
Synthetic Soil Aggregates: Bioprinted Habitats for High-Throughput Microbial Metaphenomics

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
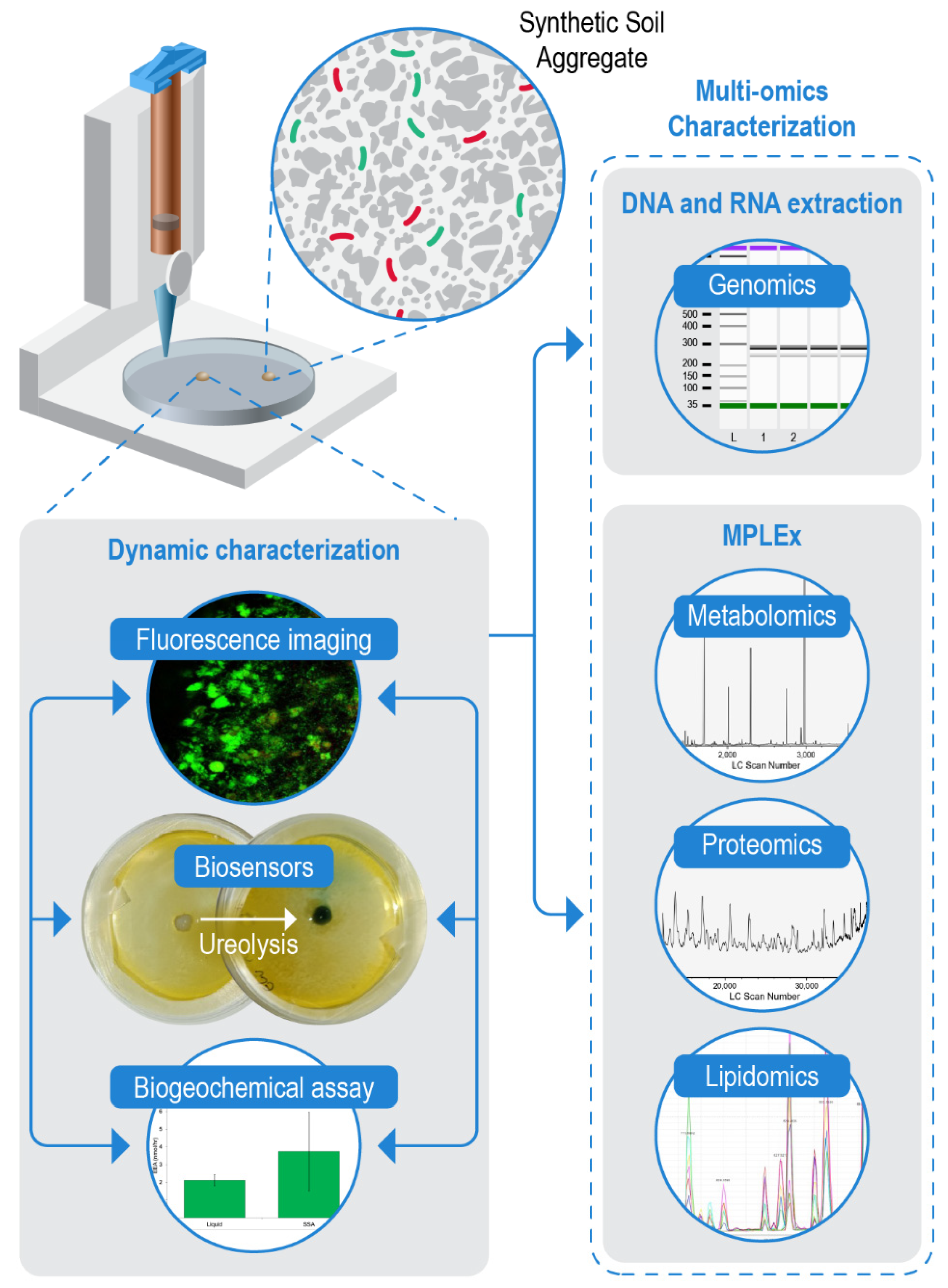
2.1. Bioprinting a Synthetic Soil Aggregate
2.2. Scanning Electron Microscopy
2.3. Pore Size Characterization
2.4. Diffusion of TRTIC-Dextran in GelMA C
2.5. Microbial Strains and Culture Conditions
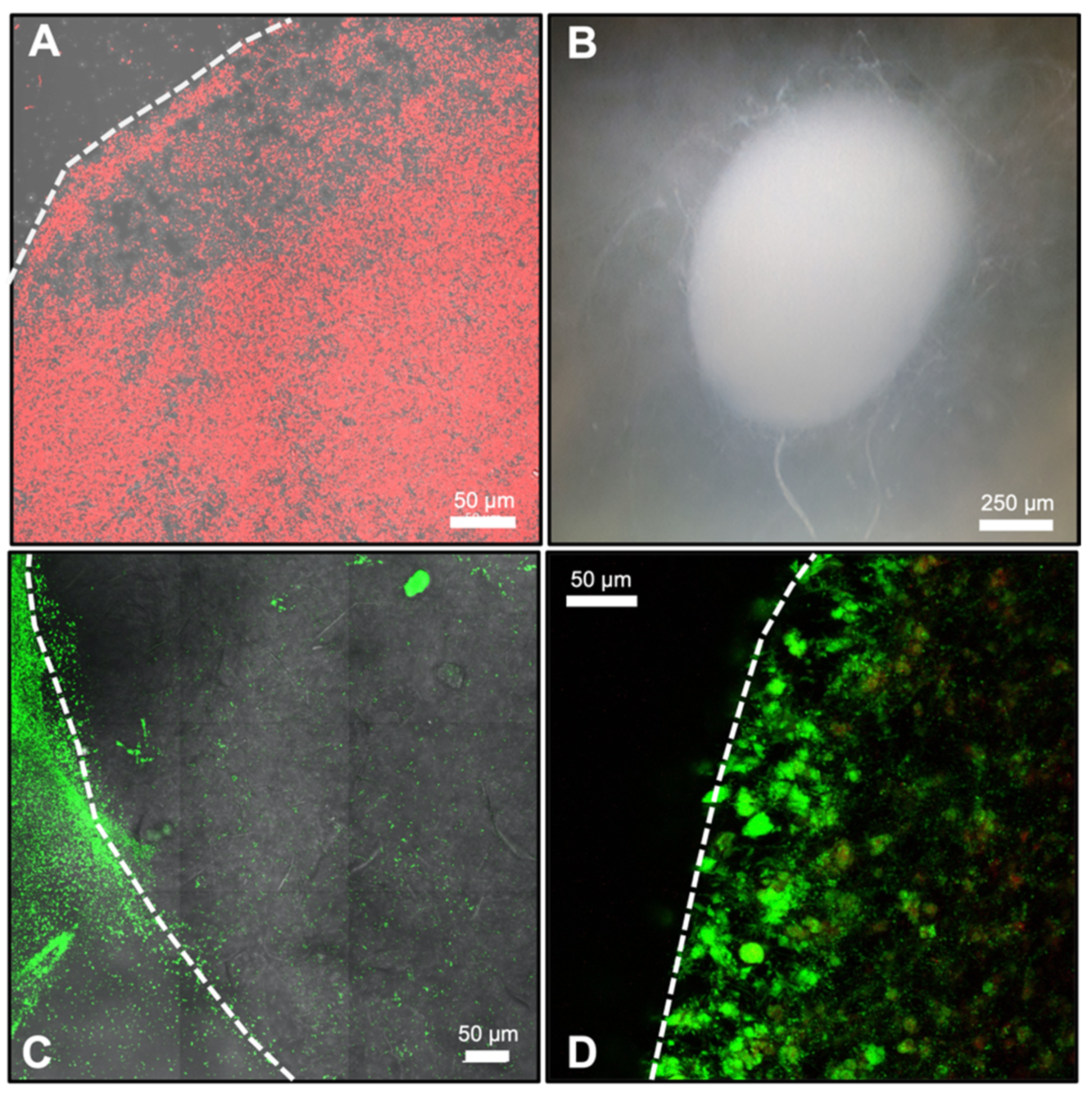
2.6. Validation of Microbial Viability within Synthetic Soil Aggregates
2.7. Multi-Omic Analysis of Synthetic Soil Aggregates
2.8. Biogeochemical Analyses in Synthetic Soil Aggregates
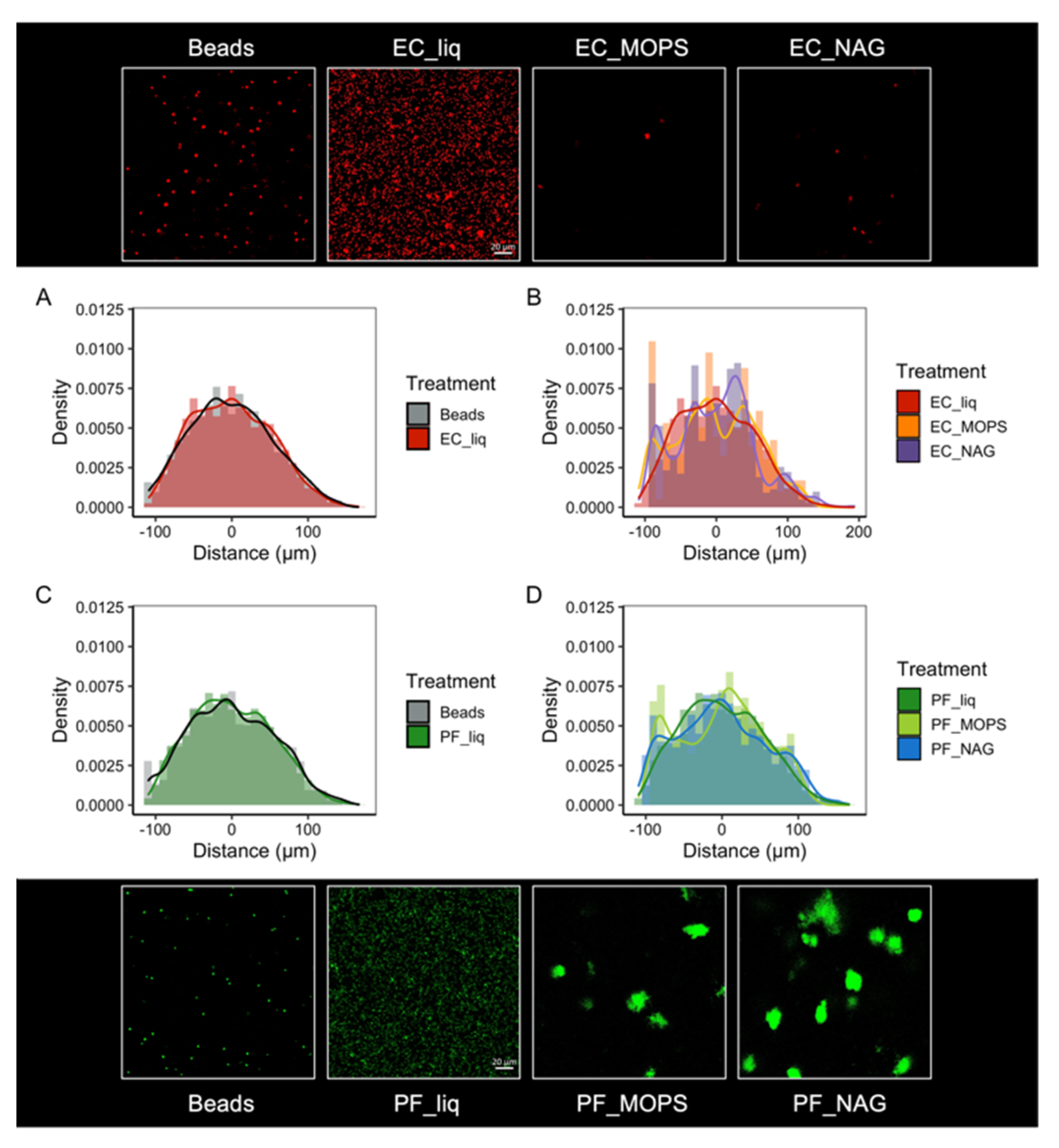
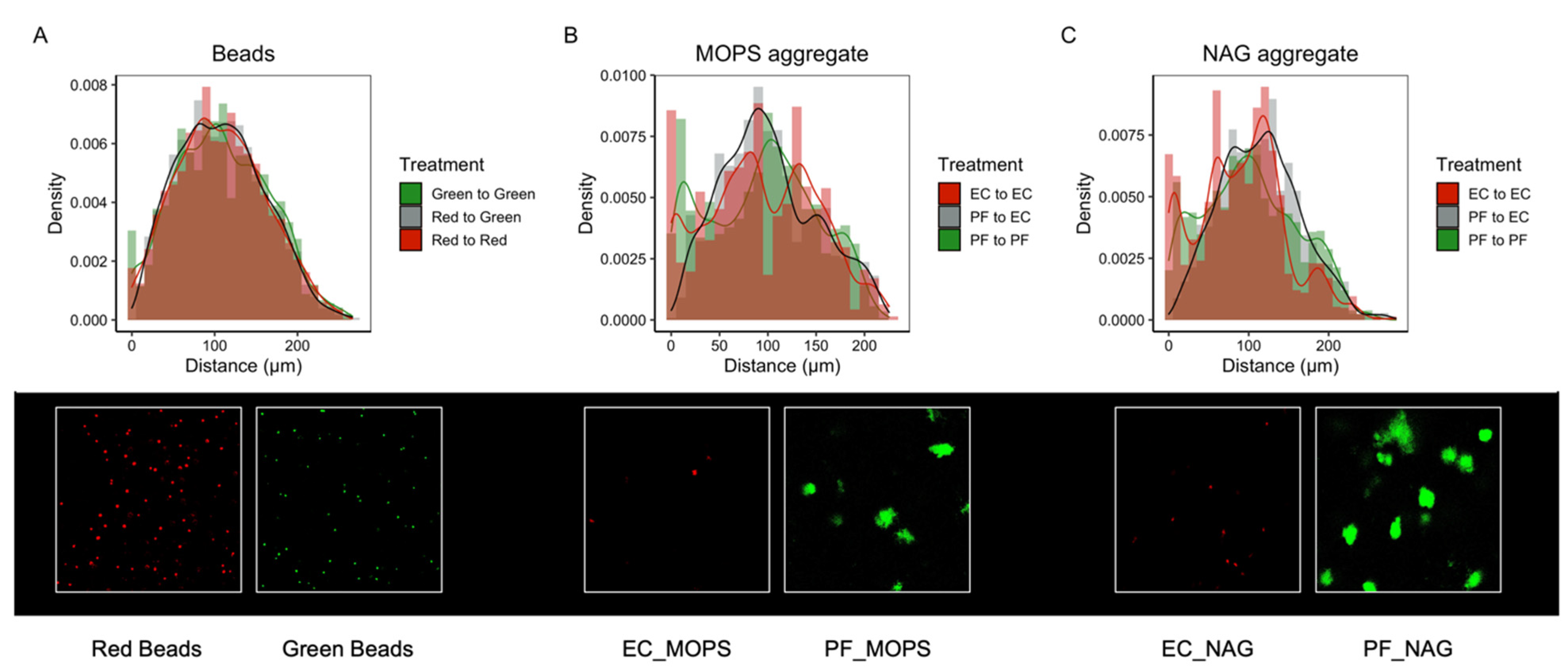
2.9. Spatial Organization of a Synthetic Community within SSAs
3. Results and Discussion
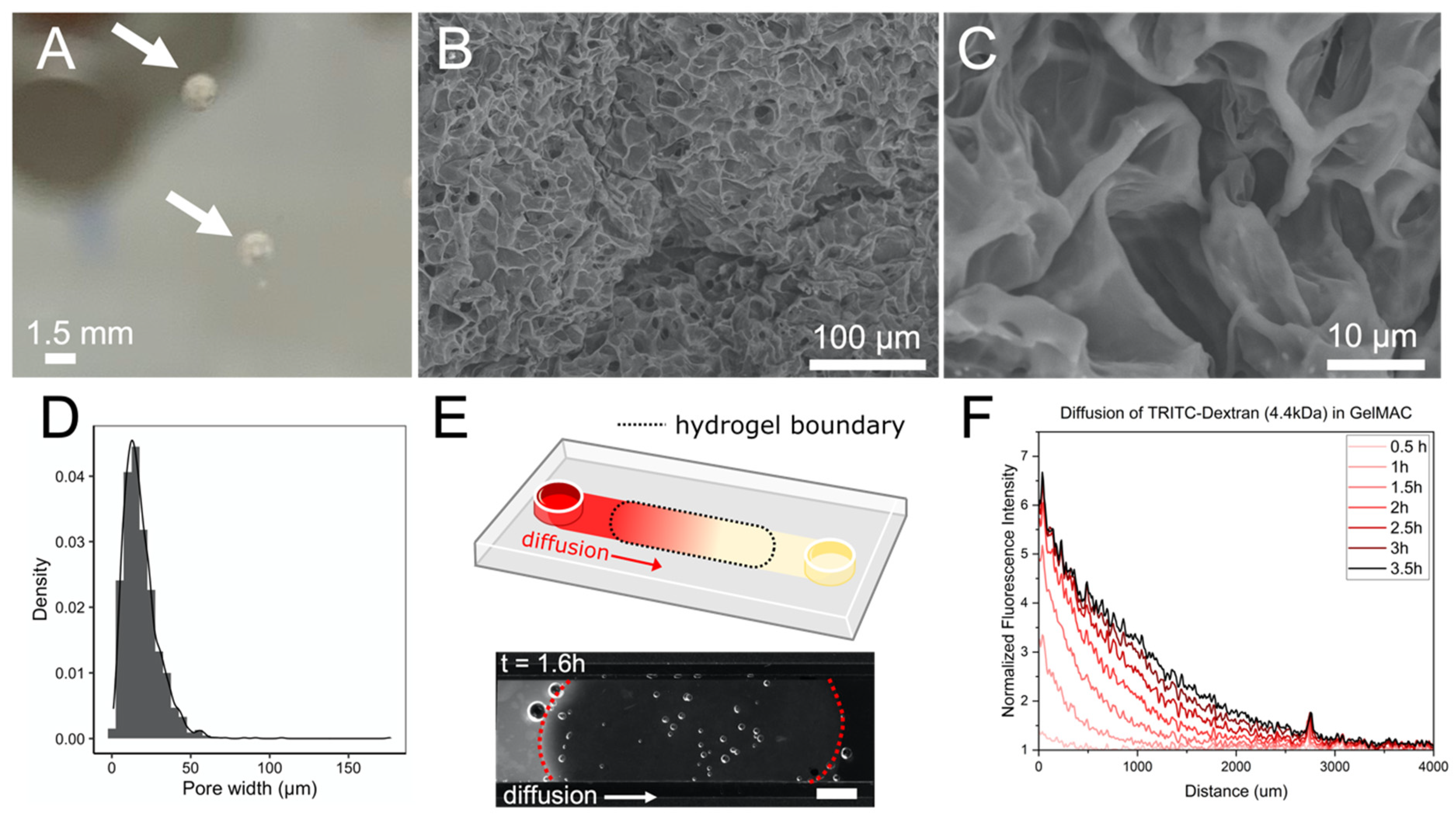
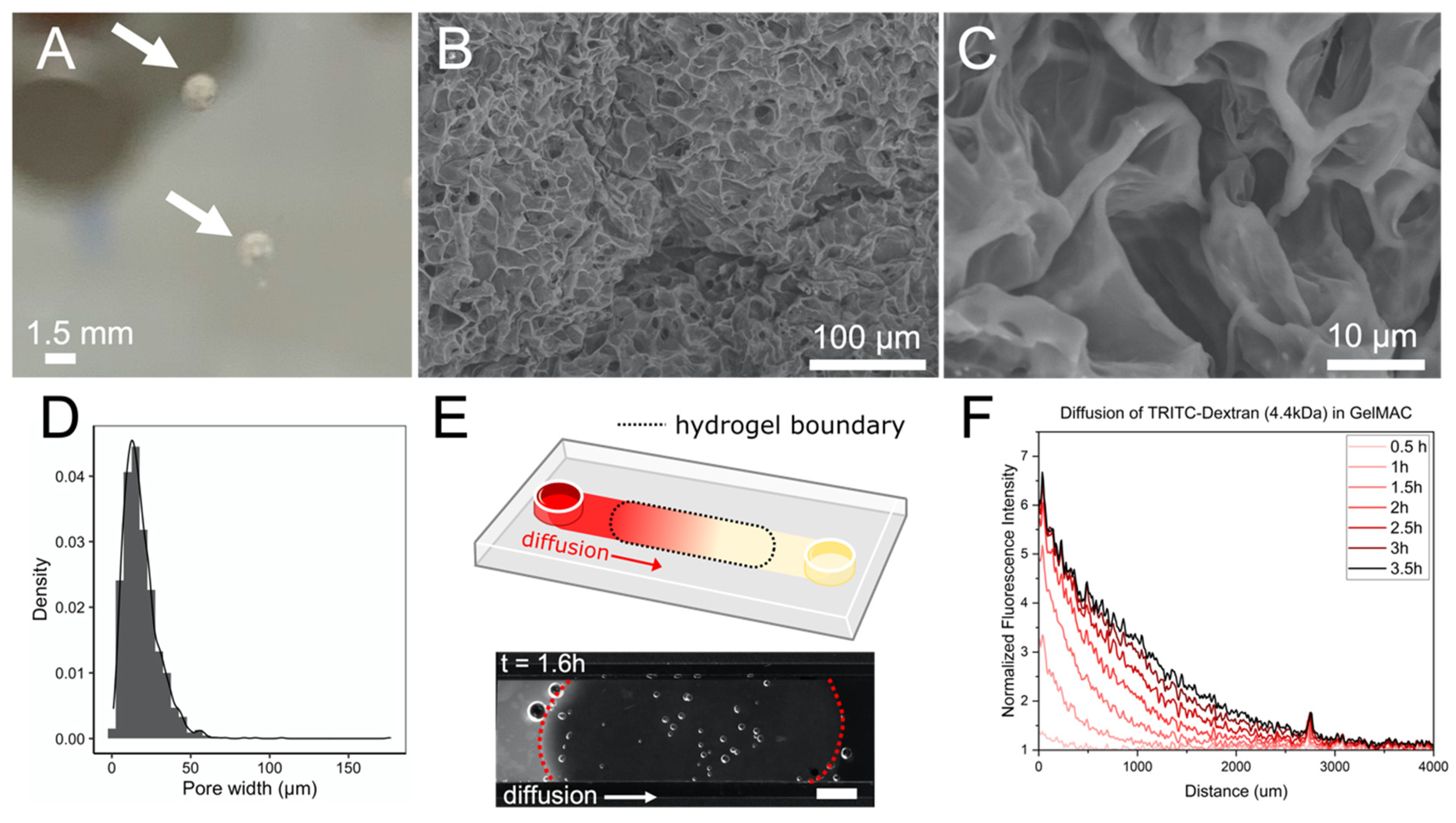
3.1. Characteristics of Synthetic Soil Aggregates
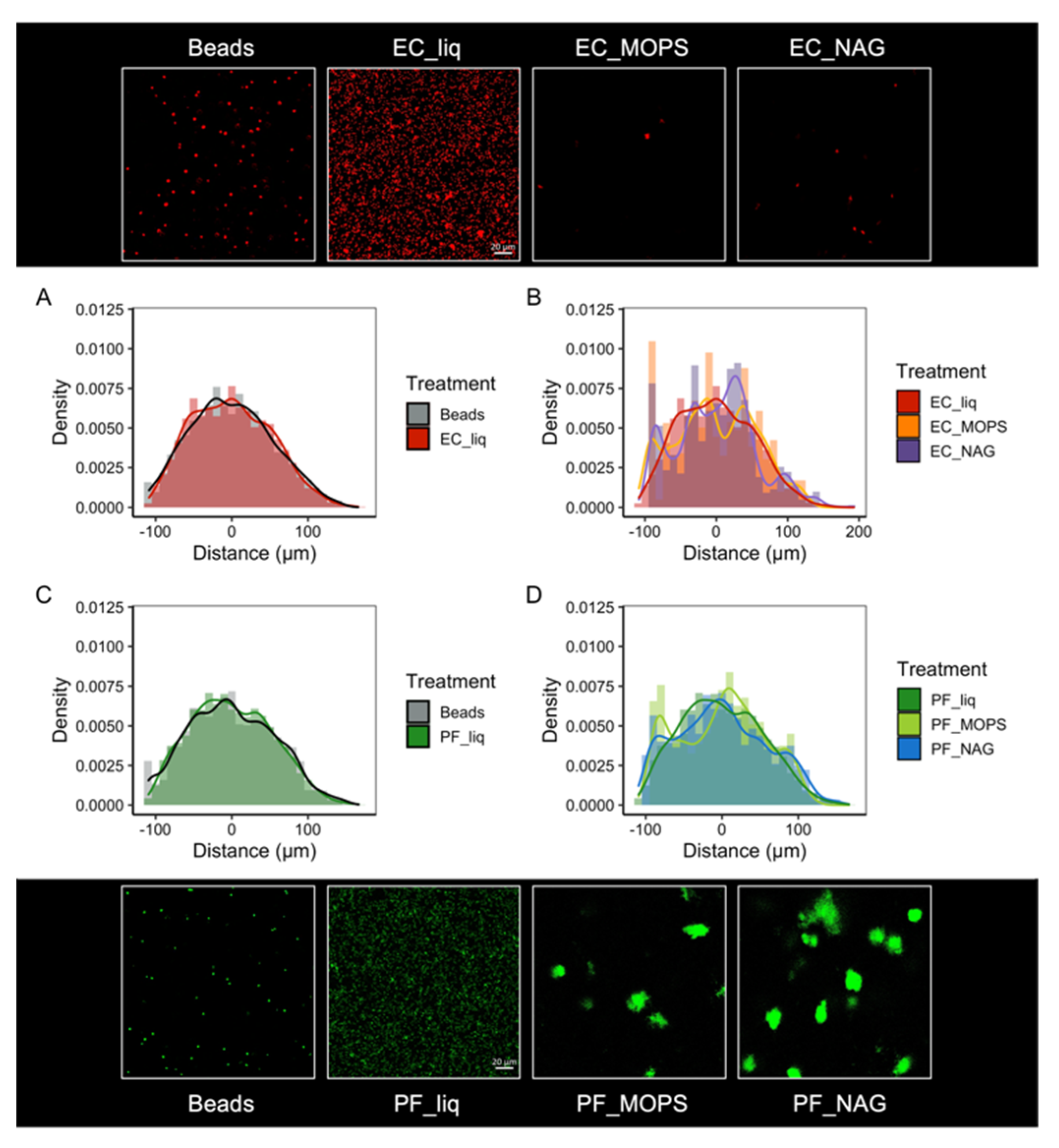
3.2. Microbial Viability within Synthetic Soil Aggregates
3.3. In Situ Metaphenomics
3.4. Biogeochemical Analyses and High-Throughput In Situ Assays
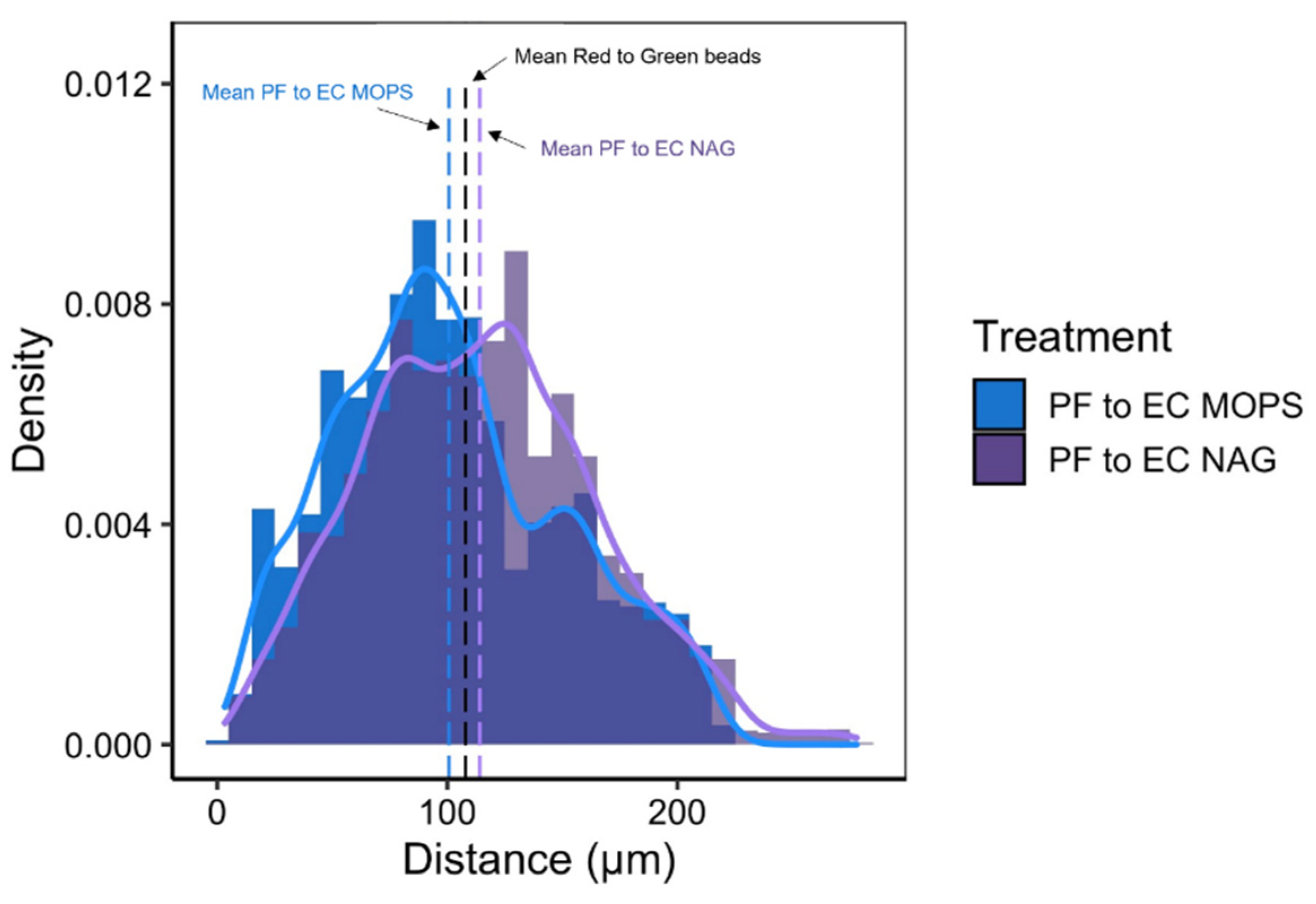
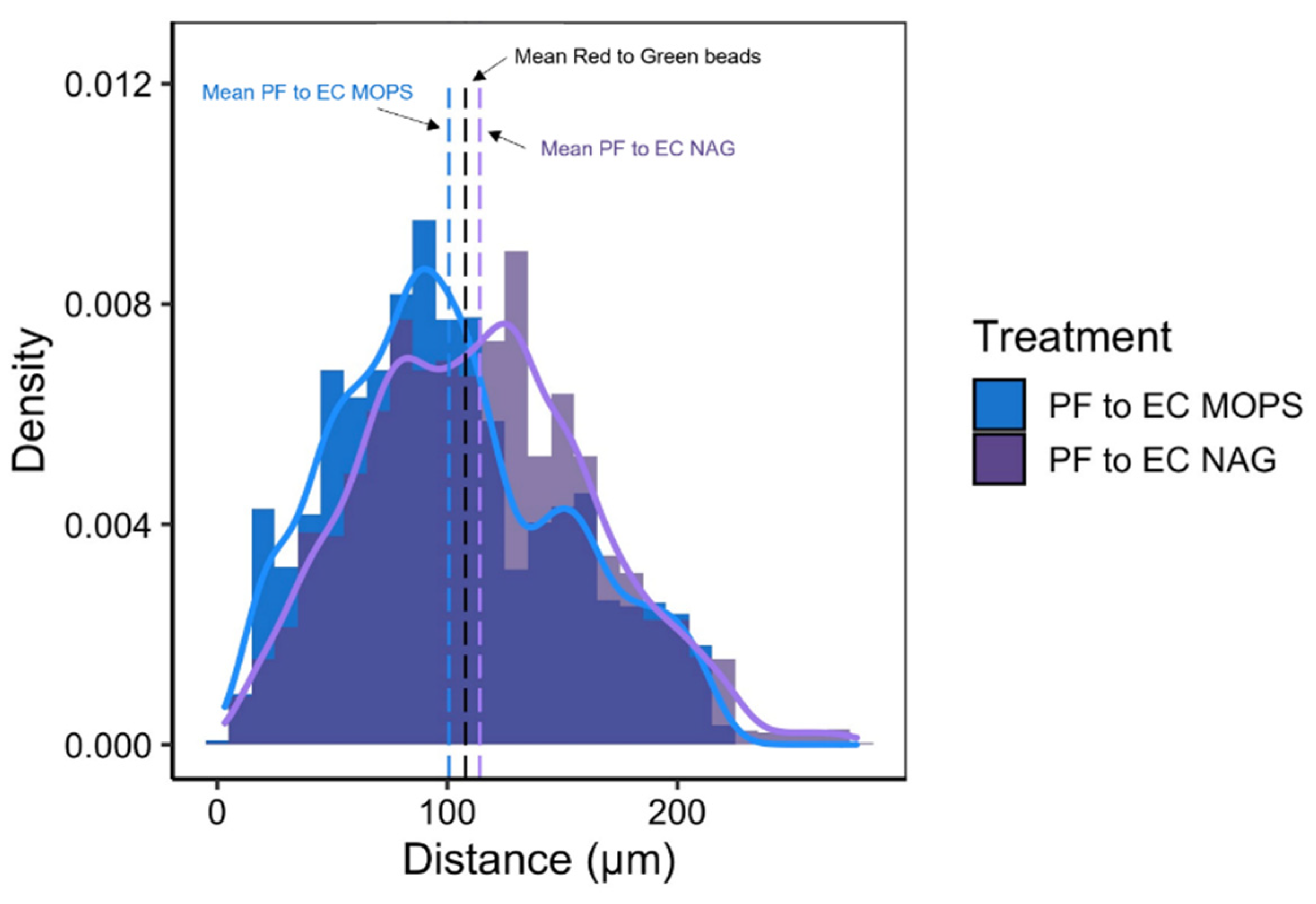
3.5. Synthetic Community Interactions in a Structured Environment
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Falkowski, P.G.; Fenchel, T.; Delong, E.F. The microbial engines that drive Earth’s biogeochemical cycles. Science 2008, 320, 1034–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansson, J.K.; Hofmockel, K.S. Soil microbiomes and climate change. Nat. Rev. Microbiol. 2020, 18, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Smercina, D.N.; Bailey, V.L.; Hofmockel, K.S. Micro on a macroscale: Relating microbial-scale soil processes to global ecosystem function. FEMS Microbiol. Ecol. 2021, 97, fiab091. [Google Scholar] [CrossRef] [PubMed]
- Hall, E.K.; Bernhardt, E.S.; Bier, R.L.; Bradford, M.A.; Boot, C.M.; Cotner, J.B.; del Giorgio, P.A.; Evans, S.E.; Graham, E.B.; Jones, S.E. Understanding how microbiomes influence the systems they inhabit. Nat. Microbiol. 2018, 3, 977–982. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Blagodatskaya, E. Microbial hotspots and hot moments in soil: Concept & review. Soil Biol. Biochem. 2015, 83, 184–199. [Google Scholar]
- Rillig, M.C.; Muller, L.A.; Lehmann, A. Soil aggregates as massively concurrent evolutionary incubators. ISME J. 2017, 11, 1943–1948. [Google Scholar] [CrossRef]
- Wang, B.; Brewer, P.E.; Shugart, H.H.; Lerdau, M.T.; Allison, S.D. Soil aggregates as biogeochemical reactors and implications for soil–atmosphere exchange of greenhouse gases—A concept. Glob. Chang. Biol. 2019, 25, 373–385. [Google Scholar] [CrossRef]
- Bach, E.M.; Hofmockel, K.S. Soil aggregate isolation method affects measures of intra-aggregate extracellular enzyme activity. Soil Biol. Biochem. 2014, 69, 54–62. [Google Scholar] [CrossRef]
- Bailey, V.L.; Bilskis, C.L.; Fansler, S.J.; McCue, L.A.; Smith, J.L.; Konopka, A. Measurements of microbial community activities in individual soil macroaggregates. Soil Biol. Biochem. 2012, 48, 192–195. [Google Scholar] [CrossRef]
- Bailey, V.L.; Fansler, S.J.; Stegen, J.C.; McCue, L.A. Linking microbial community structure to β-glucosidic function in soil aggregates. ISME J. 2013, 7, 2044–2053. [Google Scholar] [CrossRef]
- Wilpiszeski, R.L.; Aufrecht, J.A.; Retterer, S.T.; Sullivan, M.B.; Graham, D.E.; Pierce, E.M.; Zablocki, O.D.; Palumbo, A.V.; Elias, D.A. Soil aggregate microbial communities: Towards understanding microbiome interactions at biologically relevant scales. Appl. Environ. Microbiol. 2019, 85, e00319–e00324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.M.; Yeong, W.Y. Design and printing strategies in 3D bioprinting of cell–hydrogels: A review. Adv. Healthcare Mater. 2016, 5, 2856–2865. [Google Scholar] [CrossRef] [PubMed]
- Unagolla, J.M.; Jayasuriya, A.C. Hydrogel-based 3D bioprinting: A comprehensive review on cell-laden hydrogels, bioink formulations, and future perspectives. Appl. Mater. Today 2020, 18, 100479. [Google Scholar] [CrossRef] [PubMed]
- Legland, D.; Arganda-Carreras, I.; Andrey, P. MorphoLibJ: Integrated library and plugins for mathematical morphology with ImageJ. Bioinformatics 2016, 32, 3532–3534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Park, G.Y. Diffusion coefficient calculated by complementary error function for the sublimation diffusion of disperse dye. J. Eng. Fibers Fabr. 2019, 14, 1558925019866592. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Sadler, N.C.; Egbert, R.G.; Anderton, C.R.; Hofmockel, K.S.; Jansson, J.K.; Song, H.S. Deep learning predicts microbial interactions from self-organized spatiotemporal patterns. Comput. Struct. Biotechnol. J. 2020, 18, 1259–1269. [Google Scholar] [CrossRef]
- Zambare, N.M.; Naser, N.Y.; Gerlach, R.; Chang, C.B. Mineralogy of microbially induced calcium carbonate precipitates formed using single cell drop-based microfluidics. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Connolly, J.; Kaufman, M.; Rothman, A.; Gupta, R.; Redden, G.; Schuster, M.; Colwell, F.; Gerlach, R. Construction of two ureolytic model organisms for the study of microbially induced calcium carbonate precipitation. J. Microbiol. Methods 2013, 94, 290–299. [Google Scholar] [CrossRef] [Green Version]
- Ash, C.; Priest, F.G.; Collins, M.D. Molecular identification of rRNA group 3 bacilli (Ash, Farrow, Wallbanks and Collins) using a PCR probe test. Antonie Van Leeuwenhoek 1993, 64, 253–260. [Google Scholar] [CrossRef]
- Berepiki, A.; Lichius, A.; Shoji, J.-Y.; Tilsner, J.; Read, N.D. F-actin dynamics in Neurospora crassa. Eukaryot. Cell 2010, 9, 547–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayasu, E.S.; Nicora, C.D.; Sims, A.C.; Burnum-Johnson, K.E.; Kim, Y.M.; Kyle, J.E.; Matzke, M.M.; Shukla, A.K.; Chu, R.K.; Schepmoes, A.A. MPLEx: A robust and universal protocol for single-sample integrative proteomic, metabolomic, and lipidomic analyses. MSystems 2016, 1, e00043-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Qian, W.J.; Mottaz, H.M.; Clauss, T.R.; Anderson, D.J.; Moore, R.J.; Camp, D.G.; Khan, A.H.; Sforza, D.M.; Pallavicini, M. Development and evaluation of a micro-and nanoscale proteomic sample preparation method. J. Proteome Res. 2005, 4, 2397–2403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saiya-Cork, K.; Sinsabaugh, R.; Zak, D. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biol. Biochem. 2002, 34, 1309–1315. [Google Scholar] [CrossRef]
- Bailey, V.L.; McCue, L.A.; Fansler, S.J.; Boyanov, M.I.; DeCarlo, F.; Kemner, K.M.; Konopka, A. Micrometer-scale physical structure and microbial composition of soil macroaggregates. Soil Biol. Biochem. 2013, 65, 60–68. [Google Scholar] [CrossRef]
- Vos, M.; Wolf, A.B.; Jennings, S.J.; Kowalchuk, G.A. Micro-scale determinants of bacterial diversity in soil. FEMS Microbiol. Rev. 2013, 37, 936–954. [Google Scholar] [CrossRef] [Green Version]
- Stanley, N.R.; Lazazzera, B.A. Environmental signals and regulatory pathways that influence biofilm formation. Mol. Microbiol. 2004, 52, 917–924. [Google Scholar] [CrossRef]
- Kravchenko, A.N.; Guber, A.K. Soil pores and their contributions to soil carbon processes. Geoderma 2017, 287, 31–39. [Google Scholar] [CrossRef]
- Kravchenko, A.N.; Guber, A.K.; Razavi, B.S.; Koestel, J.; Quigley, M.Y.; Robertson, G.P.; Kuzyakov, Y. Microbial spatial footprint as a driver of soil carbon stabilization. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- van Gestel, J.; Bareia, T.; Tenennbaum, B.; Dal Co, A.; Guler, P.; Aframian, N.; Puyesky, S.; Grinberg, I.; D’Souza, G.G.; Erez, Z. Short-range quorum sensing controls horizontal gene transfer at micron scale in bacterial communities. Nat. Commun. 2021, 12, 1–11. [Google Scholar] [CrossRef]
- Hart, T.; Chamberlain, A.; Lynch, J.; Newling, B.; McDonald, P. A stray field magnetic resonance study of water diffusion in bacterial exopolysaccharides. Enzym. Microb. Technol. 1999, 24, 339–347. [Google Scholar] [CrossRef]
- Olesen, T.; Moldrup, P.; Gamst, J. Solute diffusion and adsorption in six soils along a soil texture gradient. Soil Sci. Soc. Am. J. 1999, 63, 519–524. [Google Scholar] [CrossRef]
- Yu, J.S.; Correia-Melo, C.; Zorrilla, F.; Herrera-Dominguez, L.; Wu, M.Y.; Hartl, J.; Campbell, K.; Blasche, S.; Kreidl, M.; Egger, A.-S. Microbial communities form rich extracellular metabolomes that foster metabolic interactions and promote drug tolerance. Nat. Microbiol. 2022, 58, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Aleklett, K.; Kiers, E.T.; Ohlsson, P.; Shimizu, T.S.; Caldas, V.E.; Hammer, E.C. Build your own soil: Exploring microfluidics to create microbial habitat structures. ISME J. 2018, 12, 312–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aufrecht, J.A.; Fowlkes, J.D.; Bible, A.N.; Morrell-Falvey, J.; Doktycz, M.J.; Retterer, S.T. Pore-scale hydrodynamics influence the spatial evolution of bacterial biofilms in a microfluidic porous network. PLoS ONE 2019, 14, e0218316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanley, C.E.; Grossmann, G.; i Solvas, X.C.; deMello, A.J. Soil-on-a-Chip: Microfluidic platforms for environmental organismal studies. Lab Chip 2016, 16, 228–241. [Google Scholar] [CrossRef]
- Deng, J.; Orner, E.P.; Chau, J.F.; Anderson, E.M.; Kadilak, A.L.; Rubinstein, R.L.; Bouchillon, G.M.; Goodwin, R.A.; Gage, D.J.; Shor, L.M. Synergistic effects of soil microstructure and bacterial EPS on drying rate in emulated soil micromodels. Soil Biol. Biochem. 2015, 83, 116–124. [Google Scholar] [CrossRef]
- Gladieux, P.; Bellis, F.D.; Hann-Soden, C.; Svedberg, J.; Johannesson, H.; Taylor, J.W. Neurospora from natural populations: Population genomics insights into the life history of a model microbial eukaryote. Stat. Popul. Genom. 2020, 14, 313–336. [Google Scholar]
- Jousset, A.; Jousset, A.; Bienhold, C.; Chatzinotas, A.; Gallien, L.; Gobet, A.; Kurm, V.; Küsel, K.; Rillig, M.C.; Rivett, D.W.; et al. Where less may be more: How the rare biosphere pulls ecosystems strings. ISME J. 2017, 11, 853–862. [Google Scholar] [CrossRef]
- Jansson, J.K.; Hofmockel, K.S. The soil microbiome—from metagenomics to metaphenomics. Curr. Opin. Microbiol. 2018, 43, 162–168. [Google Scholar] [CrossRef]
- Carini, P.; Marsden, P.J.; Leff, J.W.; Morgan, E.E.; Strickland, M.S.; Fierer, N. Relic DNA is abundant in soil and obscures estimates of soil microbial diversity. Nat. Microbiol. 2016, 2, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Upton, R.N.; Bach, E.M.; Hofmockel, K.S. Spatio-temporal microbial community dynamics within soil aggregates. Soil Biol. Biochem. 2019, 132, 58–68. [Google Scholar] [CrossRef]
- Graham, E.B.; Crump, A.R.; Kennedy, D.W.; Arntzen, E.; Fansler, S.; Purvine, S.O.; Nicora, C.D.; Nelson, W.; Tfaily, M.M.; Stegen, J.C. Multi’omics comparison reveals metabolome biochemistry, not microbiome composition or gene expression, corresponds to elevated biogeochemical function in the hyporheic zone. Sci. Total Environ. 2018, 642, 742–753. [Google Scholar] [CrossRef] [PubMed]
- Nicora, C.D.; Burnum-Johnson, K.E.; Nakayasu, E.S.; Casey, C.P.; White, R.A.; Chowdhury, T.R.; Kyle, J.E.; Kim, Y.M.; Smith, R.D.; Metz, T.O. The MPLEx protocol for multi-omic analyses of soil samples. JoVE J. Vis. Exp. 2018, 145, e57343. [Google Scholar] [CrossRef] [PubMed]
- Roy Chowdhury, T.; Lee, J.Y.; Bottos, E.M.; Brislawn, C.J.; White, R.A.; Bramer, L.M.; Brown, J.; Zucker, J.D.; Kim, Y.M.; Jumpponen, A. Metaphenomic responses of a native prairie soil microbiome to moisture perturbations. Msystems 2019, 4, e00061-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starke, R.; Mondéjar, R.L.; Human, Z.R.; Navrátilová, D.; Štursová, M.; Větrovský, T.; Olson, H.M.; Orton, D.J.; Callister, S.J.; Lipton, M.S. Niche differentiation of bacteria and fungi in carbon and nitrogen cycling of different habitats in a temperate coniferous forest: A metaproteomic approach. Soil Biol. Biochem. 2021, 155, 108170. [Google Scholar] [CrossRef]
- Graham, E.B.; Knelman, J.E.; Schindlbacher, A.; Siciliano, S.; Breulmann, M.; Yannarell, A.; Beman, J.; Abell, G.; Philippot, L.; Prosser, J. Microbes as engines of ecosystem function: When does community structure enhance predictions of ecosystem processes? Front. Microbiol. 2016, 7, 214. [Google Scholar] [CrossRef] [Green Version]
- Weintraub, M.N.; Scott-Denton, L.E.; Schmidt, S.K.; Monson, R.K. The effects of tree rhizodeposition on soil exoenzyme activity, dissolved organic carbon, and nutrient availability in a subalpine forest ecosystem. Oecologia 2007, 154, 327–338. [Google Scholar] [CrossRef]
- Fansler, S.J.; Smith, J.L.; Bolton, H.; Bailey, V.L. Distribution of two C cycle enzymes in soil aggregates of a prairie chronosequence. Biol. Fertil. Soils 2005, 42, 17–23. [Google Scholar] [CrossRef]
- German, D.P.; Weintraub, M.N.; Grandy, A.S.; Lauber, C.L.; Rinkes, Z.L.; Allison, S.D. Optimization of hydrolytic and oxidative enzyme methods for ecosystem studies. Soil Biol. Biochem. 2011, 43, 1387–1397. [Google Scholar] [CrossRef]
- Zeng, N.; Wu, Y.; Chen, W.; Huang, Q.; Cai, P. Whole-cell microbial bioreporter for soil contaminants detection. Front. Bioeng. Biotechnol. 2021, 9, 622994. [Google Scholar] [CrossRef] [PubMed]
- Borer, B.; Tecon, R.; Or, D. Spatial organization of bacterial populations in response to oxygen and carbon counter-gradients in pore networks. Nat. Commun. 2018, 9, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miri, A.; Hosseinabadi, H.; Cecen, B.; Hassan, S.; Zhang, Y. Permeability mapping of gelatin methacryloyl hydrogels. Acta Biomater. 2018, 77, 38–47. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | Validation Method | Optimal Growth Period |
|---|---|---|
| Escherichia coli |
| 48 h |
| Pseudomonas fluorescens | Confocal microscopy—Green fluorescent protein | 72 h |
| Neurospora crassa | Confocal microscopy—tdTomato fluorescent protein | <24 h |
| Escherichia coli (urea hydrolyzing) |
| 24 h |
| Paenibacilluspolymyxa | Confocal microscopy—Live/dead stain | 72 h |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smercina, D.; Zambare, N.; Hofmockel, K.; Sadler, N.; Bredeweg, E.L.; Nicora, C.; Markillie, L.M.; Aufrecht, J. Synthetic Soil Aggregates: Bioprinted Habitats for High-Throughput Microbial Metaphenomics. Microorganisms 2022, 10, 944. https://doi.org/10.3390/microorganisms10050944
Smercina D, Zambare N, Hofmockel K, Sadler N, Bredeweg EL, Nicora C, Markillie LM, Aufrecht J. Synthetic Soil Aggregates: Bioprinted Habitats for High-Throughput Microbial Metaphenomics. Microorganisms. 2022; 10(5):944. https://doi.org/10.3390/microorganisms10050944
Chicago/Turabian StyleSmercina, Darian, Neerja Zambare, Kirsten Hofmockel, Natalie Sadler, Erin L. Bredeweg, Carrie Nicora, Lye Meng Markillie, and Jayde Aufrecht. 2022. "Synthetic Soil Aggregates: Bioprinted Habitats for High-Throughput Microbial Metaphenomics" Microorganisms 10, no. 5: 944. https://doi.org/10.3390/microorganisms10050944
APA StyleSmercina, D., Zambare, N., Hofmockel, K., Sadler, N., Bredeweg, E. L., Nicora, C., Markillie, L. M., & Aufrecht, J. (2022). Synthetic Soil Aggregates: Bioprinted Habitats for High-Throughput Microbial Metaphenomics. Microorganisms, 10(5), 944. https://doi.org/10.3390/microorganisms10050944