhesion |
| ICU | intensive care unit; IS: insertion sequence |
| MIC | minimum inhibitory concentration |
| MRSH | methicillin-resistant S. haemolyticus |
| PBP | penicillin-binding protein |
| PIA | polysaccharide intercellular adhesion |
| PSMs | phenol-soluble modulins |
| tcaRAB | teicoplanin resistance operon |
| tcaR | transcriptional regulator |
| TIRs | Toll/interleukin-1 like |
| SCCmec | Staphylococcal cassette chromosome mec |
| ST25 | sequence type 25 |
| S. | Staphylococcus. |
| UTIs | urinary tract infections |
References
- Eng, R.H.; Wang, C.; Person, A.; Kiehn, T.E.; Armstrong, D. Species identification of coagulase-negative staphylococcal isolates from blood cultures. J. Clin. Microbiol. 1982, 15, 439–442. [Google Scholar] [CrossRef] [PubMed]
- John, J.F., Jr.; Gramling, P.K.; O’Dell, N.M. Species identification of coagulase-negative staphylococci from urinary tract isolates. J. Clin. Microbiol. 1978, 8, 435–437. [Google Scholar] [CrossRef] [PubMed]
- Liekweg, W.G., Jr.; Greenfield, L.J. Vascular prosthetic infections: Collected experience and results of treatment. Surgery 1977, 81, 335–342. [Google Scholar] [PubMed]
- Rupp, M.E.; Archer, G.L. Coagulase-negative staphylococci: Pathogens associated with medical progress. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 1994, 19, 231–243, quiz 244–235. [Google Scholar] [CrossRef]
- Kloos, W.E.; Schleifer, K.H. Simplified scheme for routine identification of human Staphylococcus species. J. Clin. Microbiol. 1975, 1, 82–88. [Google Scholar] [CrossRef]
- Schaberg, D.R.; Culver, D.H.; Gaynes, R.P. Major trends in the microbial etiology of nosocomial infection. Am. J. Med. 1991, 91, 72S–75S. [Google Scholar] [CrossRef]
- Konstantinidis, K.T.; Ramette, A.; Tiedje, J.M. The bacterial species definition in the genomic era. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2006, 361, 1929–1940. [Google Scholar] [CrossRef]
- Lamers, R.P.; Muthukrishnan, G.; Castoe, T.A.; Tafur, S.; Cole, A.M.; Parkinson, C.L. Phylogenetic relationships among Staphylococcus species and refinement of cluster groups based on multilocus data. BMC Evol. Biol. 2012, 12, 171. [Google Scholar] [CrossRef]
- Ben Saida, N.; Marzouk, M.; Ferjeni, A.; Boukadida, J. A three-year surveillance of nosocomial infections by methicillin-resistant Staphylococcus haemolyticus in newborns reveals the disinfectant as a possible reservoir. Pathol. Biol. 2009, 57, 35. [Google Scholar] [CrossRef]
- Renaud, F.; Etienne, J.; Bertrand, A.; Brun, Y.; Greenland, T.B.; Freney, J.; Fleurette, J. Molecular epidemiology of Staphylococcus haemolyticus strains isolated in an Albanian hospital. J. Clin. Microbiol. 1991, 29, 1493–1497. [Google Scholar] [CrossRef]
- Szczuka, E.; Grabska, K.; Kaznowski, A. In Vitro Activity of Rifampicin Combined with Daptomycin or Tigecycline on Staphylococcus haemolyticus Biofilms. Curr. Microbiol. 2015, 71, 184–189. [Google Scholar] [CrossRef]
- Schuenck, R.P.; Pereira, E.M.; Iorio, N.; Santos, K. Multiplex PCR assay to identify methicillin-resistant Staphylococcus haemolyticus. FEMS Immunol. Med. Microbiol. 2008, 52, 431–435. [Google Scholar] [CrossRef] [PubMed]
- do Ferreira, N.; Schuenck, R.P.; dos Santos, K.; do de Bastos, M.; Giambiagi-deMarval, M. Diversity of plasmids and transmission of high-levelmupirocin mupA resistance gene in Staphylococcus haemolyticus. FEMS Immunol. Med. Microbiol. 2011, 61, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Ruzauskas, M.; Siugzdiniene, R.; Klimiene, I.; Virgailis, M.; Mockeliunas, R.; Vaskeviciute, L.; Zienius, D. Prevalence of methicillin-resistant Staphylococcus haemolyticus in companion animals: A cross-sectional study. Ann. Clin. Microbiol. Antimicrob. 2014, 13, 56. [Google Scholar] [CrossRef] [PubMed]
- Eltwisy, H.O.; Abdel-Fattah, M.; Elsisi, A.M.; Omar, M.M.; Abdelmoteleb, A.; El-Mokhtar, M.A. Pathogenesis of Staphylococcus haemolyticus on primary human skin fibroblast cells. Virulence 2020, 11, 1142–1157. [Google Scholar] [CrossRef]
- Cavanagh, J.P.; Hjerde, E.; Holden, M.T.G.; Kahlke, T.; Klingenberg, C.; Flaegstad, T.; Parkhill, J.; Bentley, S.D.; Sollid, J.U.E. Whole-genome sequencing reveals clonal expansion of multiresistant Staphylococcus haemolyticus in European hospitals. J. Antimicrob. Chemoth. 2014, 69, 2920–2927. [Google Scholar] [CrossRef]
- Hosseinkhani, F.; Jabalameli, F.; Farahani, N.N.; Taherikalani, M.; van Leeuwen, W.B.; Emaneini, M. Variable number of tandem repeat profiles and antimicrobial resistance patterns of Staphylococcus haemolyticus strains isolated from blood cultures in children. Infection 2016, 38, 19–21. [Google Scholar] [CrossRef]
- Ahmed, N.H.; Baruah, F.K.; Grover, R.K. Staphylococcal Blood Stream Infections in Cancer Patients. Ann. Med. Health Sci. Res. 2015, 5, 226–227. [Google Scholar] [CrossRef]
- Bryce, A.N.; Doocey, R.; Handy, R. Staphylococcus haemolyticus meningitis and bacteremia in an allogenic stem cell transplant patient. IDCases 2021, 26, e01259. [Google Scholar] [CrossRef]
- Viale, P.; Stefani, S. Vascular catheter-associated infections: A microbiological and therapeutic update. J. Chemother. 2006, 18, 235–249. [Google Scholar] [CrossRef]
- Sloos, J.H.; Dijkshoorn, L.; van Boven, C.P. Septicaemias caused by a strain of Staphylococcus haemolyticus exhibiting intermediate susceptibility to teicoplanin in multiple intensive care unit patients. J. Antimicrob. Chemother. 2000, 45, 410–411. [Google Scholar] [CrossRef] [PubMed]
- Spare, M.K.; Tebbs, S.E.; Lang, S.; Lambert, P.A.; Worthington, T.; Lipkin, G.W.; Elliott, T.S.J. Genotypic and phenotypic properties of coagulase-negative staphylococci causing dialysis catheter-related sepsis. J. Hosp. Infect. 2003, 54, 272–278. [Google Scholar] [CrossRef]
- Sader, H.S.; Jones, R.N.; Gales, A.C.; Silva, J.B.; Pignatari, A.C.; America, S. SENTRY antimicrobial surveillance program report: Latin american and brazilian results for 1997 through 2001. Braz. J. Infect. Dis. 2004, 8, 25–79. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Satti, L.; Zaman, G.; Gardezi, A.; Sabir, N.; Khadim, M. Catheter related recurrent blood stream infection caused by linezolid-resistant, methicillin resistant Staphylococcus haemolyticus; an emerging super bug. JPMA J. Pak. Med. Assoc. 2019, 69, 261–263. [Google Scholar]
- Panda, S.; Kar, S.; Sharma, S.; Singh, D.V. Multidrug-resistant Staphylococcus haemolyticus isolates from infected eyes and healthy conjunctivae in India. J. Glob. Antimicrob. Resist. 2016, 6, 154–159. [Google Scholar] [CrossRef]
- Malathi, J.; Sowmiya, M.; Margarita, S.; Madhavan, H.N.; Therese, K.L. Application of PCR based-RFLP for species identification of ocular isolates of methicillin resistant staphylococci (MRS). Indian J. Med. Res. 2009, 130, 78–84. [Google Scholar]
- Makki, A.R.; Sharma, S.; Duggirala, A.; Prashanth, K.; Garg, P.; Das, T. Phenotypic and genotypic characterization of coagulase negative staphylococci (CoNS) other than Staphylococcus epidermidis isolated from ocular infections. Investig. Ophthalmol. Vis. Sci. 2011, 52, 9018–9022. [Google Scholar] [CrossRef]
- Wong, M.; Baumrind, B.R.; Frank, J.H.; Halpern, R.L. Postoperative Endophthalmitis Caused by Staphylococcus haemolyticus following Femtosecond Cataract Surgery. Case Rep. Ophthalmol. 2015, 6, 435–438. [Google Scholar] [CrossRef]
- Panda, S.; Jena, S.; Sharma, S.; Dhawan, B.; Nath, G.; Singh, D.V. Identification of Novel Sequence Types among Staphylococcus haemolyticus Isolated from Variety of Infections in India. PLoS ONE 2016, 11, e0166193. [Google Scholar] [CrossRef]
- Couto, N.; Chlebowicz, M.A.; Raangs, E.C.; Friedrich, A.W.; Rossen, J.W. Complete Genome Sequences of Two Methicillin-Resistant Staphylococcus haemolyticus Isolates of Multilocus Sequence Type 25, First Detected by Shotgun Metagenomics. Genome Announc. 2018, 6, e00036-18. [Google Scholar] [CrossRef]
- Spencer, S.R.; Dealler, S.F.; Hassett, P.D.; Todd, N.J.; Hawkey, P.M.; Noble, B.A. Bacterial contamination of intraocular lenses: The source of the bacteria. Eye 1989, 3, 685–689. [Google Scholar] [CrossRef] [PubMed]
- Kodjikian, L.; Burillon, C.; Roques, C.; Pellon, G.; Freney, J.; Renaud, F.N. Bacterial adherence of Staphylococcus epidermidis to intraocular lenses: A bioluminescence and scanning electron microscopy study. Invest. Ophthalmol. Vis. Sci. 2003, 44, 4388–4394. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.; Singh, D. Biofilm Formation by ica-Negative Ocular Isolates of Staphylococcus haemolyticus. Front. Microbiol. 2018, 9, 2687. [Google Scholar] [CrossRef] [PubMed]
- Brook, I. Bacteriology of neonatal omphalitis. J. Infect. 1982, 5, 127–131. [Google Scholar] [CrossRef]
- Bjorkqvist, M.; Liljedahl, M.; Zimmermann, J.; Schollin, J.; Soderquist, B. Colonization pattern of coagulase-negative staphylococci in preterm neonates and the relation to bacteremia. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2010, 29, 1085–1093. [Google Scholar] [CrossRef]
- Cave, R.; Misra, R.; Chen, J.; Wang, S.; Mkrtchyan, H.V. Whole genome sequencing revealed new molecular characteristics in multidrug resistant staphylococci recovered from high frequency touched surfaces in London. Sci. Rep. 2019, 9, 9637. [Google Scholar] [CrossRef]
- Mehri, H.; Jahanbakhsh, R.; Shakeri, F.; Ardebili, A.; Behnampour, N.; Khodabakhshi, B.; Ghaemi, E.A. Investigation of Glycopeptide Susceptibility of Coagulase-Negative Staphylococci (CoNS) From a Tertiary Care Hospital in Gorgan, Northern Iran. Arch. Pediatr. Infect. 2017, 5, e37264. [Google Scholar] [CrossRef]
- Soeorg, H.; Huik, K.; Parm, Ü.; Ilmoja, M.-L.; Metelskaja, N.; Metsvaht, T.; Lutsar, I. Genetic Relatedness of Coagulase-negative Staphylococci From Gastrointestinal Tract and Blood of Preterm Neonates With Late-onset Sepsis. Pediatric Infect. Dis. J. 2013, 32, 389–393. [Google Scholar] [CrossRef]
- Perdreau-Remington, F.; Stefanik, D.; Peters, G.; Ruckdeschel, G.; Haas, F.; Wenzel, R.; Pulverer, G. Methicillin-resistant Staphylococcus haemolyticus on the hands of health care workers: A route of transmission or a source? J. Hosp. Infect. 1995, 31, 195–203. [Google Scholar] [CrossRef]
- Klingenberg, C.; Rønnestad, A.; Anderson, A.S.; Abrahamsen, T.G.; Zorman, J.; Villaruz, A.; Flaegstad, T.; Otto, M.; Sollid, J.E. Persistent strains of coagulase-negative staphylococci in a neonatal intensive care unit: Virulence factors and invasiveness. Clin. Microbiol. Infect. 2007, 13, 1100–1111. [Google Scholar] [CrossRef]
- Daniel, B.; Saleem, M.; Naseer, G.; Fida, A. Significance of Staphylococcus Haemolyticus in Hospital Acquired Infections. J. Pioneer Med. Sci. 2014, 4, 119–125. [Google Scholar]
- Mazzariol, A.; Lo Cascio, G.; Kocsis, E.; Maccacaro, L.; Fontana, R.; Cornaglia, G. Outbreak of linezolid-resistant Staphylococcus haemolyticus in an Italian intensive care unit. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2012, 31, 523–527. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, J.P.; Wolden, R.; Heise, P.; Esaiassen, E.; Klingenberg, C.; Aarag Fredheim, E.G. Antimicrobial susceptibility and body site distribution of community isolates of coagulase-negative staphylococci. APMIS Acta Pathol. Microbiol. Immunol. Scand. 2016, 124, 973–978. [Google Scholar] [CrossRef]
- Degener, J.E.; Heck, M.E.; van Leeuwen, W.J.; Heemskerk, C.; Crielaard, A.; Joosten, P.; Caesar, P. Nosocomial infection by Staphylococcus haemolyticus and typing methods for epidemiological study. J. Clin. Microbiol. 1994, 32, 2260–2265. [Google Scholar] [CrossRef] [PubMed]
- Dziri, R.; Klibi, N.; Lozano, C.; Ben Said, L.; Bellaaj, R.; Tenorio, C.; Boudabous, A.; Ben Slama, K.; Torres, C. High prevalence of Staphylococcus haemolyticus and Staphylococcus saprophyticus in environmental samples of a Tunisian hospital. Diagn. Microbiol. Infect. Dis. 2016, 85, 136–140. [Google Scholar] [CrossRef]
- Bouchami, O.; de Lencastre, H.; Miragaia, M. Impact of Insertion Sequences and Recombination on the Population Structure of Staphylococcus haemolyticus. PLoS ONE 2016, 11, e0156653. [Google Scholar] [CrossRef]
- Fraczek, M.; Piasecka, M.; Gaczarzewicz, D.; Szumala-Kakol, A.; Kazienko, A.; Lenart, S.; Laszczynska, M.; Kurpisz, M. Membrane stability and mitochondrial activity of human-ejaculated spermatozoa during in vitro experimental infection with Escherichia coli, Staphylococcus haemolyticus and Bacteroides ureolyticus. Andrologia 2012, 44, 315–329. [Google Scholar] [CrossRef]
- Fraczek, M.; Wiland, E.; Piasecka, M.; Boksa, M.; Gaczarzewicz, D.; Szumala-Kakol, A.; Kolanowski, T.; Beutin, L.; Kurpisz, M. Fertilizing potential of ejaculated human spermatozoa during in vitro semen bacterial infection. Fertil. Steril. 2014, 102, 711–719.E1. [Google Scholar] [CrossRef]
- Fraczek, M.; Hryhorowicz, M.; Gaczarzewicz, D.; Szumala-Kakol, A.; Kolanowski, T.J.; Beutin, L.; Kurpisz, M. Can apoptosis and necrosis coexist in ejaculated human spermatozoa during in vitro semen bacterial infection? J. Assist. Reprod. Genet. 2015, 32, 771–779.e711. [Google Scholar] [CrossRef]
- Pindar, C.; Viau, R.A. Staphylococcus haemolyticus epididymo-orchitis and bacteraemia: A case report. JMM Case Rep. 2018, 5, e005157. [Google Scholar] [CrossRef]
- Mazzoli, S. Biofilms in chronic bacterial prostatitis (NIH-II) and in prostatic calcifications. FEMS Immunol. Med. Microbiol. 2010, 59, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, E.; Ribes-Koninckx, C.; Calabuig, M.; Sanz, Y. Intestinal Staphylococcus spp. and virulent features associated with coeliac disease. J. Clin. Pathol. 2012, 65, 830. [Google Scholar] [CrossRef] [PubMed]
- Shittu, A.; Lin, J.; Morrison, D.; Kolawole, D. Isolation and molecular characterization of multiresistant Staphylococcus sciuri and Staphylococcus haemolyticus associated with skin and soft-tissue infections. J. Med. Microbiol. 2004, 53, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Gunn, B.A.; Davis, C.E. Staphylococcus haemolyticus urinary tract infection in a male patient. J. Clin. Microbiol. 1988, 26, 1055–1057. [Google Scholar] [CrossRef]
- Takeuchi, F.; Watanabe, S.; Baba, T.; Yuzawa, H.; Ito, T.; Morimoto, Y.; Kuroda, M.; Cui, L.; Takahashi, M.; Ankai, A.; et al. Whole-genome sequencing of staphylococcus haemolyticus uncovers the extreme plasticity of its genome and the evolution of human-colonizing staphylococcal species. J. Bacteriol. 2005, 187, 7292–7308. [Google Scholar] [CrossRef]
- Suen, K.; Mashhadian, A.; Figarsky, I.; Payumo, J.; Liu, A. A rare and important case of Staphylococcus haemolyticus-associated ventricular atrial shunt nephritis. Clin. Case Rep. 2017, 5, 2012–2016. [Google Scholar] [CrossRef]
- Gamberini, S.; Anania, G.; Incasa, E.; Zangirolami, A.; Tampieri, M.; Boari, B.; Benea, G.; Manfredini, R. Staphylococcus hemolyticus liver abscess as an uncommon presentation of silent colonic cancer: A case report. J. Am. Geriatr. Soc. 2006, 54, 1619–1620. [Google Scholar] [CrossRef]
- Bhargava, K.; Zhang, Y. Multidrug-resistant coagulase-negative Staphylococci in food animals. J. Appl. Microbiol. 2012, 113, 1027–1036. [Google Scholar] [CrossRef]
- Hope, R.; Livermore, D.M.; Brick, G.; Lillie, M.; Reynolds, R.; Su, B.W.P.R. Non-susceptibility trends among staphylococci from bacteraemias in the UK and Ireland, 2001–2006. J. Antimicrob. Chemoth. 2008, 62, Ii65–Ii74. [Google Scholar] [CrossRef]
- Bouchami, O.; Achour, W.; Mekni, M.A.; Rolo, J.; Ben Hassen, A. Antibiotic resistance and molecular characterization of clinical isolates of methicillin-resistant coagulase-negative staphylococci isolated from bacteremic patients in oncohematology. Folia Microbiol. 2011, 56, 122–130. [Google Scholar] [CrossRef]
- Rodriguez-Aranda, A.; Daskalaki, M.; Villar, J.; Sanz, F.; Otero, J.R.; Chaves, F. Nosocomial spread of linezolid-resistant Staphylococcus haemolyticus infections in an intensive care unit. Diagn. Microbiol. Infect. Dis. 2009, 63, 398–402. [Google Scholar] [CrossRef] [PubMed]
- Czekaj, T.; Ciszewski, M.; Szewczyk, E.M. Staphylococcus haemolyticus—An emerging threat in the twilight of the antibiotics age. Microbiology 2015, 161, 2061–2068. [Google Scholar] [CrossRef] [PubMed]
- Hosseinkhani, F.; Emaneini, M.; van Leeuwen, W. High-Quality Genome Sequence of the Highly Resistant Bacterium Staphylococcus haemolyticus, Isolated from a Neonatal Bloodstream Infection. Genome Announc. 2017, 5, e00683-17. [Google Scholar] [CrossRef] [PubMed]
- Bakthavatchalam, Y.D.; Sudarsanam, T.D.; Babu, P.; Munuswamy, E.; Muthuirulandi Sethuvel, D.P.; Devanga Ragupathi, N.K.; Veeraraghavan, B. Methicillin-Susceptible Teicoplanin-Resistant Staphylococcus haemolyticus Isolate from a Bloodstream Infection with Novel Mutations in the tcaRAB Teicoplanin Resistance Operon. Jpn J. Infect. Dis. 2017, 70, 458–460. [Google Scholar] [CrossRef] [PubMed]
- Anthonisen, I.L.; Sunde, M.; Steinum, T.M.; Sidhu, M.S.; Sørum, H. Organization of the antiseptic resistance gene qacA and Tn552-related beta-lactamase genes in multidrug- resistant Staphylococcus haemolyticus strains of animal and human origins. Antimicrob. Agents Chemother. 2002, 46, 3606–3612. [Google Scholar] [CrossRef]
- Falcone, M.; Giannella, M.; Raponi, G.; Mancini, C.; Venditti, M. Teicoplanin use and emergence of Staphylococcus haemolyticus: Is there a link? Clin. Microbiol. Infect. 2006, 12, 96–97. [Google Scholar] [CrossRef]
- Rossi, C.C.; Ferreira, N.C.; Coelho, M.L.; Schuenck, R.P.; Bastos Mdo, C.; Giambiagi-deMarval, M. Transfer of mupirocin resistance from Staphylococcus haemolyticus clinical strains to Staphylococcus aureus through conjugative and mobilizable plasmids. FEMS Microbiol. Lett. 2016, 363, fnw121. [Google Scholar] [CrossRef]
- Kim, H.J.; Jang, S. Draft genome sequence of multidrug-resistant Staphylococcus haemolyticus IPK_TSA25 harbouring a Staphylococcus aureus plasmid, pS0385-1. J. Glob. Antimicrob. Resist. 2017, 11, 8–9. [Google Scholar] [CrossRef]
- Correa, J.E.; Paulis, D.A.; Predari, S.; Sordelli, D.O.; Jeric, P.E. First report of qacG, qacH and qacJ genes in Staphylococcus haemolyticus human clinical isolates. J. Antimicrob. Chemoth. 2008, 62, 956–960. [Google Scholar] [CrossRef]
- Bochniarz, M.; Wawron, W.; Szczubial, M. Resistance to methicillin of coagulase-negative staphylococci (CNS) isolated from bovine mastitis. Pol. J. Vet. Sci. 2013, 16, 687–692. [Google Scholar] [CrossRef][Green Version]
- Billot-Klein, D.; Gutmann, L.; Bryant, D.; Bell, D.; Van Heijenoort, J.; Grewal, J.; Shlaes, D.M. Peptidoglycan synthesis and structure in Staphylococcus haemolyticus expressing increasing levels of resistance to glycopeptide antibiotics. J. Bacteriol. 1996, 178, 4696–4703. [Google Scholar] [CrossRef] [PubMed]
- Quiles-Melero, I.; Gómez-Gil, R.; Romero-Gómez, M.P.; Sánchez-Díaz, A.M.; Pablos, M.d.; García-Rodriguez, J.; Gutiérrez, A.; Mingorance, J. Mechanisms of Linezolid Resistance among Staphylococci in a Tertiary Hospital. J. Clin. Microbiol. 2013, 51, 998–1001. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Rawre, J.; Trikha, A.; Sreenivas, V.; Sood, S.; Kapil, A.; Dhawan, B. Linezolid-resistant Staphylococcus haemolyticus: Emergence of G2447U & C2534U mutations at the domain V of 23S ribosomal RNA gene in a tertiary care hospital in India. Indian J. Med. Res. 2019, 149, 795–798. [Google Scholar] [CrossRef] [PubMed]
- Brijwal, M.; Dhawan, B.; Rawre, J.; Sebastian, S.; Kapil, A. Clonal dissemination of linezolid-resistant Staphylococcus haemolyticus harbouring a G2576T mutation and the cfr gene in an Indian hospital. J. Med. Microbiol. 2016, 65, 698–700. [Google Scholar] [CrossRef]
- Long, K.S.; Vester, B. Resistance to linezolid caused by modifications at its binding site on the ribosome. Antimicrob. Agents Chemother. 2012, 56, 603–612. [Google Scholar] [CrossRef]
- Brisson-Noel, A.; Delrieu, P.; Samain, D.; Courvalin, P. Inactivation of lincosaminide antibiotics in Staphylococcus. Identification of lincosaminide O-nucleotidyltransferases and comparison of the corresponding resistance genes. J. Biol. Chem. 1988, 263, 15880–15887. [Google Scholar] [CrossRef]
- Brisson-Noël, A.; Courvalin, P. Nucleotide sequence of gene linA encoding resistance to lincosamides in Staphylococcus haemolyticus. Gene 1986, 43, 247–253. [Google Scholar] [CrossRef]
- Novotna, G.; Janata, J. A New Evolutionary Variant of the Streptogramin A Resistance Protein, Vga(A)LC, from Staphylococcus haemolyticus with Shifted Substrate Specificity towards Lincosamides. Antimicrob. Agents Chemother. 2006, 50, 4070–4076. [Google Scholar] [CrossRef]
- Thomas, C.M.; Hothersall, J.; Willis, C.L.; Simpson, T.J. Resistance to and synthesis of the antibiotic mupirocin. Nat. Rev. Microbiol. 2010, 8, 281–289. [Google Scholar] [CrossRef]
- Barros, E.M.; Ceotto, H.; Bastos, M.C.F.; dos Santos, K.R.N.; Giambiagi-deMarval, M. Staphylococcus haemolyticus as an Important Hospital Pathogen and Carrier of Methicillin Resistance Genes. J. Clin. Microbiol. 2012, 50, 166–168. [Google Scholar] [CrossRef]
- Manoharan, M.; Sistla, S.; Ray, P. Prevalence and Molecular Determinants of Antimicrobial Resistance in Clinical Isolates of Staphylococcus haemolyticus from India. Microb. Drug Resist. 2021, 27, 501–508. [Google Scholar] [CrossRef] [PubMed]
- De Vecchi, E.; George, D.A.; Romanò, C.L.; Pregliasco, F.E.; Mattina, R.; Drago, L. Antibiotic sensitivities of coagulase-negative staphylococci and Staphylococcus aureus in hip and knee periprosthetic joint infections: Does this differ if patients meet the International Consensus Meeting Criteria? Infect. Drug Resist. 2018, 11, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Stewart, G.T. Changes in Sensitivity of Staphylococci to Methicillin. Br. Med. J. 1961, 1, 863. [Google Scholar] [CrossRef]
- John, J.F.; Harvin, A.M. History and evolution of antibiotic resistance in coagulase-negative staphylococci: Susceptibility profiles of new anti-staphylococcal agents. Ther. Clin. Risk Manag. 2007, 3, 1143–1152. [Google Scholar] [PubMed]
- Hanssen, A.M.; Ericson Sollid, J.U. SCCmec in staphylococci: Genes on the move. FEMS Immunol. Med. Microbiol. 2006, 46, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Katayama, Y.; Asada, K.; Mori, N.; Tsutsumimoto, K.; Tiensasitorn, C.; Hiramatsu, K. Structural comparison of three types of staphylococcal cassette chromosome mec integrated in the chromosome in methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2001, 45, 1323–1336. [Google Scholar] [CrossRef]
- Zong, Z.; Peng, C.; Lü, X. Diversity of SCCmec elements in methicillin-resistant coagulase-negative staphylococci clinical isolates. PLoS ONE 2011, 6, e20191. [Google Scholar] [CrossRef]
- Kim, J.S.; Kim, H.S.; Park, J.Y.; Koo, H.S.; Choi, C.S.; Song, W.; Cho, H.C.; Lee, K.M. Contamination of X-ray cassettes with methicillin-resistant Staphylococcus aureus and methicillin-resistant Staphylococcus haemolyticus in a radiology department. Ann. Lab. Med. 2012, 32, 206–209. [Google Scholar] [CrossRef]
- Koksal, F.; Yasar, H.; Samasti, M. Antibiotic resistance patterns of coagulase-negative staphylococcus strains isolated from blood cultures of septicemic patients in Turkey. Microbiol. Res. 2009, 164, 404–410. [Google Scholar] [CrossRef]
- Berglund, C.; Söderquist, B. The origin of a methicillin-resistant Staphylococcus aureus isolate at a neonatal ward in Sweden—possible horizontal transfer of a staphylococcal cassette chromosome mec between methicillin-resistant Staphylococcus haemolyticus and Staphylococcus aureus. Clin. Microbiol. Infect. 2008, 14, 1048–1056. [Google Scholar] [CrossRef]
- Guggenheim, M.; Zbinden, R.; Handschin, A.E.; Burns, G.-A. Changes in bacterial isolates from burn wounds and their antibiograms: A 20-year study (1986–2005). Burns 2009, 35, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Kresken, M.; Becker, K.; Seifert, H.; Leitner, E.; Korber-Irrgang, B.; von Eiff, C.; Loschmann, P.A.; Study, G. Resistance trends and in vitro activity of tigecycline and 17 other antimicrobial agents against Gram-positive and Gram-negative organisms, including multidrug-resistant pathogens, in Germany. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2011, 30, 1095–1103. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sader, H.S.; Jones, R.N. Antimicrobial activity of daptomycin in comparison to glycopeptides and other antimicrobials when tested against numerous species of coagulase-negative Staphylococcus. Diagn. Microbiol. Infect. Dis. 2012, 73, 212–214. [Google Scholar] [CrossRef] [PubMed]
- von Eiff, C.; Friedrich, A.W.; Becker, K.; Peters, G. Comparative in vitro activity of ceftobiprole against staphylococci displaying normal and small-colony variant phenotypes. Antimicrob. Agents Chemother. 2005, 49, 4372–4374. [Google Scholar] [CrossRef]
- James, R.C.; Pierce, J.G.; Okano, A.; Xie, J.; Boger, D.L. Redesign of glycopeptide antibiotics: Back to the future. ACS Chem. Biol. 2012, 7, 797–804. [Google Scholar] [CrossRef]
- Tacconelli, E.; Tumbarello, M.; de Gaetano Donati, K.; Bettio, M.; Spanu, T.; Leone, F.; Sechi, L.A.; Zanetti, S.; Fadda, G.; Cauda, R. Glycopeptide Resistance among Coagulase-Negative Staphylococci that Cause Bacteremia: Epidemiological and Clinical Findings from a Case-Control Study. Clin. Infect. Dis. 2001, 33, 1628–1635. [Google Scholar] [CrossRef]
- Aubert, G.; Passot, S.; Lucht, F.; Dorche, G. Selection of vancomycin- and teicoplanin-resistant Staphylococcus haemolyticus during teicoplanin treatment of S. epidermidis infection. J. Antimicrob. Chemother. 1990, 25, 491–493. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Staphylococcus aureus resistant to vancomycin--United States, 2002. MMWR. Morb. Mortal. Wkly. Rep. 2002, 51, 565–567. [Google Scholar]
- Schwalbe, R.S.; Stapleton, J.T.; Gilligan, P.H. Emergence of vancomycin resistance in coagulase-negative staphylococci. N. Engl. J. Med. 1987, 316, 927–931. [Google Scholar] [CrossRef]
- Sanyal, D.; Greenwood, D. An electronmicroscope study of glycopeptide antibiotic-resistant strains of Staphylococcus epidermidis. J. Med. Microbiol. 1993, 39, 204–210. [Google Scholar] [CrossRef]
- Giovanetti, E.; Biavasco, F.; Pugnaloni, A.; Lupidi, R.; Biagini, G.; Varaldo, P.E. An electron microscopic study of clinical and laboratory-derived strains of teicoplanin-resistant Staphylococcus haemolyticus. Microb. Drug Resist. 1996, 2, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Biavasco, F.; Vignaroli, C.; Varaldo, P.E. Glycopeptide resistance in coagulase-negative staphylococci. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2000, 19, 403–417. [Google Scholar] [CrossRef]
- Cunningham, R.; Gurnell, M.; Bayston, R.; Cockayne, A.; Shelton, A. Teicoplanin resistance in Staphylococcus haemolyticus, developing during treatment. J. Antimicrob. Chemother. 1997, 39, 438–439. [Google Scholar] [CrossRef] [PubMed]
- Schwalbe, R.S.; Ritz, W.J.; Verma, P.R.; Barranco, E.A.; Gilligan, P.H. Selection for vancomycin resistance in clinical isolates of Staphylococcus haemolyticus. J. Infect. Dis. 1990, 161, 45–51. [Google Scholar] [CrossRef]
- Tsakris, A.; Papadimitriou, E.; Douboyas, J.; Antoniadis, A. Emergence of teicoplanin-resistant Staphylococcus haemolyticus clinical isolates in Greece. J. Antimicrob. Chemother. 2000, 46, 1040–1041. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Blans, M.; Troelstra, A. Glycopeptide resistance in Staphylococcus haemolyticus during treatment with teicoplanin. Infect. Control Hosp. Epidemiol. 2001, 22, 263–264. [Google Scholar] [CrossRef][Green Version]
- Rajan, V.; Kumar, V.G.; Gopal, S. A cfr-positive clinical staphylococcal isolate from India with multiple mechanisms of linezolid-resistance. Indian J. Med. Res. 2014, 139, 463–467. [Google Scholar]
- Matlani, M.; Shende, T.; Bhandari, V.; Dawar, R.; Sardana, R.; Gaind, R. Linezolid-resistant mucoid Staphylococcus haemolyticus from a tertiary-care centre in Delhi. New Microbes New Infect. 2016, 11, 57–58. [Google Scholar] [CrossRef][Green Version]
- de Almeida, L.M.; Lincopan, N.; de Araújo, M.R.; Mamizuka, E.M. Clonal dissemination of linezolid-resistant Staphylococcus haemolyticus exhibiting the G2576T mutation in the 23S rRNA gene in a tertiary care hospital in Brazil. Antimicrob. Agents Chemother. 2012, 56, 2792–2793. [Google Scholar] [CrossRef]
- Kehrenberg, C.; Schwarz, S.; Jacobsen, L.; Hansen, L.H.; Vester, B. A new mechanism for chloramphenicol, florfenicol and clindamycin resistance: Methylation of 23S ribosomal RNA at A2503. Mol. Microbiol. 2005, 57, 1064–1073. [Google Scholar] [CrossRef]
- Leclercq, R.; Carlier, C.; Duval, J.; Courvalin, P. Plasmid-mediated resistance to lincomycin by inactivation in Staphylococcus haemolyticus. Antimicrob. Agents Chemother. 1985, 28, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Novotna, G.; Adamkova, V.; Janata, J.; Melter, O.; Spizek, J. Prevalence of resistance mechanisms against macrolides and lincosamides in methicillin-resistant coagulase-negative staphylococci in the Czech Republic and occurrence of an undefined mechanism of resistance to lincosamides. Antimicrob. Agents Chemother. 2005, 49, 3586–3589. [Google Scholar] [CrossRef] [PubMed]
- Bean, D.C.; Wigmore, S.M.; Wareham, D.W. Draft Genome Sequence of a Canine Isolate of Methicillin-Resistant Staphylococcus haemolyticus. Genome Announc. 2017, 5, e00146-17. [Google Scholar] [CrossRef] [PubMed]
- Kizerwetter-Świda, M.; Chrobak-Chmiel, D.; Rzewuska, M. High-level mupirocin resistance in methicillin-resistant staphylococci isolated from dogs and cats. BMC Vet. Res. 2019, 15, 238. [Google Scholar] [CrossRef]
- Chan, K.-G.; Ng, K.; Chong, T.; Pang, Y.; Kamarulzaman, A.; Yin, W.-F.; Tee, K. Antibiotic Resistant and Virulence Determinants of Staphylococcus haemolyticus C10A as Revealed by Whole Genome Sequencing. J. Genom. 2015, 3, 72–74. [Google Scholar] [CrossRef]
- Da, F.; Joo, H.-S.; Cheung, G.Y.C.; Villaruz, A.E.; Rohde, H.; Luo, X.; Otto, M. Phenol-Soluble Modulin Toxins of Staphylococcus haemolyticus. Front. Cell. Infect. Microbiol. 2017, 7, 206. [Google Scholar] [CrossRef]
- Pereira-Ribeiro, P.M.; Sued-Karam, B.R.; Faria, Y.V.; Nogueira, B.A.; Colodette, S.S.; Fracalanzza, S.E.; Duarte, J.L.; Junior, R.H.; Mattos-Guaraldi, A.L. Influence of antibiotics on biofilm formation by different clones of nosocomial Staphylococcus haemolyticus. Future Microbiol. 2019, 14, 789–799. [Google Scholar] [CrossRef]
- Fredheim, E.; Klingenberg, C.; Rohde, H.; Frankenberger, S.; Gaustad, P.; Flægstad, T.; Sollid, J. Biofilm Formation by Staphylococcus haemolyticus. J. Clin. Microbiol. 2009, 47, 1172–1180. [Google Scholar] [CrossRef]
- Silva, P.V.; Cruz, R.S.; Keim, L.S.; Paula, G.R.; Carvalho, B.T.; Coelho, L.R.; Carvalho, M.C.; Rosa, J.M.; Figueiredo, A.M.; Teixeira, L.A. The antimicrobial susceptibility, biofilm formation and genotypic profiles of Staphylococcus haemolyticus from bloodstream infections. Mem. Inst. Oswaldo Cruz 2013, 108, 812–813. [Google Scholar] [CrossRef]
- Flahaut, S.; Vinogradov, E.; Kelley, K.A.; Brennan, S.; Hiramatsu, K.; Lee, J.C. Structural and biological characterization of a capsular polysaccharide produced by Staphylococcus haemolyticus. J. Bacteriol. 2008, 190, 1649–1657. [Google Scholar] [CrossRef]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [PubMed]
- de Araujo, G.L.; Coelho, L.R.; de Carvalho, C.B.; Maciel, R.M.; Coronado, A.Z.; Rozenbaum, R.; Ferreira-Carvalho, B.T.; Figueiredo, A.M.; Teixeira, L.A. Commensal isolates of methicillin-resistant Staphylococcus epidermidis are also well equipped to produce biofilm on polystyrene surfaces. J. Antimicrob. Chemother. 2006, 57, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Wolden, R.; Pain, M.; Karlsson, R.; Karlsson, A.; Aarag Fredheim, E.G.; Cavanagh, J.P. Identification of surface proteins in a clinical Staphylococcus haemolyticus isolate by bacterial surface shaving. BMC Microbiol. 2020, 20, 80. [Google Scholar] [CrossRef] [PubMed]
- Valle, J.; Gomez-Lucia, E.; Piriz, S.; Goyache, J.; Orden, J.A.; Vadillo, S. Enterotoxin production by staphylococci isolated from healthy goats. Appl. Environ. Microbiol. 1990, 56, 1323–1326. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, L.; Brito, C.I.; de Oliveira, A.; Martins, P.Y.; Pereira, V.C.; da Cunha Mde, L. Staphylococcus epidermidis and Staphylococcus haemolyticus: Molecular Detection of Cytotoxin and Enterotoxin Genes. Toxins 2015, 7, 3688–3699. [Google Scholar] [CrossRef]
- da Cunha Mde, L.; Calsolari, R.A.; Júnior, J.P. Detection of enterotoxin and toxic shock syndrome toxin 1 genes in Staphylococcus, with emphasis on coagulase-negative staphylococci. Microbiol. Immunol. 2007, 51, 381–390. [Google Scholar] [CrossRef]
- Taylor, A.L.; Llewelyn, M.J. Superantigen-induced proliferation of human CD4+CD25- T cells is followed by a switch to a functional regulatory phenotype. J. Immunol. 2010, 185, 6591–6598. [Google Scholar] [CrossRef]
- de Freitas Guimarães, F.; Nóbrega, D.B.; Richini-Pereira, V.B.; Marson, P.M.; de Figueiredo Pantoja, J.C.; Langoni, H. Enterotoxin genes in coagulase-negative and coagulase-positive staphylococci isolated from bovine milk. J. Dairy Sci. 2013, 96, 2866–2872. [Google Scholar] [CrossRef]
- Barretti, P.; Montelli, A.C.; Batalha, J.E.; Caramori, J.C.; Cunha Mde, L. The role of virulence factors in the outcome of staphylococcal peritonitis in CAPD patients. BMC Infect. Dis. 2009, 9, 212. [Google Scholar] [CrossRef]
- Vasconcelos, N.G.; Pereira, V.C.; Araújo Júnior, J.P.; da Cunha Mde, L. Molecular detection of enterotoxins E, G, H and I in Staphylococcus aureus and coagulase-negative staphylococci isolated from clinical samples of newborns in Brazil. J. Appl. Microbiol. 2011, 111, 749–762. [Google Scholar] [CrossRef]
- Breuer, K.; Wittmann, M.; Kempe, K.; Kapp, A.; Mai, U.; Dittrich-Breiholz, O.; Kracht, M.; Mrabet-Dahbi, S.; Werfel, T. Alpha-toxin is produced by skin colonizing Staphylococcus aureus and induces a T helper type 1 response in atopic dermatitis. Clin. Exp. Allergy 2005, 35, 1088–1095. [Google Scholar] [CrossRef] [PubMed]
- Berube, B.J.; Bubeck Wardenburg, J. Staphylococcus aureus α-toxin: Nearly a century of intrigue. Toxins 2013, 5, 1140–1166. [Google Scholar] [CrossRef] [PubMed]
- Moraveji, Z.; Tabatabaei, M.; Shirzad Aski, H.; Khoshbakht, R. Characterization of hemolysins of Staphylococcus strains isolated from human and bovine, southern Iran. Iran. J. Vet. Res. 2014, 15, 326–330. [Google Scholar] [PubMed]
- Huseby, M.; Shi, K.; Brown, C.K.; Digre, J.; Mengistu, F.; Seo, K.S.; Bohach, G.A.; Schlievert, P.M.; Ohlendorf, D.H.; Earhart, C.A. Structure and biological activities of beta toxin from Staphylococcus aureus. J. Bacteriol. 2007, 189, 8719–8726. [Google Scholar] [CrossRef]
- Novick, R.P. Autoinduction and signal transduction in the regulation of staphylococcal virulence. Mol. Microbiol. 2003, 48, 1429–1449. [Google Scholar] [CrossRef]
- Krzyminska, S.; Szczuka, E.; Kaznowski, A. Staphylococcus haemolyticus strains target mitochondria and induce caspase-dependent apoptosis of macrophages. Antonie Van Leeuwenhoek 2012, 102, 611–620. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Antibiotic | Antibiotic Action | Resistance Gene | Mechanism of Resistance | Reference |
|---|---|---|---|---|
| Methicillin | Inhibits bacterial peptidoglycan cross-linking through inhibition of transpeptidase enzyme |
|
| [70] |
| Glycopeptides (Vancomycin and teicoplanin) | Binds to the D-Ala-D-Ala leading to suppression of bacterial cell wall synthesis |
|
| [64,71] |
|
| |||
| Linezolid | Inhibits bacterial protein expression via interfering with the 23S rRNA in the ribosome |
|
| [42,72,73,74,75] |
| Lincosamides (lincomycin, clindamycin, Pirlimycin) | Interferes with bacterial protein formation through binding to the 23S/ 50s ribosome |
|
| [76,77] |
| [78] | ||||
| Mupirocin | Inhibits bacterial protein synthesis via targeting isoleucyl-tRNA synthetase |
|
| [79] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eltwisy, H.O.; Twisy, H.O.; Hafez, M.H.; Sayed, I.M.; El-Mokhtar, M.A. Clinical Infections, Antibiotic Resistance, and Pathogenesis of Staphylococcus haemolyticus. Microorganisms 2022, 10, 1130. https://doi.org/10.3390/microorganisms10061130
Eltwisy HO, Twisy HO, Hafez MH, Sayed IM, El-Mokhtar MA. Clinical Infections, Antibiotic Resistance, and Pathogenesis of Staphylococcus haemolyticus. Microorganisms. 2022; 10(6):1130. https://doi.org/10.3390/microorganisms10061130
Chicago/Turabian StyleEltwisy, Hala O., Howida Omar Twisy, Mahmoud HR Hafez, Ibrahim M. Sayed, and Mohamed A. El-Mokhtar. 2022. "Clinical Infections, Antibiotic Resistance, and Pathogenesis of Staphylococcus haemolyticus" Microorganisms 10, no. 6: 1130. https://doi.org/10.3390/microorganisms10061130
APA StyleEltwisy, H. O., Twisy, H. O., Hafez, M. H., Sayed, I. M., & El-Mokhtar, M. A. (2022). Clinical Infections, Antibiotic Resistance, and Pathogenesis of Staphylococcus haemolyticus. Microorganisms, 10(6), 1130. https://doi.org/10.3390/microorganisms10061130