
Essential Oils Biofilm Modulation Activity and Machine Learning Analysis on Pseudomonas aeruginosa Isolates from Cystic Fibrosis Patients


 , ,
, ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Approval and Informed Consent
2.2. Description of P. aeruginosa Clinical Isolates from CF Patients
2.3. Biofilm Production Assay in the Presence of EO
2.4. Essential Oil Chemical Composition Analysis
2.5. Machine Learning Binary Classification Modeling
- A first coarse ML model generation was run with 10 random hyperparameter combination runs from all possible considered combinations (Tables S5 and S6) [28];
- A second level of investigation was run with 100 random hyperparameter combination runs from all possible considered combinations (Tables S6 and S7) to select the optimal DA settings;
- A pre-final level was run with 1000 random hyperparameter combinations to check for protocol correctness, while extracting statistical coefficients for preliminary model evaluation;
- A final hyperparameter combination selection was performed by running 10,000 random combinations;
- The best model was finally further investigated with 1000 runs of DA perturbations, and the top scored model was used to deeply analyze the data.
3. Results
3.1. Biofilm Production Modulation by EOs
3.2. Essential Oil Chemical Composition
3.3. Machine Learning Models
3.3.1. Datasets
3.3.2. Classification Models
3.3.3. Chemical Components Importance and Partial Dependences
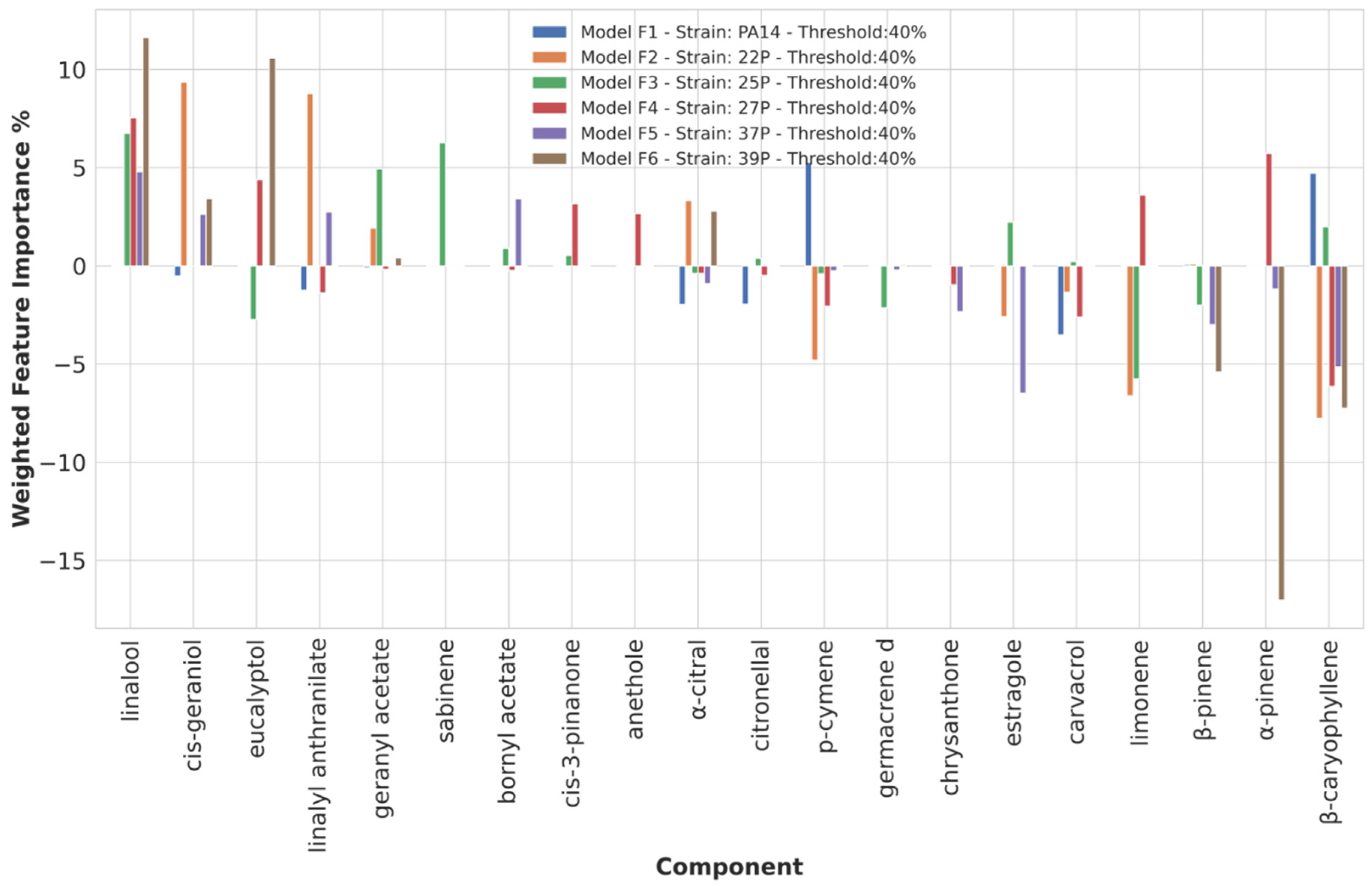
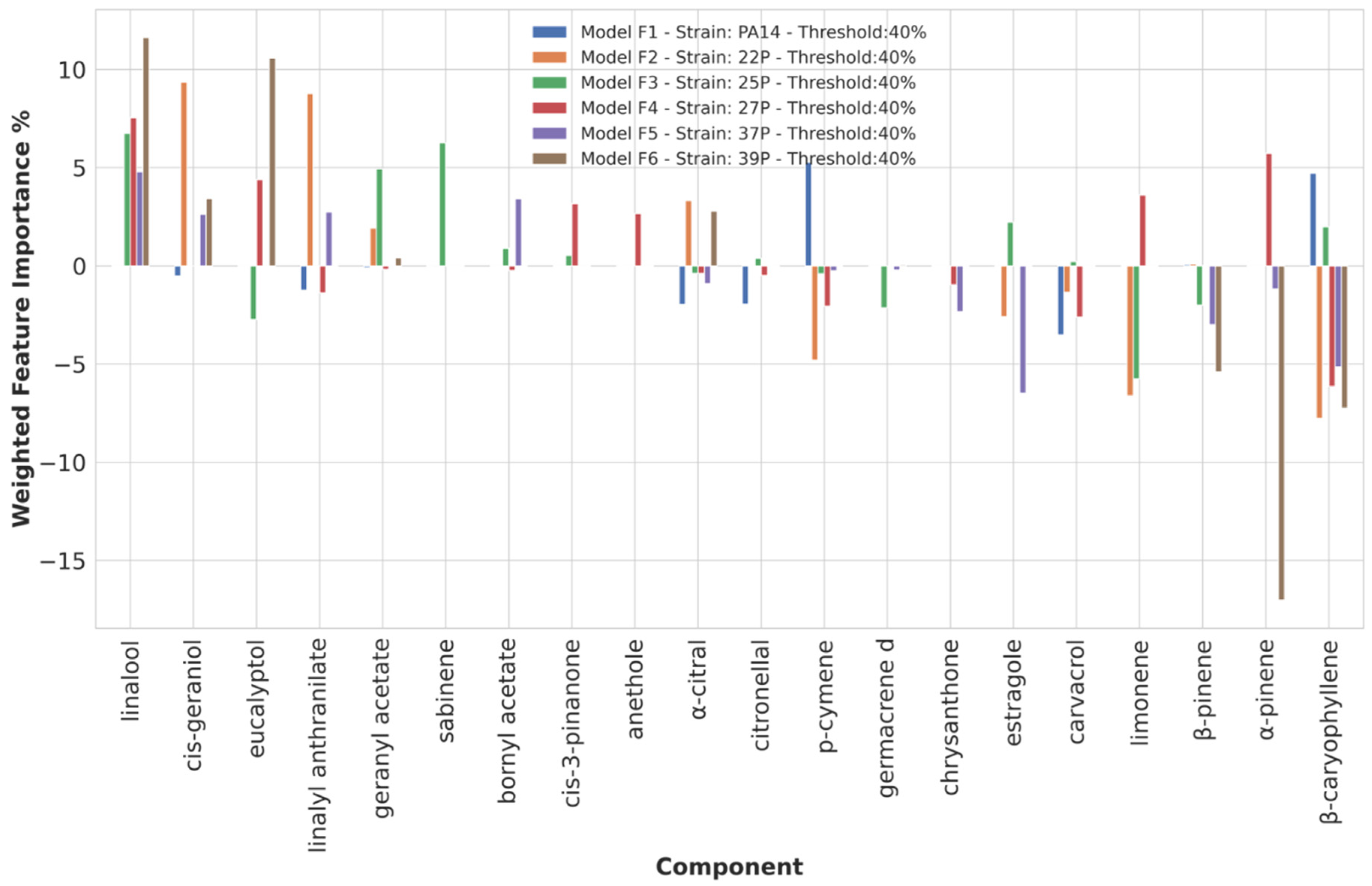
3.3.4. Chemical Components Importance and Partial Dependences at 40% Biofilm Production Threshold Value
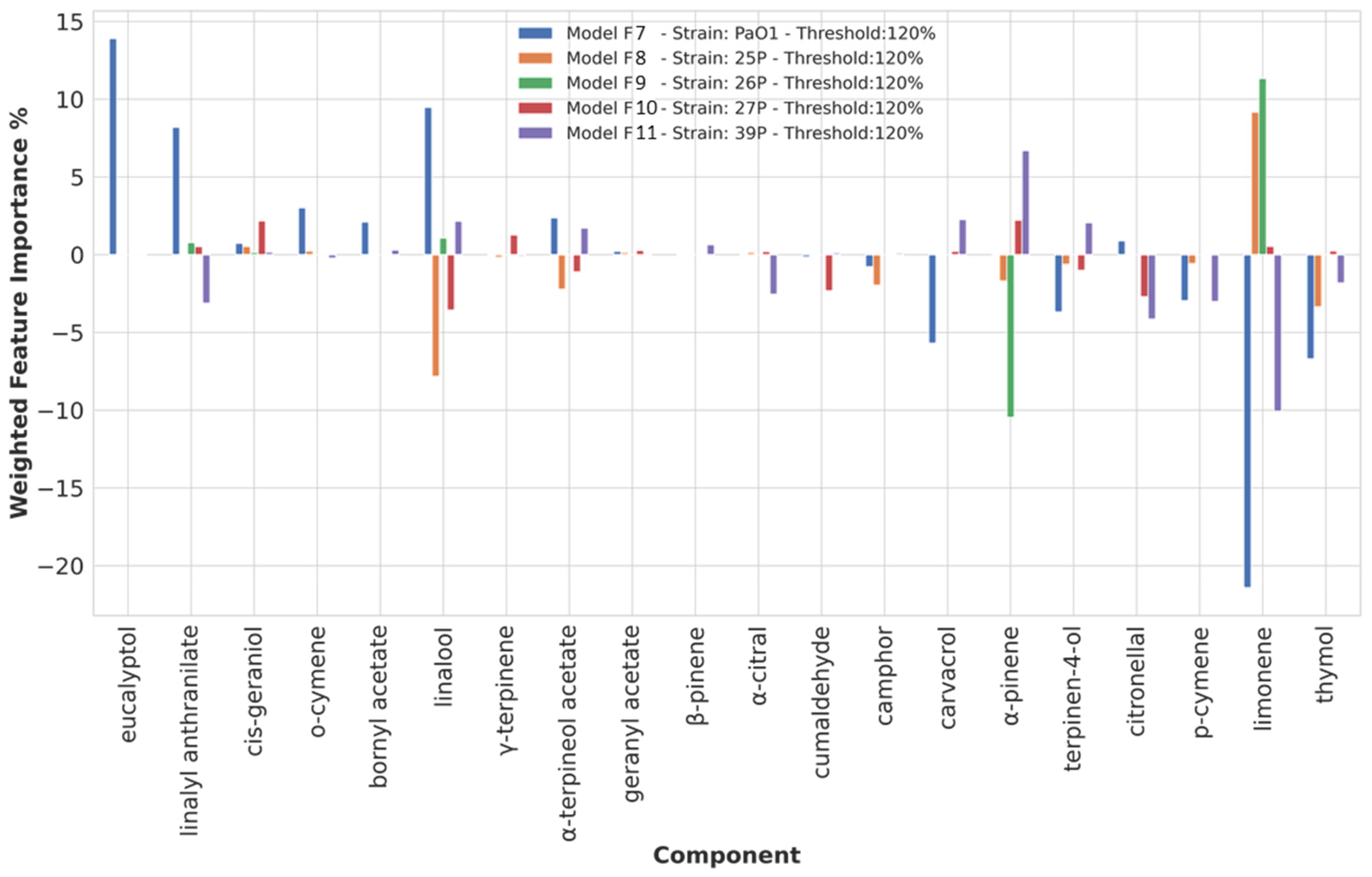
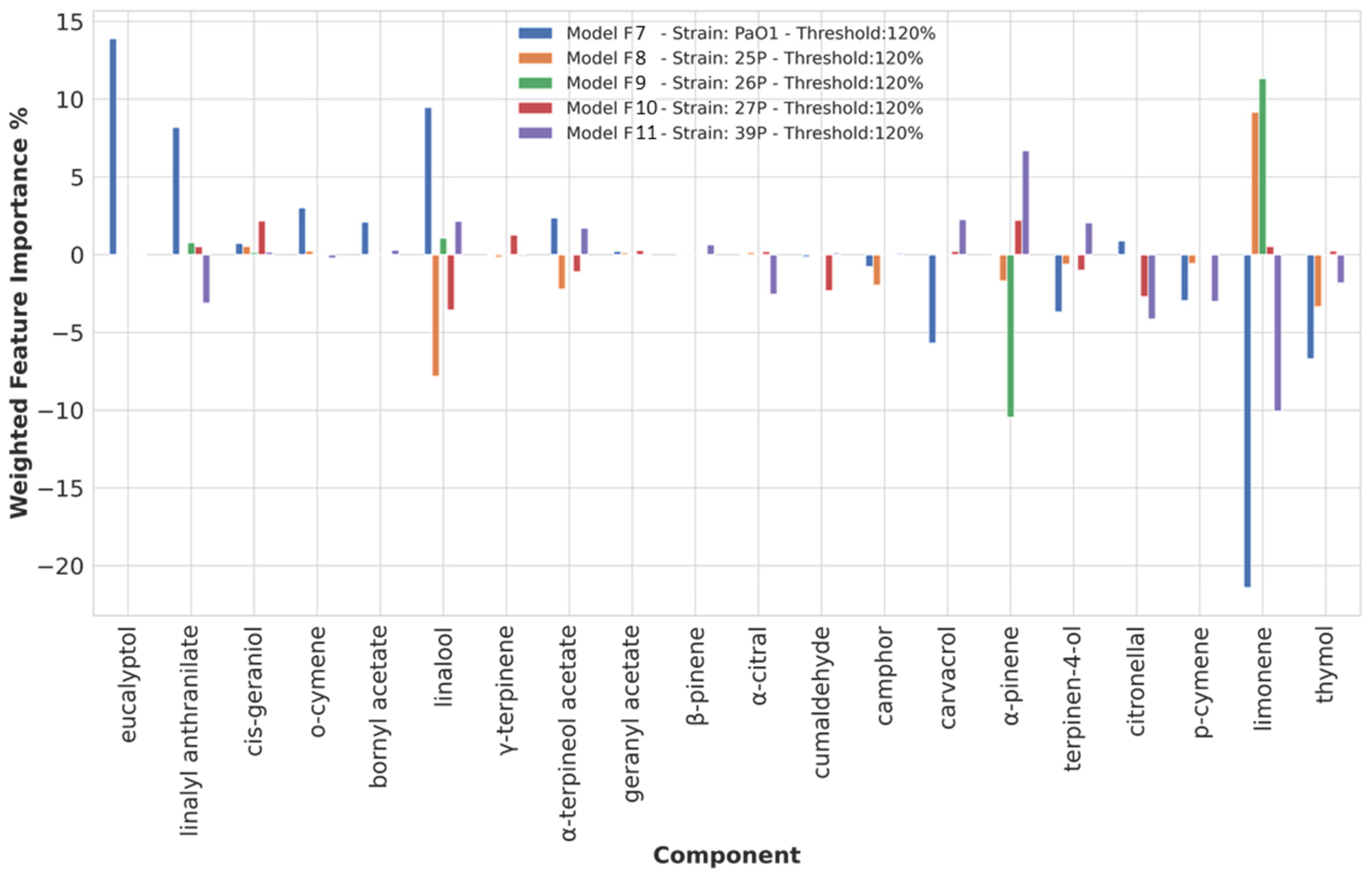
3.3.5. Chemical Components Importance and Partial Dependences at a 120% Biofilm Production Threshold Value
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Jurado-Martin, I.; Sainz-Mejias, M.; McClean, S. Pseudomonas aeruginosa: An audacious pathogen with an adaptable arsenal of virulence factors. Int. J. Mol. Sci. 2021, 22, 3128. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Barat, L.; Ferrer, M.; De Rosa, F.; Gabarrus, A.; Esperatti, M.; Terraneo, S.; Rinaudo, M.; Li Bassi, G.; Torres, A. Intensive care unit-acquired pneumonia due to Pseudomonas aeruginosa with and without multidrug resistance. J. Infect. 2017, 74, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Nunez, M.; Marti, S.; Puig, C.; Perez-Brocal, V.; Millares, L.; Santos, S.; Ardanuy, C.; Moya, A.; Linares, J.; Monso, E. Bronchial microbiome, PA biofilm-forming capacity and exacerbation in severe COPD patients colonized by P. aeruginosa. Future Microbiol. 2017, 12, 379–392. [Google Scholar] [CrossRef]
- Winstanley, C.; O’Brien, S.; Brockhurst, M.A. Pseudomonas aeruginosa evolutionary adaptation and diversification in cystic fibrosis chronic lung infections. Trends Microbiol. 2016, 24, 327–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Riquelme, S.A.; Ahn, D.; Prince, A. Pseudomonas aeruginosa and Klebsiella pneumoniae adaptation to innate immune clearance mechanisms in the lung. J. Innate Immun. 2018, 10, 442–454. [Google Scholar] [CrossRef]
- Hubble, V.B.; Hubbard, B.A.; Minrovic, B.M.; Melander, R.J.; Melander, C. Using small-molecule adjuvants to repurpose azithromycin for use against Pseudomonas aeruginosa. ACS Infect. Dis. 2019, 5, 141–151. [Google Scholar] [CrossRef]
- Lee, K.; Yoon, S.S. Pseudomonas aeruginosa biofilm, a programmed bacterial life for fitness. J. Microbiol. Biotechnol. 2017, 27, 1053–1064. [Google Scholar] [CrossRef] [Green Version]
- Yan, S.; Wu, G. Can biofilm be reversed through quorum sensing in Pseudomonas aeruginosa? Front. Microbiol. 2019, 10, 1582. [Google Scholar] [CrossRef] [Green Version]
- Martinez, M.; Goncalves, S.; Felicio, M.R.; Maturana, P.; Santos, N.C.; Semorile, L.; Hollmann, A.; Maffia, P.C. Synergistic and antibiofilm activity of the antimicrobial peptide P5 against carbapenem-resistant Pseudomonas aeruginosa. Biochim. Biophys. Acta Biomembr. 2019, 1861, 1329–1337. [Google Scholar] [CrossRef]
- Nair, S.; Desai, S.; Poonacha, N.; Vipra, A.; Sharma, U. Antibiofilm activity and synergistic inhibition of Staphylococcus aureus biofilms by bactericidal protein P128 in combination with antibiotics. Antimicrob. Agents Chemother. 2016, 60, 7280–7289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijesinghe, G.K.; Feiria, S.B.; Maia, F.C.; Oliveira, T.R.; Joia, F.; Barbosa, J.P.; Boni, G.C.; Höfling, J.F. In-vitro antibacterial and antibiofilm activity of cinnamomum verum leaf oil against Pseudomonas aeruginosa, Staphylococcus aureus and Klebsiella pneumoniae. An. da Acad. Bras. de Ciências 2021, 93, e20201507. [Google Scholar] [CrossRef] [PubMed]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Solorzano-Santos, F.; Miranda-Novales, M.G. Essential oils from aromatic herbs as antimicrobial agents. Curr. Opin. Biotechnol. 2012, 23, 136–141. [Google Scholar] [CrossRef]
- Artini, M.; Patsilinakos, A.; Papa, R.; Bozovic, M.; Sabatino, M.; Garzoli, S.; Vrenna, G.; Tilotta, M.; Pepi, F.; Ragno, R.; et al. Antimicrobial and antibiofilm activity and machine learning classification analysis of essential oils from different Mediterranean plants against Pseudomonas aeruginosa. Molecules 2018, 23, 482. [Google Scholar] [CrossRef] [Green Version]
- Papa, R.; Garzoli, S.; Vrenna, G.; Sabatino, M.; Sapienza, F.; Relucenti, M.; Donfrancesco, O.; Fiscarelli, E.V.; Artini, M.; Selan, L.; et al. Essential oils biofilm modulation activity, chemical and machine learning analysis. application on Staphylococcus aureus isolates from cystic fibrosis patients. Int. J. Mol. Sci. 2020, 21, 9258. [Google Scholar] [CrossRef]
- Ragno, R.; Papa, R.; Patsilinakos, A.; Vrenna, G.; Garzoli, S.; Tuccio, V.; Fiscarelli, E.; Selan, L.; Artini, M. Essential oils against bacterial isolates from cystic fibrosis patients by means of antimicrobial and unsupervised machine learning approaches. Sci. Rep. 2020, 10, 2653. [Google Scholar] [CrossRef]
- Kerem, E.; Conway, S.; Elborn, S.; Heijerman, H.; Consensus, C. Standards of care for patients with cystic fibrosis: A European consensus. J. Cyst. Fibros. 2005, 4, 7–26. [Google Scholar] [CrossRef] [Green Version]
- Mikkelsen, H.; McMullan, R.; Filloux, A. The Pseudomonas aeruginosa reference strain PA14 displays increased virulence due to a mutation in ladS. PLoS ONE 2011, 6, e29113. [Google Scholar] [CrossRef] [Green Version]
- Patsilinakos, A.; Artini, M.; Papa, R.; Sabatino, M.; Bozovic, M.; Garzoli, S.; Vrenna, G.; Buzzi, R.; Manfredini, S.; Selan, L.; et al. Machine Learning analyses on data including essential oil chemical composition and in vitro experimental antibiofilm activities against Staphylococcus species. Molecules 2019, 24, 890. [Google Scholar] [CrossRef] [Green Version]
- McKinney, W. Python for Data Analysis; O’Reilly: Beijing, China, 2013; p. xiii. 447p. [Google Scholar]
- Perkel, J.M. Programming: Pick up Python. Nature 2015, 518, 125–126. [Google Scholar] [CrossRef] [PubMed]
- Pedregosa, F.; Varoquaux, G.e.l.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- McKinney, W. Python for Data Analysis: Data Wrangling with Pandas, NumPy, and IPython, 2nd ed.; O’Reilly Media, Inc.: Sebastopol, CA, USA, 2018; p. xvi. 524p. [Google Scholar]
- McKinney, W. Pandas: Powerful Python Data Analysis Toolkit; O’Reilly Media, Inc.: Sebastopol, CA, USA, 2011. [Google Scholar]
- Pechenizkiy, M.; Tsymbal, A.; Puuronen, S. PCA-based feature transformation for classification: Issues in medical diagnostics. In Proceedings of the 17th IEEE Symposium on Computer-Based Medical Systems, Proceedings, Bethesda, MD, USA, 24–25 June 2004; pp. 535–540. [Google Scholar]
- Ragno, A.; Baldisserotto, A.; Antonini, L.; Sabatino, M.; Sapienza, F.; Baldini, E.; Buzzi, R.; Vertuani, S.; Manfredini, S. Machine learning data augmentation as a tool to enhance quantitative composition-activity relationships of complex mixtures. A new application to dissect the role of main chemical components in bioactive essential oils. Molecules 2021, 26, 6279. [Google Scholar] [CrossRef] [PubMed]
- Bergstra, J.; Bardenet, R.; Kégl, B.; Bengio, Y. Algorithms for Hyper-Parameter Optimization. In Advances in Neural Information Processing Systems 24 (NIPS 2011); Shawe-Taylor, J., Zemel, R., Bartlett, P., Pereira, F., Weinberger, K.Q., Eds.; Curran Associates Inc.: Red Hook, NY, USA, 2011. [Google Scholar]
- Friedman, J.H. Greedy function approximation: A gradient boosting machine. Ann. Stat. 2001, 29, 1189–1232. [Google Scholar] [CrossRef]
- Choudhary, P.; Kramer, A. Datascience.com Team. Datascienceinc/Skater: Enable Interpretability via Rule Extraction (BRL) (v1.1.0-b1). Zenodo. 2018. Available online: https://zenodo.org/record/1198885#.YmSui9NBxPY (accessed on 21 April 2022).
- Wei, P.; Lu, Z.; Song, J. Variable importance analysis: A comprehensive review. Reliab. Eng. Syst. Saf. 2015, 142, 399–432. [Google Scholar] [CrossRef]
- Baldi, P.; Brunak, S.; Chauvin, Y.; Andersen, C.A.F.; Nielsen, H. Assessing the accuracy of prediction algorithms for classification: An overview. Bioinformatics 2000, 16, 412–424. [Google Scholar] [CrossRef] [Green Version]
- Shorten, C.; Khoshgoftaar, T.M. A survey on image data augmentation for deep learning. J. Big Data 2019, 6, 60. [Google Scholar] [CrossRef]
- Bozovic, M.; Garzoli, S.; Sabatino, M.; Pepi, F.; Baldisserotto, A.; Andreotti, E.; Romagnoli, C.; Mai, A.; Manfredini, S.; Ragno, R. Essential oil extraction, chemical analysis and anti-Candida activity of Calamintha Nepeta (l.) Savi subsp. Glandulosa (req.) ball-new approaches. Molecules 2017, 22, 203. [Google Scholar]
- Garzoli, S.; Bozovic, M.; Baldisserotto, A.; Andreotti, E.; Pepi, F.; Tadic, V.; Manfredini, S.; Ragno, R. Sideritis romana L. subsp. purpurea (Tal. ex Benth.) Heywood, a new chemotype from Montenegro. Nat. Prod. Res. 2018, 32, 1056–1061. [Google Scholar] [CrossRef]
- Garzoli, S.; Bozovic, M.; Baldisserotto, A.; Sabatino, M.; Cesa, S.; Pepi, F.; Vicentini, C.B.; Manfredini, S.; Ragno, R. Essential oil extraction, chemical analysis and anti-Candida activity of Foeniculum vulgare Miller—New approaches. Nat. Prod. Res. 2018, 32, 1254–1259. [Google Scholar] [CrossRef]
- Oliva, A.; Costantini, S.; De Angelis, M.; Garzoli, S.; Bozovic, M.; Mascellino, M.T.; Vullo, V.; Ragno, R. High Potency of Melaleuca alternifolia Essential Oil against Multi-Drug Resistant Gram-Negative bacteria and Methicillin-Resistant Staphylococcus aureus. Molecules 2018, 23, 2584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozovic, M.; Garzoli, S.; Baldisserotto, A.; Andreotti, E.; Cesa, S.; Pepi, F.; Vertuani, S.; Manfredini, S.; Ragno, R. Variation in essential oil content and composition of Ridolfia segetum Moris based on 30-hour prolonged fractionated extraction procedure. Nat. Prod. Res. 2020, 34, 1923–1926. [Google Scholar] [CrossRef] [PubMed]
- Di Vito, M.; Bellardi, M.G.; Sanguinetti, M.; Mondello, F.; Girolamo, A.; Barbanti, L.; Garzoli, S.; Sabatino, M.; Ragno, R.; Vitali, A.; et al. Potent in vitro activity of Citrus aurantium essential oil and vitis vinifera hydrolate against gut yeast isolates from irritable bowel syndrome patients-the right mix for potential therapeutic use. Nutrients 2020, 12, 1329. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, F.; Oliva, A.; Sabatino, M.; Imbriano, A.; Hanieh, P.N.; Garzoli, S.; Mastroianni, C.M.; De Angelis, M.; Miele, M.C.; Arnaut, M.; et al. Antimicrobial essential oil formulation: Chitosan coated nanoemulsions for nose to brain delivery. Pharmaceutics 2020, 12, 678. [Google Scholar] [CrossRef]
- Bozovic, M.; Garzoli, S.; Vujovic, S.; Sapienza, F.; Ragno, R. Foeniculum vulgare Miller, a new chemotype from Montenegro. Plants 2021, 11, 42. [Google Scholar] [CrossRef]
- Vrenna, G.; Artini, M.; Ragno, R.; Relucenti, M.; Fiscarelli, E.V.; Tuccio Guarna Assanti, V.; Papa, R.; Selan, L. Anti-Virulence Properties of Coridothymus capitatus essential oil against Pseudomonas aeruginosa clinical isolates from cystic fibrosis patients. Microorganisms 2021, 9, 2257. [Google Scholar] [CrossRef]
- Artini, M.; Cicatiello, P.; Ricciardelli, A.; Papa, R.; Selan, L.; Dardano, P.; Tilotta, M.; Vrenna, G.; Tutino, M.L.; Giardina, P.; et al. Hydrophobin coating prevents Staphylococcus epidermidis biofilm formation on different surfaces. Biofouling 2017, 33, 601–611. [Google Scholar] [CrossRef]
- Casillo, A.; Papa, R.; Ricciardelli, A.; Sannino, F.; Ziaco, M.; Tilotta, M.; Selan, L.; Marino, G.; Corsaro, M.M.; Tutino, M.L.; et al. Anti-Biofilm activity of a long-chain fatty aldehyde from Antarctic Pseudoalteromonas haloplanktis TAC125 against Staphylococcus epidermidis biofilm. Front. Cell Infect. Microbiol. 2017, 7, 46. [Google Scholar] [CrossRef] [Green Version]
- Ricciardelli, A.; Casillo, A.; Papa, R.; Monti, D.M.; Imbimbo, P.; Vrenna, G.; Artini, M.; Selan, L.; Corsaro, M.M.; Tutino, M.L.; et al. Pentadecanal inspired molecules as new anti-biofilm agents against Staphylococcus epidermidis. Biofouling 2018, 34, 1110–1120. [Google Scholar] [CrossRef]
- Selan, L.; Vrenna, G.; Ettorre, E.; Papa, R.; Artini, M. Virulence of MRSA USA300 is enhanced by sub-inhibitory concentration of two different classes of antibiotics. J. Chemother. 2018, 30, 384–388. [Google Scholar] [CrossRef]
- Artini, M.; Papa, R.; Vrenna, G.; Lauro, C.; Ricciardelli, A.; Casillo, A.; Corsaro, M.M.; Tutino, M.L.; Parrilli, E.; Selan, L. Cold-adapted bacterial extracts as a source of anti-infective and antimicrobial compounds against Staphylococcus aureus. Future Microbiol. 2019, 14, 1369–1382. [Google Scholar] [CrossRef] [PubMed]
- Bossu, M.; Selan, L.; Artini, M.; Relucenti, M.; Familiari, G.; Papa, R.; Vrenna, G.; Spigaglia, P.; Barbanti, F.; Salucci, A.; et al. Characterization of Scardovia wiggsiae biofilm by original Scanning Electron Microscopy protocol. Microorganisms 2020, 8, 807. [Google Scholar] [CrossRef] [PubMed]
- Kifer, D.; Muzinic, V.; Klaric, M.S. Antimicrobial potency of single and combined mupirocin and monoterpenes, thymol, menthol and 1,8-cineole against Staphylococcus aureus planktonic and biofilm growth. J. Antibiot. 2016, 69, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Karuppiah, V.; Thirunanasambandham, R.; Thangaraj, G. Anti-quorum sensing and antibiofilm potential of 1,8-cineole derived from Musa paradisiaca against Pseudomonas aeruginosa strain PAO1. World J. Microbiol. Biotechnol. 2021, 37, 66. [Google Scholar] [CrossRef] [PubMed]
- Lahiri, D.; Nag, M.; Dutta, B.; Dey, S.; Mukherjee, D.; Joshi, S.J.; Ray, R.R. Antibiofilm and anti-quorum sensing activities of eugenol and linalool from Ocimum tenuiflorum against Pseudomonas aeruginosa biofilm. J. Appl. Microbiol. 2021, 131, 2821–2837. [Google Scholar] [CrossRef]
- Kerekes, E.B.; Deak, E.; Tako, M.; Tserennadmid, R.; Petkovits, T.; Vagvolgyi, C.; Krisch, J. Anti-biofilm forming and anti-quorum sensing activity of selected essential oils and their main components on food-related micro-organisms. J. Appl. Microbiol. 2013, 115, 933–9429. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| ID EOs | PaO1 | PA14 | 22P | 25P | 26P | 27P | 37P | 39P |
|---|---|---|---|---|---|---|---|---|
| EO1 | 171.68 ± 8.65 | 102.90 ± 5.14 | 87.26 ± 4.13 | 38.44 ± 1.92 | 191.93 ± 9.60 | 134.12 ± 6.71 | 137.52 ± 6.87 | 142.70 ± 7.13 |
| EO2 | 244.23 ± 12.22 | 54.32 ± 3.26 | 101.82 ± 5.09 | 17.12 ± 1.03 | 113.99 ± 5.70 | 215.33 ± 12.92 | 97.38 ± 4.87 | 116.77 ± 5.84 |
| EO3 | 143.37 ± 7.10 | 152.01 ± 7,60 | NA | 105.72 ± 5.29 | 79.10 ± 4.75 | 59.25 ± 2.96 | NA | 118.93 ± 5.95 |
| EO4 | 183.16 ± 9.16 | 48.88 ± 8.65 | NA | 68.33 ± 3.41 | 193.18 ± 9.66 | 170.23 ± 10.21 | NA | 6.18 ± 0.37 |
| EO5 | 80.92 ± 4.03 | 70.52 ± 4,23 | 89.54 ± 4.77 | 84.55 ± 5.07 | 66.84 ± 4.01 | 90.86 ± 4.54 | 0.59 ± 0.04 | 47.66 ± 2.86 |
| EO6 | 133.04 ± 7.98 | 56.16 ± 2.80 | 86.25 ± 5.17 | 25.20 ± 1.51 | 97.21 ± 4.86 | 169.64 ± 8.48 | 77.81 ± 4.69 | 8.51 ± 0.51 |
| EO7 | 119.72 ± 4.79 | 40.87 ± 2.39 | 93.03 ± 4.65 | 31.41 ± 1.89 | 82.52 ± 5.11 | 211.32 ± 12.68 | 80.80 ± 4.04 | 88.51 ± 4.42 |
| EO8 | 130.36 ± 7.82 | 37.01 ± 2.22 | 97.07 ± 5.83 | 18.85 ± 0.94 | 63.22 ± 3.79 | 197.42 ± 9.87 | 76.18 ± 3.81 | 4.17 ± 0.21 |
| EO9 | 90.15 ± 4.50 | NA | NA | NA | NA | 69.97 ± 3.50 | NA | NA |
| EO10 | 83.80 ± 5.03 | 50.20 ± 2.51 | 84.81 ± 5.09 | 77.93 ± 3.90 | 81.86 ± 4.91 | 46.79 ± 2.33 | 0.57 ± 0.03 | 3.40 ± 0.17 |
| EO11 | 161.28 ± 9.68 | 81.37 ± 4.88 | 26.42 ± 1.32 | 119.18 ± 5.96 | 45.07 ± 2.25 | 28.86 ± 1.44 | 26.21 ± 1.31 | 16.61 ± 0.83 |
| EO12 | 200.22 ± 12.01 | NA | 84.57 ± 8.65 | 16.75 ± 0,83 | 77.79 ± 3.89 | 186.92 ± 11.21 | 85.37 ± 5.12 | 1.27 ± 0.08 |
| EO13 | 150.64 ± 6.02 | 68.27 ± 3.41 | 30.90 ± 1.85 | 75.00 ± 3.75 | 30.53 ± 1.53 | 26.04 ± 1.30 | 50.39 ± 2.52 | 75.64 ± 3.78 |
| EO14 | NA | NA | 152.27 ± 9.14 | NA | 107.26 ± 5.36 | NA | NA | NA |
| EO15 | 66.96 ± 3.35 | 47.92 ± 2.39 | 32.98 ± 1.65 | 357.44 ± 17,87 | 33.60 ± 1.68 | 29.03 ± 1.45 | 48.47 ± 2.42 | 56.85 ± 2,84 |
| EO16 | 91.84 ± 5.51 | 55.12 ± 3.31 | 100.85 ± 5.04 | 79.98 ± 4.80 | 69.69 ± 3,48 | 77.60 ± 4.65 | 0.18 ± 0.01 | 5.17 ± 0.31 |
| EO17 | 174.80 ± 10.49 | 25.28 ± 1.26 | 86.63 ± 4.33 | 33.36 ± 1.67 | 83.67 ± 4.18 | 97.91 ± 4.89 | 98.82 ± 4.94 | 13.81 ± 0.83 |
| EO18 | 94.21 ± 4.71 | 51.51 ± 3.09 | 88.52 ± 5.33 | 64.22 ± 3.85 | 56.53 ± 3.40 | 62.53 ± 9.60 | 0.33 ± 0.02 | 18.14 ± 0.91 |
| EO19 | 96.59 ± 4.83 | 40.94 ± 2.05 | 83.87 ± 4.32 | 148.84 ± 7.48 | 70.91 ± 3.67 | 175.29 ± 7.40 | 68.80 ± 3.32 | 41.84 ± 1.78 |
| EO20 | 101.78 ± 5.09 | 70.73 ± 3.54 | 81.14 ± 5.23 | 76.40 ± 3.75 | 97.94 ± 4.78 | 90.46 ± 4.78 | 0.99 ± 0.04 | 129.74 ± 6.12 |
| EO21 | 121.96 ± 7.32 | 60.76 ± 3.04 | 94.28 ± 5.23 | 39.47 ± 3.85 | 86.12 ± 3.40 | 105.60 ± 5.40 | 219.63 ± 9.90 | 32.92 ± 1.76 |
| EO22 | NA | NA | NA | NA | NA | NA | NA | NA |
| EO23 | 91.76 ± 4.59 | 76.64 ± 4.61 | 85.30 ± 3.21 | 56.88 ± 3.00 | 65.40 ± 3.67 | 83.77 ± 4.41 | 93.96 ± 4.08 | 7.63 ± 3.40 |
| EO24 | 43.68 ± 2.62 | 48.46 ± 2.42 | 61.60 ± 3.56 | 93.97 ± 5.78 | 119.89 ± 5.67 | 51.13 ± 3.40 | 0.72 ± 0.06 | 5.07 ± 2.21 |
| EO25 | 80.63 ± 4,03 | 100.33 ± 5.02 | 62.94 ± 2.45 | 165.61 ± 8.98 | 95.18 ± 4.78 | 25.93 ± 1.78 | 91.99 ± 3.88 | 78.40 ± 3.87 |
| EO26 | 81.08 ± 4.05 | 45.23 ± 2.26 | 280.83 ± 14.34 | 82.41 ± 3.96 | 63.88 ± 3.43 | 52.13 ± 2.61 | 0.89 ± 0.02 | 6.61 ± 3.21 |
| EO27 | 38.50 ± 1.92 | 48.49 ± 2.91 | 60.12 ± 4.12 | 58.27 ± 2.65 | 143.64 ± 7.40 | 146.67 ± 7.89 | 0.34 ± 0.01 | 2.85 ± 0.65 |
| EO28 | 69.88 ± 3,50 | 84.53 ± 5.07 | 83.90 ± 5.34 | 47.96 ± 2.21 | 84.26 ± 5.72 | 116.53 ± 5.12 | 101.67 ± 6.01 | 55.73 ± 3.01 |
| EO29 | 99.57 ± 4.98 | 40.37 ± 2.42 | 68.14 ± 3.89 | 16.64 ± 0.69 | 79.53 ± 3.69 | 125.97 ± 5.67 | 89.27 ± 5.21 | 7.42 ± 3.43 |
| EO30 | 101.11 ± 5.05 | 115.87 ± 5.79 | 79.12 ± 4.88 | 64.16 ± 3.44 | 90.40 ± 4.65 | 120.19 ± 6.71 | 100.04 ± 3.79 | 29.97 ± 1.21 |
| EO31 | 91.35 ± 4.57 | 54.85 ± 2.74 | 116.72 ± 8.67 | 40.53 ± 2.01 | 154.39 ± 7.39 | 102.96 ± 5.12 | 106.85 ± 5.38 | 228.52 ± 9.91 |
| EO32 | NA | NA | NA | NA | NA | NA | NA | NA |
| EO33 | 78.70 ± 3.94 | 38.38 ± 1.92 | 244.16 ± 10.67 | 123.39 ± 6.43 | 122.63 ± 6.40 | 97.78 ± 4.78 | 80.55 ± 4.01 | 107.29 ± 5.21 |
| EO34 | 58.60 ± 2.93 | 43.65 ± 2.19 | 59.81 ± 3.45 | 59.00 ± 2.67 | 76.95 ± 3.61 | 129.69 ± 6.12 | 0.52 ± 0.02 | 6.31 ± 3.01 |
| EO35 | 99.57 ± 5.97 | 40.37 ± 2.02 | 68.14 ± 3.89 | 16.64 ± 0.53 | 79.53 ± 3.23 | 125.97 ± 6.28 | 89.27 ± 3.56 | 7.42 ± 3.74 |
| EO36 | 57.65 ± 2.89 | 65.71 ± 3.29 | 74.02 ± 4.56 | 84.40 ± 4.21 | 87.98 ± 3.89 | 66.84 ± 3.46 | 0.96 ± 0.05 | 122.25 ± 6.02 |
| EO37 | 150.48 ± 7.52 | 117.81 ± 5.80 | NA | 60.23 ± 3.03 | 141.57 ± 7.89 | 109.93 ± 4.99 | NA | NA |
| EO38 | 149.62 ± 7.48 | 64.64 ± 3.23 | 22.61 ± 1.09 | 70.36 ± 3.43 | 48.92 ± 2.78 | 28.28 ± 1.98 | NA | 131.84 ± 5.89 |
| EO39 | NA | NA | NA | NA | NA | NA | NA | NA |
| EO40 | 122.50 ± 7.35 | 32.00 ± 1.60 | 80.43 ± 5.01 | 15.06 ± 0.43 | 67.86 ± 3.41 | 142.49 ± 6.42 | 72.49 ± 3.72 | 13.90 ± 0.99 |
| EO41 | 141.92 ± 7.10 | 46.02 ± 2.31 | 14.36 ± 0.99 | 504.44 ± 19.95 | 128.47 ± 6 | 60.11 ± 3.11 | 98.48 ± 4.41 | 30.01 ± 1.23 |
| EO42 | 86.65 ± 4.33 | 39.05 ± 1.78 | 84.45 ± 5.32 | 15.48 ± 0.32 | 77.26 ± 3.78 | 177.47 ± 8.91 | 88.80 ± 4.28 | 3.43 ± 0.18 |
| EO43 | 127.46 ± 7.65 | 96.81 ± 4.34 | 14.11 ± 0.75 | 105.29 ± 5.42 | 95.01 ± 3.99 | 36.43 ± 1.21 | 71.83 ± 3.32 | 5.89 ± 0.78 |
| EO44 | 71.79 ± 3.60 | 49.55 ± 2.53 | 15.25 ± 0.98 | 50.23 ± 2.32 | 111.58 ± 5.78 | 45.07 ± 2.13 | 88.43 ± 4.21 | 11.95 ± 0.98 |
| EO45 | 148.56 ± 7.43 | 56.12 ± 2.76 | 23.45 ± 1.12 | 68.57 ± 3.79 | 50.81 ± 2.65 | 33.61 ± 1.21 | 38.15 ± 1.89 | 95.07 ± 4.01 |
| EO46 | 147.21 ± 7.40 | 33.51 ± 1.78 | 15.90 ± 0.88 | 47.32 ± 2.01 | 103.80 ± 5.76 | 70.50 ± 3.56 | NA | NA |
| EO47 | 58.99 ± 2.95 | 66.60 ± 3.45 | 22.08 ± 1.23 | 330.42 ± 14.54 | 24.85 ± 1.21 | 32.98 ± 1.67 | 33.64 ± 1.17 | 66.73 ± 3.23 |
| EO48 | 304.94 ± 15.25 | NA | 50.99 ± 3.45 | 128.35 ± 7.89 | 244.63 ± 11.24 | 169.66 ± 7.47 | 11.06 ± 4.56 | NA |
| EO49 | 78.66 ± 6.71 | 38.08 ± 1.92 | 142.03 ± 5.99 | 78.75 ± 3.65 | 94.61 ± 5.40 | 74.11 ± 3.33 | 131.36 ± 6.12 | 31.78 ± 1.12 |
| EO50 | 103.14 ± 5.16 | 85.60 ± 4.76 | 12.10 ± 0.65 | 53.22 ± 3.01 | 122.85 ± 6.23 | 104.33 ± 5.78 | 94.66 ± 4.65 | 101.57 ± 5.62 |
| EO51 | 102.62 ± 5.13 | 64.56 ± 3.76 | 14.08 ± 0.38 | 47.79 ± 2.94 | 104.82 ± 5.24 | 52.98 ± 1.98 | 82.85 ± 4.10 | 4.75 ± 0.28 |
| EO52 | 113.76 ± 6.82 | 70.07 ± 3.56 | 71.95 ± 3.89 | 21.66 ± 1.09 | 172.35 ± 7.91 | 193.17 ± 6.78 | 87.94 ± 4.21 | 1.85 ± 0.09 |
| EO53 | 134.51 ± 6.71 | 58.00 ± 2.58 | 15.85 ± 0.94 | 46.60 ± 2.45 | 134.77 ± 35.76 | 56.71 ± 2.12 | 76.66 ± 3.56 | 5.33 ± 0.27 |
| EO54 | 100.73 ± 5.04 | 50.85 ± 2.27 | 17.21 ± 1.52 | 47.43 ± 2.76 | 106.48 ± 4.91 | 58.51 ± 3.23 | 25.76 ± 1.21 | 6.26 ± 0.82 |
| EO55 | 118.70 ± 5.93 | 66.92 ± 3.34 | 144.27 ± 7.98 | 34.77 ± 1.96 | 86.90 ± 4.20 | 58.66 ± 3.01 | 118.57 ± 5.67 | 60.23 ± 3.21 |
| EO56 | 90.11 ± 4.50 | 71.63 ± 6.76 | 110.29 ± 6.89 | 45.86 ± 2.91 | 91.28 ± 4.61 | 87.53 ± 3.78 | 163.12 ± 8.54 | 66.06 ± 3.21 |
| EO57 | 55.72 ± 3.34 | 69.00 ± 4.65 | 15.78 ± 0.54 | 47.85 ± 2.04 | 190.73 ± 9.67 | 110.55 ± 6.11 | 92.72 ± 3.78 | 88.24 ± 3.79 |
| EO58 | 76.24 ± 3.81 | 37.82 ± 2.18 | 21.54 ± 1.33 | 153.50 ± 6.72 | 78.89 ± 3.46 | 34.90 ± 1.67 | 54.29 ± 2.21 | 102.60 ± 3.89 |
| EO59 | 160.70 ± 8.03 | 44.94 ± 2.56 | 17.32 ± 0.77 | 46.50 ± 2.78 | 166.22 ± 5.89 | 106.13 ± 5.43 | NA | NA |
| EO60 | 232.46 ± 11.62 | 137.74 ± 7.53 | 82.93 ± 4.67 | 23.82 ± 1.21 | 284.60 ± 19.11 | 280.46 ± 7.12 | NA | NA |
| EO61 | 352.32 ± 17.67 | 652.82 ± 38.65 | 105.04 ± 5.33 | 67.31 ± 3.49 | 627.58 ± 28.12 | 210.31 ± 8.98 | 58.53 ± 3.40 | NA |
| Number Values | 57 | 54 | 54 | 56 | 57 | 57 | 49 | 50 |
| Model | Strain | Threshold | ML Method | MCC | ACC | F1 | |||
|---|---|---|---|---|---|---|---|---|---|
| Fit | CV | Fit | CV | Fit | CV | ||||
| F1 | PA14 | 40 | gb | 1.00 | 0.62 | 1.00 | 0.91 | 1.00 | 0.67 |
| F2 | 22P | dt | 0.66 | 0.11 | 0.85 | 0.69 | 0.69 | 0.19 | |
| F3 | 25P | gb | 1.00 | 0.47 | 1.00 | 0.80 | 1.00 | 0.59 | |
| F4 | 27P | gb | 1.00 | 0.66 | 1.00 | 0.91 | 1.00 | 0.71 | |
| F5 | 37P | gb | 0.71 | 0.71 | 0.88 | 0.88 | 0.80 | 0.80 | |
| F6 | 39P | rf | 1.00 | 0.59 | 1.00 | 0.80 | 1.00 | 0.82 | |
| F7 | PAO1 | 120 | svm | 0.41 | 0.56 | 0.72 | 0.79 | 0.77 | 0.84 |
| F8 | 25P | dt | 0.78 | 0.35 | 0.93 | 0.79 | 0.96 | 0.87 | |
| F9 | 26P | dt | 1.00 | 0.64 | 1.00 | 0.86 | 1.00 | 0.90 | |
| F10 | 27P | svm | 0.50 | 0.64 | 0.79 | 0.84 | 0.85 | 0.89 | |
| F11 | PAO1 | svm | 1.00 | 0.88 | 1.00 | 0.98 | 1.00 | 0.99 | |
| Threshold | Anti-Biofilm | Strains | Pro-Biofilm | Strains |
|---|---|---|---|---|
| 40 | linalool | 22P, 25P, 27P, 39P | ||
| eucalyptol | 27P, 39P | |||
| linalyl anthranilate | 22P, 37P | |||
| geranyl acetate | 22P, 25P | |||
| bornyl acetate | 37P | |||
| cis-3-pinanone | 27P | |||
| cis-geraniol | 22P, 37P, 39P | |||
| sabinene | 25P | |||
| β-caryophyllene | PA14, 25P | β-caryophyllene | 22P, 27P, 37P, 39P | |
| α-pinene | 27P | α-pinene | 39P | |
| β-pinene | 39P | |||
| limonene | 27P | limonene | 22P, 25P | |
| carvacrol | PA14 | |||
| p-cymene | PA14 | p-cymene | 22P | |
| 120 | eucalyptol | PAO1 | ||
| linalyl anthranilate | PAO1 | linalyl anthranilate | 37P | |
| o-cymene | PAO1 | |||
| linalool | PAO1 | linalool | 25P, 27P | |
| thymol | PAO1, 25P | |||
| limonene | 25P, 26P | limonene | PAO1, 37P | |
| p-cymene | PAO1, 37P | |||
| citronellal | 37P | |||
| terpinen-4-ol | PAO1 | |||
| α-pinene | 37P | α-pinene | 26P | |
| carvacrol | PAO1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Artini, M.; Papa, R.; Sapienza, F.; Božović, M.; Vrenna, G.; Tuccio Guarna Assanti, V.; Sabatino, M.; Garzoli, S.; Fiscarelli, E.V.; Ragno, R.; et al. Essential Oils Biofilm Modulation Activity and Machine Learning Analysis on Pseudomonas aeruginosa Isolates from Cystic Fibrosis Patients. Microorganisms 2022, 10, 887. https://doi.org/10.3390/microorganisms10050887
Artini M, Papa R, Sapienza F, Božović M, Vrenna G, Tuccio Guarna Assanti V, Sabatino M, Garzoli S, Fiscarelli EV, Ragno R, et al. Essential Oils Biofilm Modulation Activity and Machine Learning Analysis on Pseudomonas aeruginosa Isolates from Cystic Fibrosis Patients. Microorganisms. 2022; 10(5):887. https://doi.org/10.3390/microorganisms10050887
Chicago/Turabian StyleArtini, Marco, Rosanna Papa, Filippo Sapienza, Mijat Božović, Gianluca Vrenna, Vanessa Tuccio Guarna Assanti, Manuela Sabatino, Stefania Garzoli, Ersilia Vita Fiscarelli, Rino Ragno, and et al. 2022. "Essential Oils Biofilm Modulation Activity and Machine Learning Analysis on Pseudomonas aeruginosa Isolates from Cystic Fibrosis Patients" Microorganisms 10, no. 5: 887. https://doi.org/10.3390/microorganisms10050887
APA StyleArtini, M., Papa, R., Sapienza, F., Božović, M., Vrenna, G., Tuccio Guarna Assanti, V., Sabatino, M., Garzoli, S., Fiscarelli, E. V., Ragno, R., & Selan, L. (2022). Essential Oils Biofilm Modulation Activity and Machine Learning Analysis on Pseudomonas aeruginosa Isolates from Cystic Fibrosis Patients. Microorganisms, 10(5), 887. https://doi.org/10.3390/microorganisms10050887