
Genomic and Functional Variation of the Chlorophyll d-Producing Cyanobacterium Acaryochloris marina

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Laboratory Strain Isolation
2.2. Genomics
2.3. Phylogenetics
2.4. Genomics and Bioinformatics
2.5. Nitrogen Fixation and Salt Tolerance Experiments
3. Results and Discussion
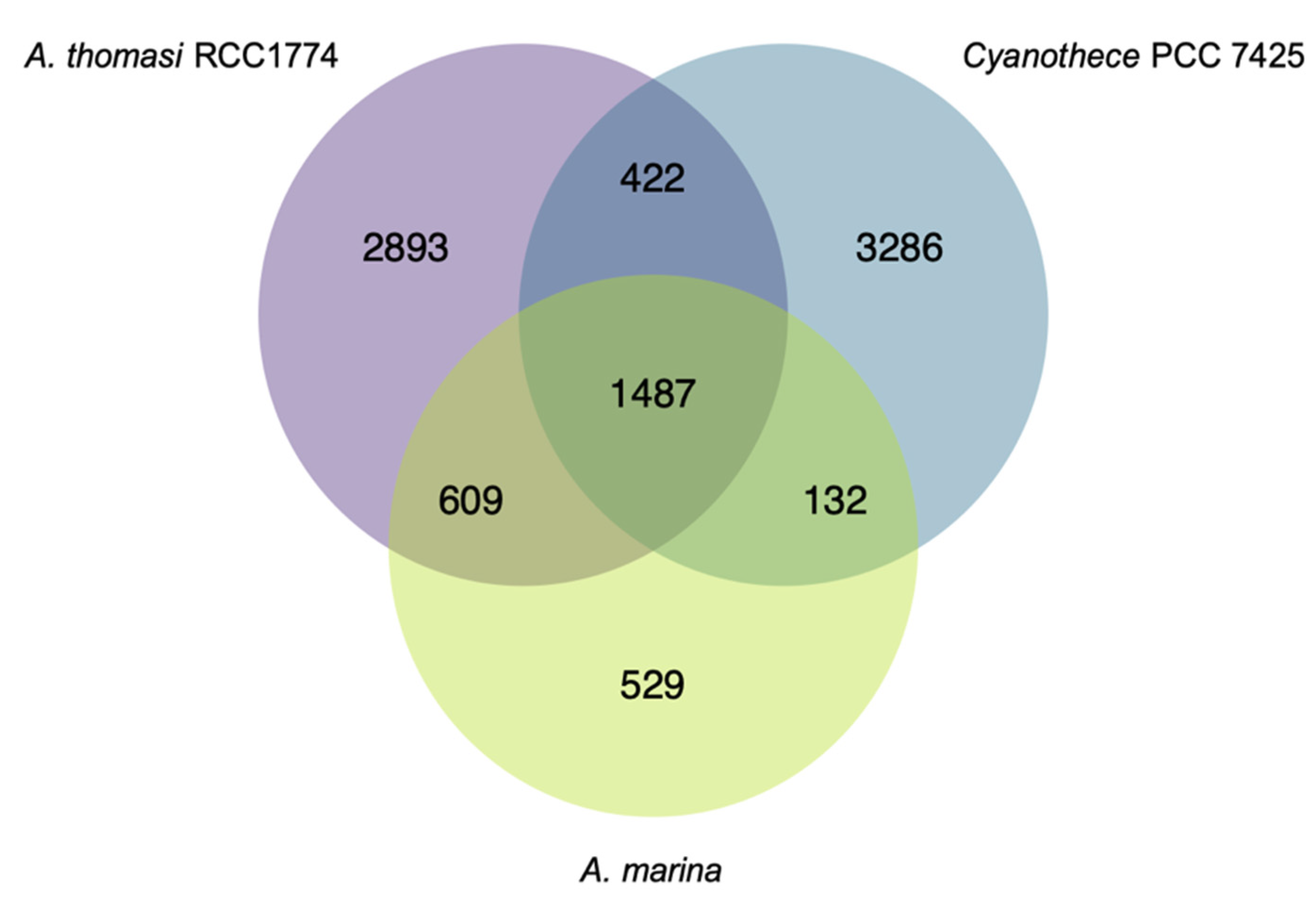
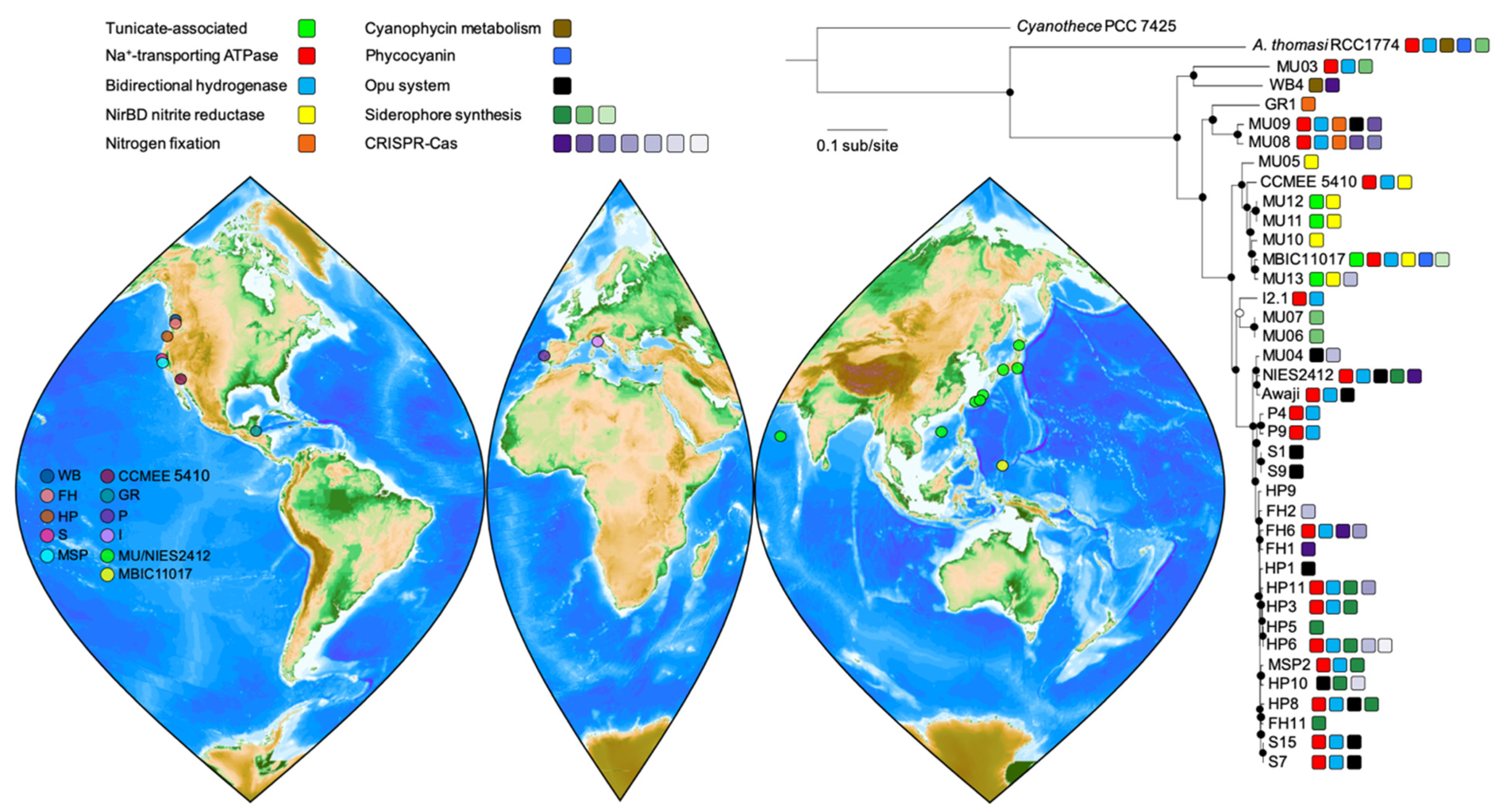
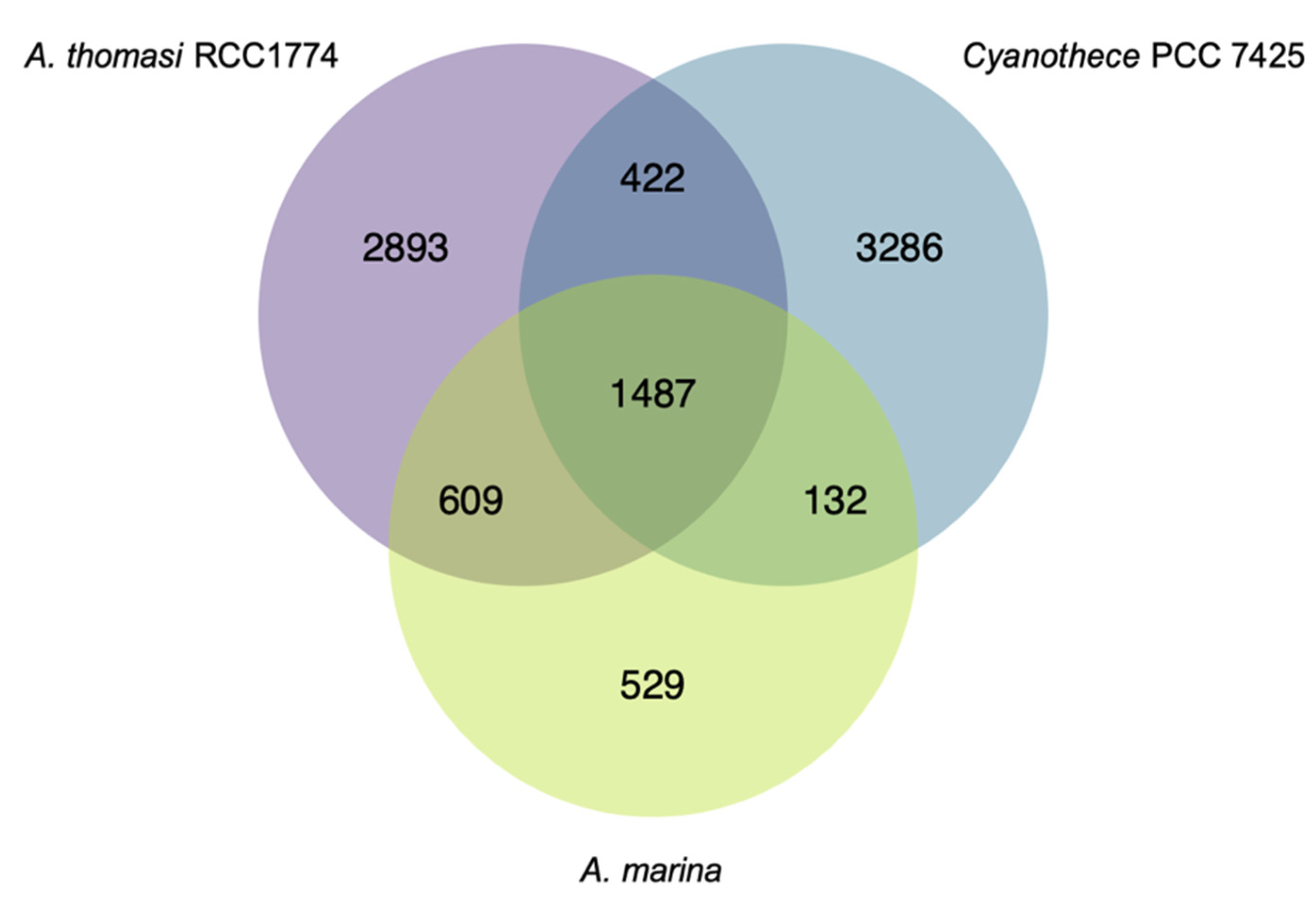
3.1. A. marina Phylogeny and Core Genome
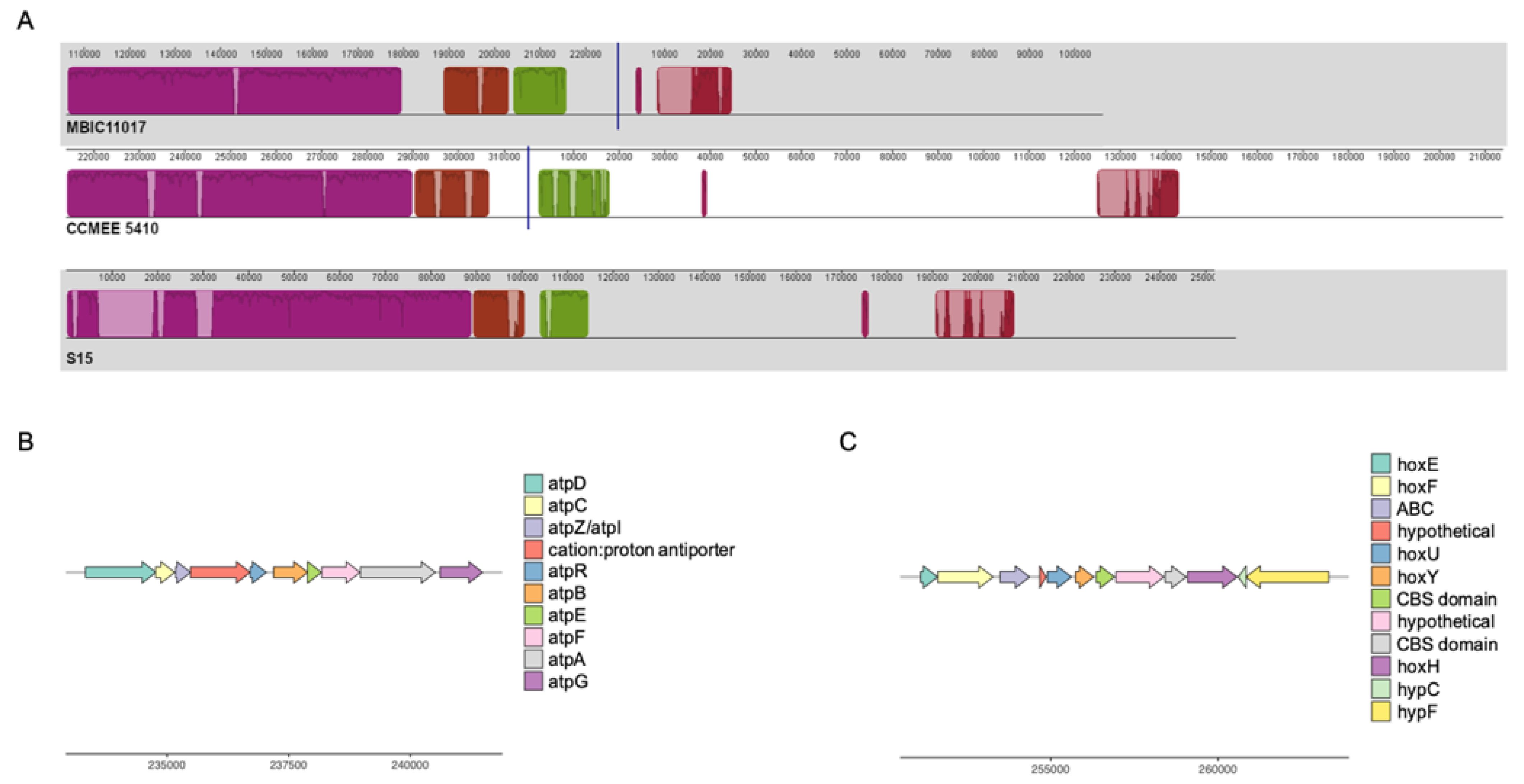
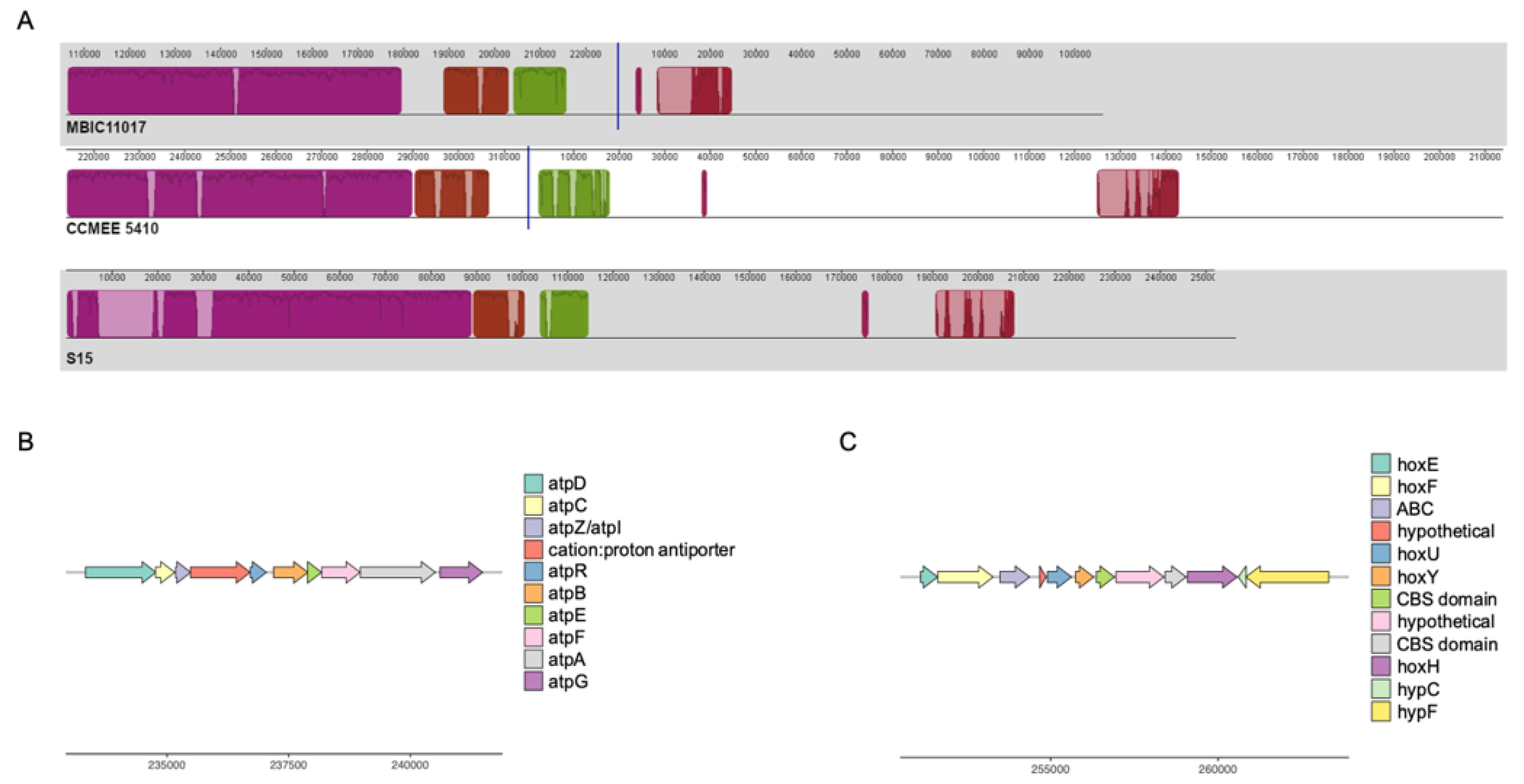
3.2. Differential Retention of Ancestral Plasmid Gene Content Contributes to A. marina Functional Variation
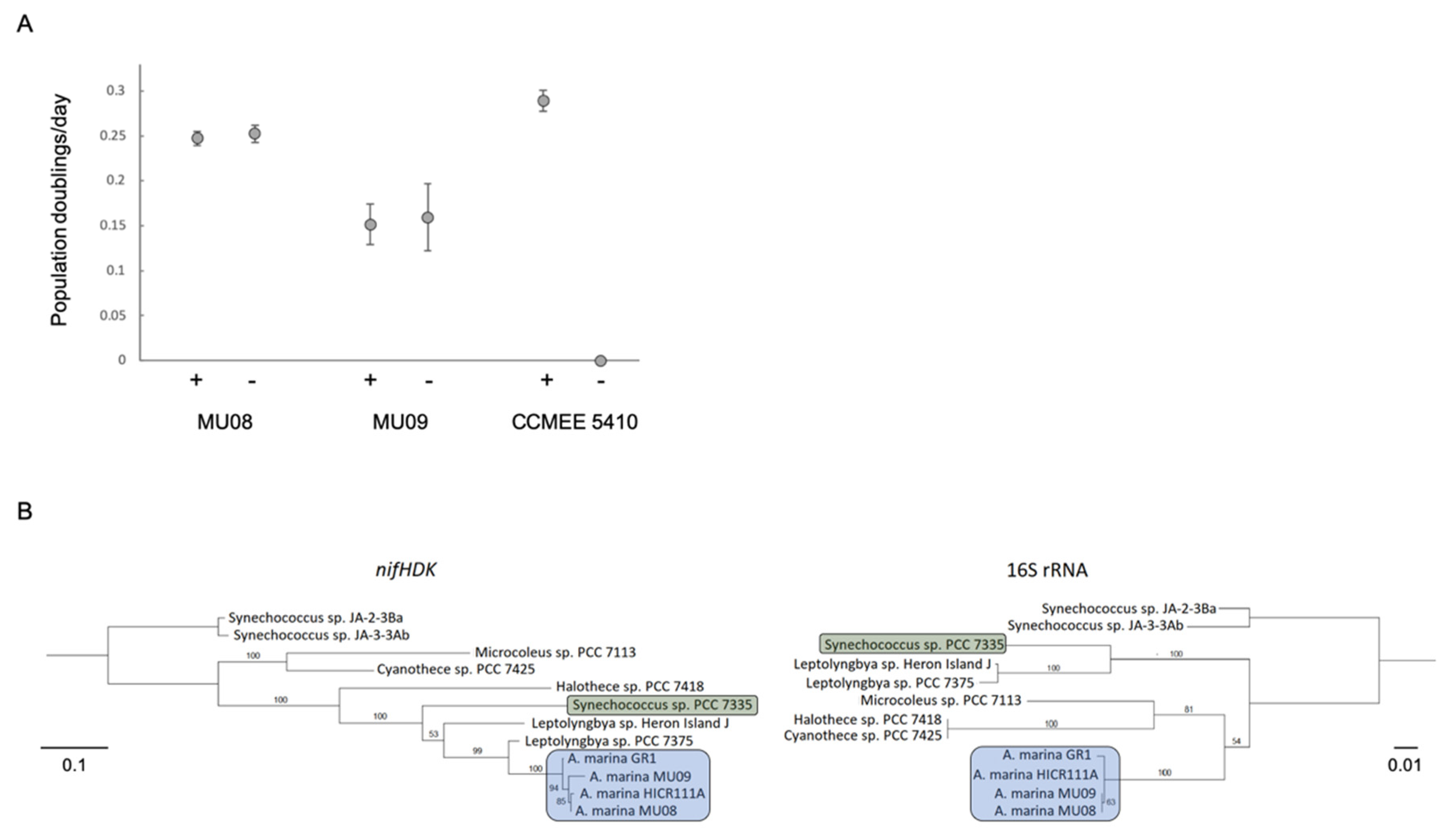
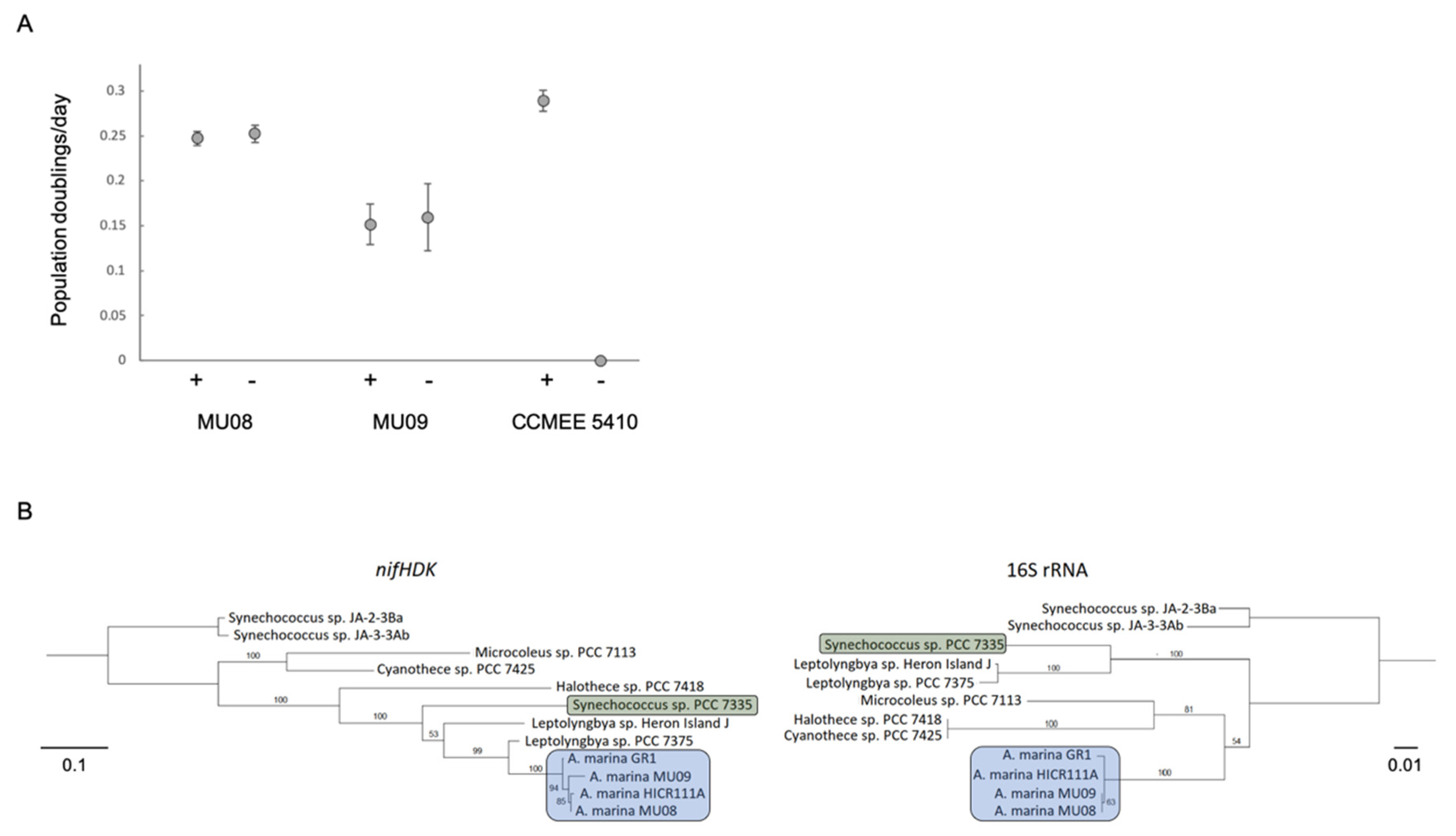
3.3. HGT and the Evolution of Nitrogen Metabolism
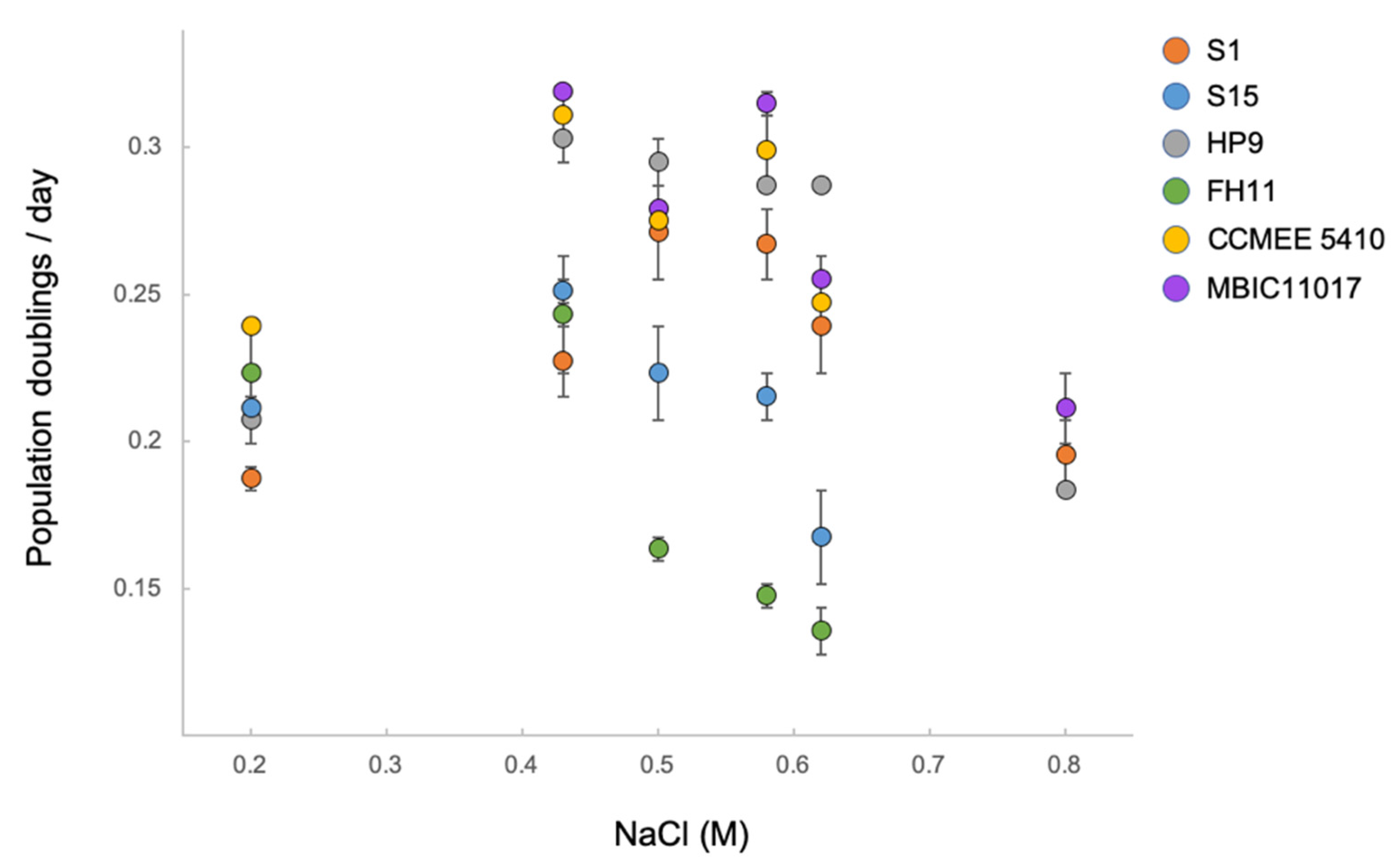
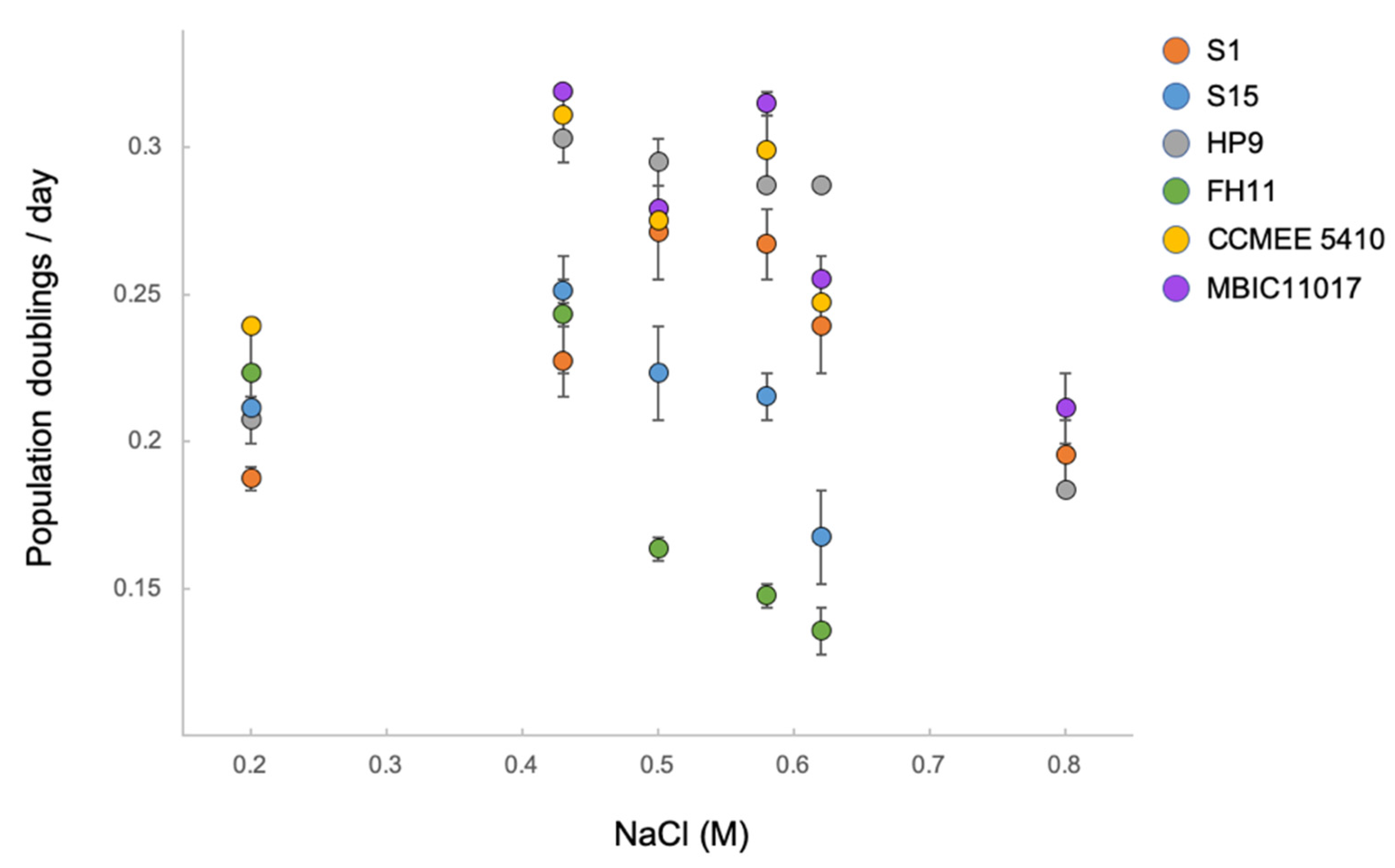
3.4. Ecotypic Variation in Salt Tolerance
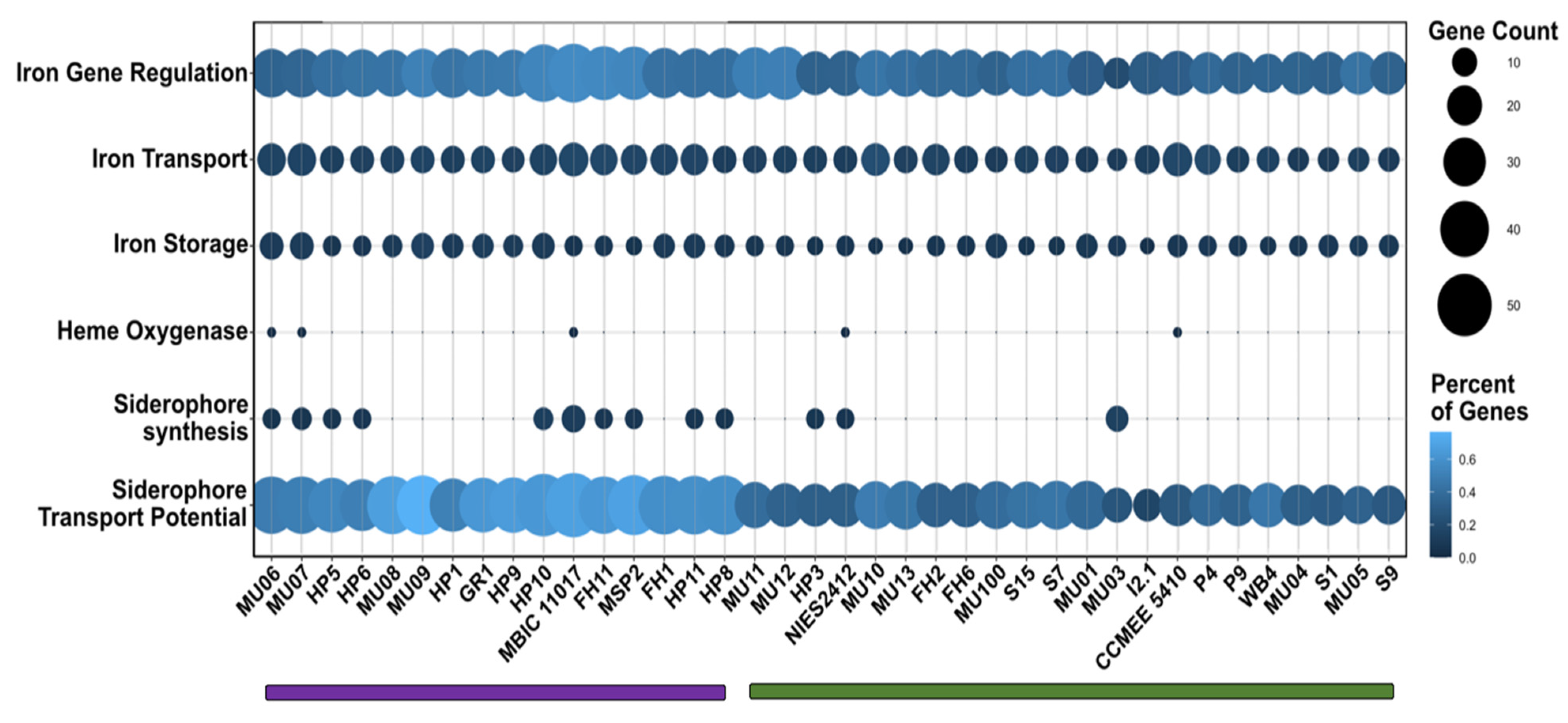
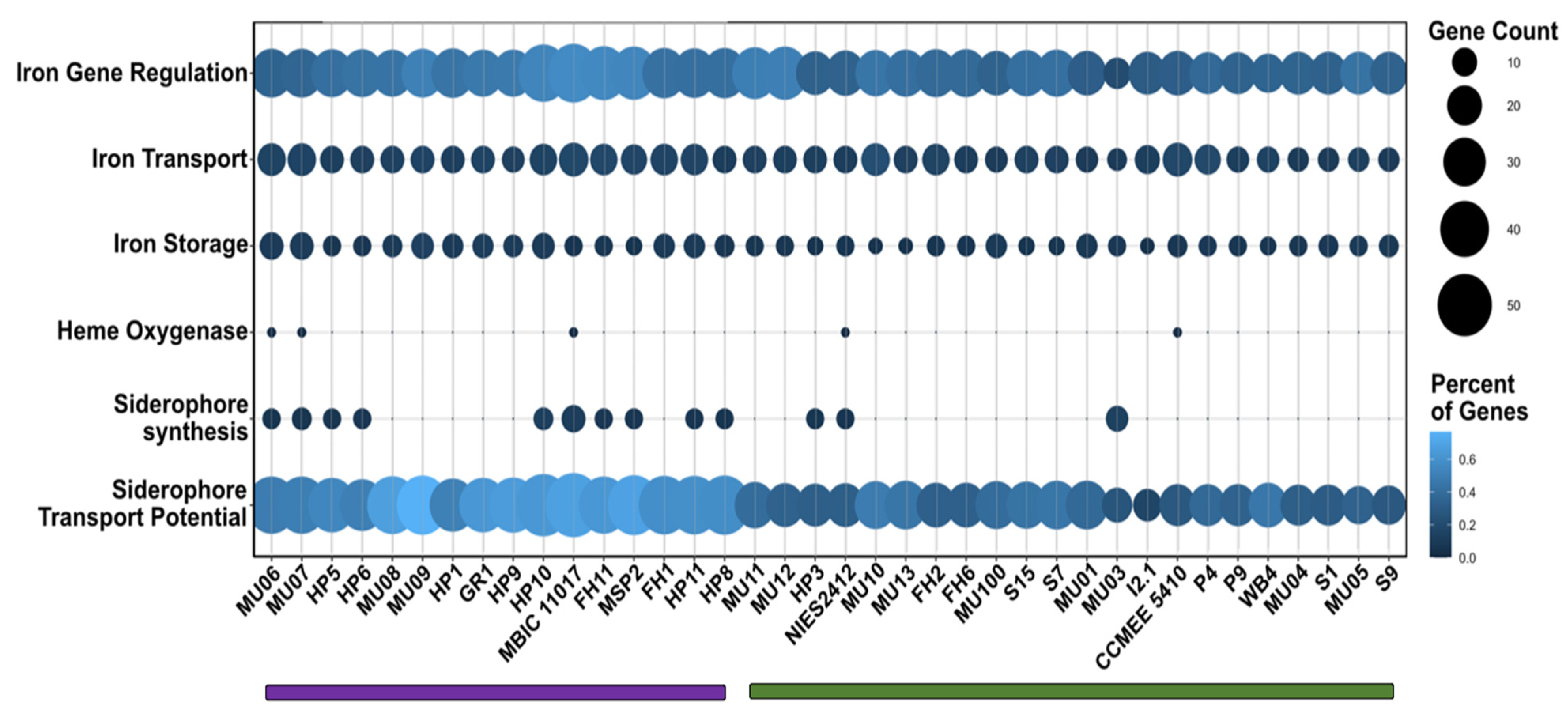
3.5. Iron Metabolism
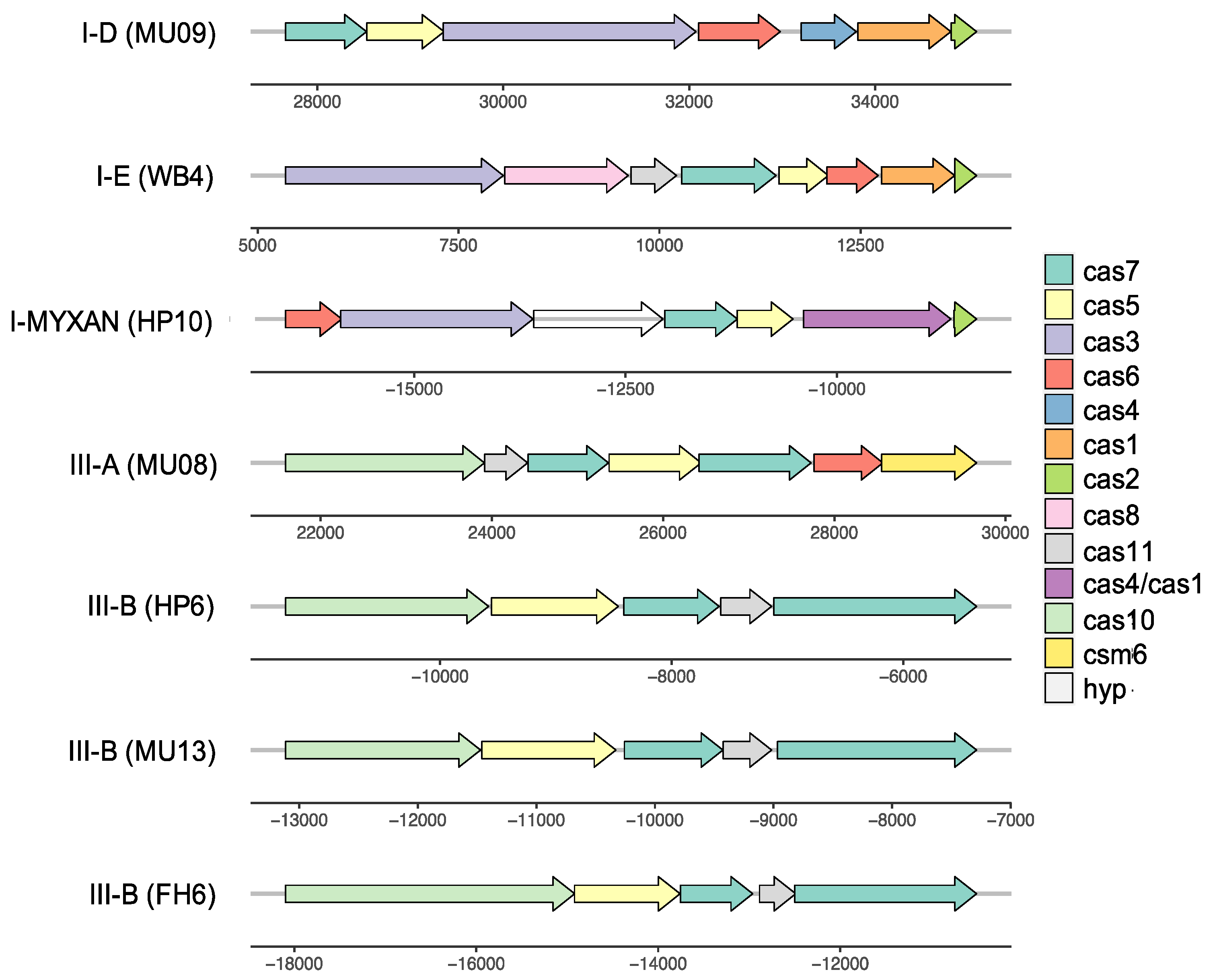
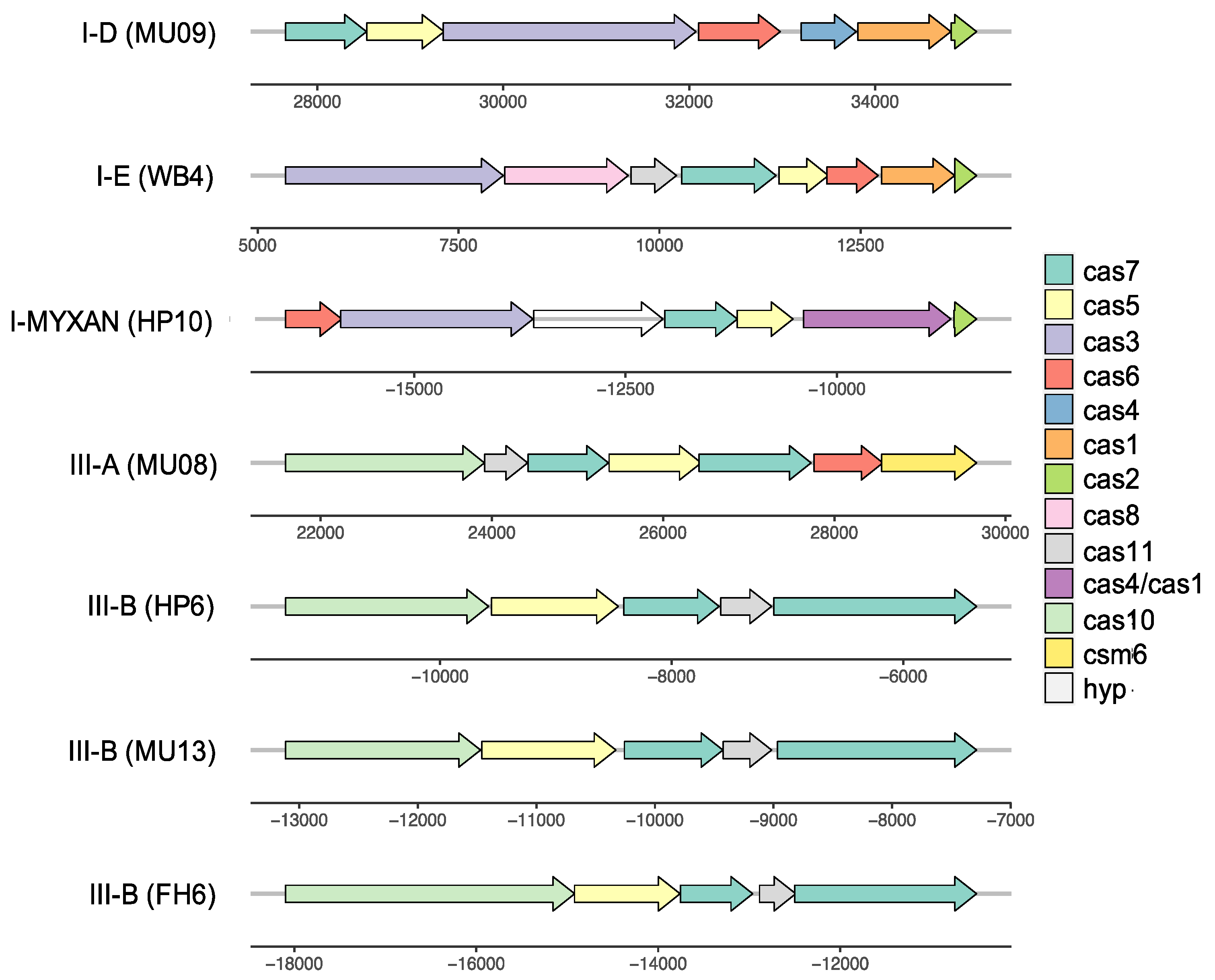
3.6. Complex History of CRISPR-Cas Systems
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miyashita, H.; Ikemoto, H.; Kurano, N.; Adachi, K.; Chihara, M.; Miyachi, S. Chlorophyll d as a major pigment. Nature 1996, 383, 402. [Google Scholar] [CrossRef]
- Behrendt, L.; Larkum, A.W.D.; Trampe, E.; Norman, A.; Sørensen, S.J.; Kühl, M. Microbial diversity of biofilm communities in microniches associated with the didemnid ascidian Lissoclinum patella. ISME J. 2012, 6, 1222–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashiyama, Y.; Miyashita, H.; Ohkubo, S.; Ogawa, N.O.; Chikaraishi, Y.; Takano, Y.; Suga, H.; Toyofuku, T.; Nomaki, H.; Kitazato, H.; et al. Evidence of global chlorophyll d. Science 2008, 321, 658. [Google Scholar] [CrossRef] [PubMed]
- Murakami, A.; Miyashita, H.; Iseki, M.; Adachi, K.; Mimuro, M. Chlorophyll d in an epiphytic cyanobacterium of red algae. Science 2004, 303, 1633. [Google Scholar] [CrossRef] [PubMed]
- Mohr, R.; Voß, B.; Schliep, M.; Kurz, T.; Maldener, I.; Adams, D.G.; Larkum, A.D.W.; Chen, M.; Hess, W.R. A new chlorophyll d-containing cyanobacterium: Evidence for niche adaptation in the genus Acaryochloris. ISME J. 2010, 4, 1456–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larkum, A.W.; Chen, M.; Li, Y.; Schliep, M.; Trampe, E.; West, J.; Salih, A.; Kühl, M. A novel epiphytic Chlorophyll d-containing cyanobacterium isolated from a mangrove-associated red alga. J. Phycol. 2012, 48, 1320–1327. [Google Scholar] [CrossRef]
- Behrendt, L.; Nielsen, J.L.; Sørensen, S.J.; Larkum, A.W.D.; Winther, J.R.; Kühl, M. Rapid TaqMan-based quantification of Chlorophyll d-containing cyanobacteria in the genus Acaryochloris. Appl. Environ. Microbiol. 2014, 80, 3244–3249. [Google Scholar] [CrossRef] [Green Version]
- Ulrich, N.J.; Uchida, H.; Kanesaki, Y.; Hirose, E.; Murakami, A.; Miller, S.R. Reacquisition of light-harvesting genes in a marine cyanobacterium confers a broader solar niche. Curr. Biol. 2021, 31, 1539–1546. [Google Scholar] [CrossRef]
- Goh, F.; Allen, M.A.; Leuko, S.; Kawaguchi, T.; Decho, A.W.; Burns, B.P.; Neilan, B. Determining the specific microbial populations and their spatial distribution within the stromatolite ecosystem of Shark Bay. ISME J. 2009, 3, 383–396. [Google Scholar] [CrossRef] [Green Version]
- Wood, A.M.; Miller, S.R.; Li, W.K.W.; Castenholz, R.W. Preliminary studies of cyanobacteria, picoplankton, and virioplankton in the Salton Sea with special attention to phylogenetic diversity among eight strains of filamentous cyanobacteria. Hydrobiologia 2002, 473, 77–92. [Google Scholar] [CrossRef]
- Miller, S.R.; Augustine, S.; Le Olson, T.; Blankenship, R.E.; Selker, J.; Wood, A.M. Discovery of a free-living chlorophyll d-producing cyanobacterium with a hybrid proteobacterial/cyanobacterial small-subunit rRNA gene. Proc. Natl. Acad. Sci. USA 2005, 102, 850–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleming, E.D.; Prufert-Bebout, L. Characterization of cyanobacterial communities from high-elevation lakes in the Bolivian Andes. J. Geophys. Res. 2010, 115. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.-C.; Li, Z.-K.; Yin, Y.-C.; Li, Y.; Jia, Y.; Chen, M.; Qiu, B.-S. Widespread occurrence and unexpected diversity of red-shifted chlorophyll producing cyanobacteria in humid subtropical forest ecosystems. Environ. Microbiol. 2019, 21, 1497–1510. [Google Scholar] [CrossRef] [PubMed]
- Swingley, W.D.; Chen, M.; Cheung, P.C.; Conrad, A.L.; Dejesa, L.C.; Hao, J.; Honchak, B.M.; Karbach, L.E.; Kurdoglu, A.; Lahiri, S.; et al. Niche adaptation and genome expansion in the chlorophyll d-producing cyanobacterium Acaryochloris marina. Proc. Natl. Acad. Sci. USA 2008, 105, 2005–2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, S.R.; Wood, A.M.; Blankenship, R.E.; Kim, M.; Ferriera, S. Dynamics of gene duplication in the genomes of chlorophyll d-producing cyanobacteria: Implications for the ecological niche. Genome Biol. Evol. 2011, 3, 601–613. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comp. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Wick, R.R.; Schultz, M.B.; Zobel, J.; Holt, K.E. Bandage: Interactive visualization of de novo genome assemblies. Bioinformatics 2015, 31, 3350–3352. [Google Scholar] [CrossRef] [Green Version]
- Wood, D.E.; Lu, J.; Langmead, B. Improved metagenomic analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef] [Green Version]
- Brettin, T.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; et al. RASTtk: A modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci. Rep. 2015, 5, 8365. [Google Scholar] [CrossRef] [Green Version]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emms, D.M.; Kelly, S. OrthoFinder: Phylogenetic orthology inference for comparative genomics. Genome Biol. 2019, 20, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2014, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Shimodaira, H.; Hasegawa, M. Multiple comparisons of log-likelihoods with applications to phylogenetic inference. Mol. Biol. Evol. 1999, 16, 1114–1116. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Huerta-Cepas, J.; Szklarczyk, D.; Heller, D.; Hernández-Plaza, A.; Forslund, S.K.; Cook, H.V.; Mende, D.R.; Letunic, I.; Rattei, T.; Jensen, L.J.; et al. eggNOG 5.0: A hierarchical, functionally and phylogenetically annotated orthology resource based on 5090 organisms and 2502 viruses. Nucleic Acids Res. 2018, 47, D309–D314. [Google Scholar] [CrossRef] [Green Version]
- Garber, A.I.; Nealson, K.H.; Okamoto, A.; McAllister, S.M.; Chan, C.S.; Barco, R.A.; Merino, N. FeGenie: A comprehensive tool for the identification of iron genes and iron gene neighborhoods in genome and metagenome assemblies. Front Microbiol. 2020, 11, 37. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, R.; Shimodaira, H. Pvclust: An R package for assessing the uncertainty in hierarchical clustering. Bioinformatics 2006, 22, 1540–1542. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grissa, I.; Vergnaud, G.; Pourcel, C. CRISPRFinder: A web tool to identify clustered regularly interspaced short palindromic repeats. Nucleic Acids Res. 2007, 35, W52–W57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, A.; Gagnon, J.N.; Brouns, S.J.J.; Fineran, P.C.; Brown, C.M. CRISPRTarget: Bioinformatic prediction and analysis of crRNA targets. RNA Biol. 2013, 10, 817–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohkubo, S.; Miyashita, H. Selective detection and phylogenetic diversity of Acaryochloris spp. that exist in association with didemnid ascidians and sponge. Microbes Environ. 2012, 27, 217–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schliep, M.; Crossett, B.; Willows, R.D.; Chen, M. 18O labeling of chlorophyll d in Acaryochloris marina reveals that chlorophyll a and molecular oxygen are precursors. J. Biol. Chem. 2010, 285, 28450–28456. [Google Scholar] [CrossRef] [Green Version]
- Yoneda, A.; Wittmann, B.J.; King, J.D.; Blankenship, R.E.; Dantas, G. Transcriptomic analysis illuminates genes involved in chlorophyll synthesis after nitrogen starvation in Acaryochloris sp. CCMEE 5410. Photosynth. Res. 2016, 129, 171–182. [Google Scholar] [CrossRef]
- Barry, S.M.; Challis, G.L. Mechanism and catalytic diversity of Rieske non-heme iron-dependent oxygenases. ACS Catal. 2013, 3, 2362–2370. [Google Scholar] [CrossRef] [Green Version]
- Sciara, G.; Kendrew, S.G.; Miele, A.E.; Marsh, N.G.; Federici, L.; Malatesta, F. The structure of ActVA-Orf6, a novel type of monooxygenase involved in actinorhodin biosynthesis. EMBO J. 2003, 22, 205–215. [Google Scholar] [CrossRef] [Green Version]
- Denisov, I.G.; Makris, T.M.; Sligar, S.G.; Schlichting, I. Structure and chemistry of cytochrome P450. Chem. Rev. 2005, 105, 2253–2277. [Google Scholar] [CrossRef]
- Pružinská, A.; Tanner, G.; Anders, I.; Roca, M.; Hörtensteiner, S. Chlorophyll breakdown: Pheophorbide a oxygenase is a Rieske-type iron–sulfur protein, encoded by the accelerated cell death 1 gene. Proc. Natl. Acad. Sci. USA 2003, 100, 15259–15264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soontharapirakkul, K.; Promden, W.; Yamada, N.; Kageyama, H.; Incharoensakdi, A.; Iwamoto-Kihara, A.; Takabe, T. Halotolerant cyanobacterium Aphanothece halophytica contains an Na+-dependent F1F0-ATP synthase with a potential role in salt-stress tolerance. J. Biol. Chem. 2011, 286, 10169–10176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soontharapirakkul, K.; Incharoensakdi, A. Na+-stimulated ATPase of alkaliphilic halotolerant cyanobacterium Aphanothece halophytica translocates Na+ into proteoliposomes via Na+ uniport mechanism. BMC Biochem. 2010, 11, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, S.; Iglesias-Cans, M.; Krah, A.; Yildiz, Ö.; Leone, V.; Matthies, D.; Cook, G.M.; Faraldo-Gómez, J.D.; Meier, T. A New Type of Na+-driven ATP synthase membrane rotor with a two-carboxylate ion-coupling motif. PLoS Biol. 2013, 11, e1001596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leone, V.; Pogoryelov, D.; Meier, T.; Faraldo-Gómez, J.D. On the principle of ion selectivity in Na⁺/H⁺-coupled membrane proteins: Experimental and theoretical studies of an ATP synthase rotor. Proc. Natl. Acad. Sci. USA 2015, 112, E1057–E1066. [Google Scholar] [CrossRef] [Green Version]
- Barz, M.; Beimgraben, C.; Staller, T.; Germer, F.; Opitz, F.; Marquardt, C.; Schwarz, C.; Gutekunst, K.; Vanselow, K.H.; Schmitz, R.; et al. Distribution analysis of hydrogenases in surface waters of marine and freshwater environments. PLoS ONE 2010, 5, e13846. [Google Scholar] [CrossRef]
- McIntosh, C.L.; Germer, F.; Schulz, R.; Appel, J.; Jones, A.K. The [NiFe]-hydrogenase of the cyanobacterium Synechocystis sp. PCC 6803 works bidirectionally with a bias to H2 production. J. Am. Chem. Soc. 2011, 133, 11308–11319. [Google Scholar] [CrossRef]
- Appel, J.; Phunpruch, S.; Steinmüller, K.; Schulz, R. The bidirectional hydrogenase of Synechocystis sp. PCC 6803 works as an electron valve during photosynthesis. Arch. Microbiol. 2000, 173, 333–338. [Google Scholar] [CrossRef]
- Carrieri, D.; Wawrousek, K.; Eckert, C.; Yu, J.; Maness, P.-C. The role of the bidirectional hydrogenase in cyanobacteria. Bioresour. Tech. 2011, 102, 8368–8377. [Google Scholar] [CrossRef]
- Gutekunst, K. The Bidirectional NiFe-hydrogenase in Synechocystis sp. PCC 6803 is reduced by flavodoxin and ferredoxin and is essential under mixotrophic, nitrate-limiting conditions. J. Biol. Chem. 2014, 289, 1930–1937. [Google Scholar] [CrossRef] [Green Version]
- Khanna, N.; Lindblad, P. Cyanobacterial hydrogenases and hydrogen metabolism revisited: Recent progress and future prospects. Int. J. Mol. Sci. 2015, 16, 10537–10561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiss, É.; Kós, P.B.; Chen, M.; Vass, I. Functioning of the bidirectional hydrogenase in different unicellular cyanobacteria. In Photosynthesis Research for Food, Fuel and the Future; Kuang, T., Lu, C., Zhang, L., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 733–736. [Google Scholar]
- Loughlin, P.; Lin, Y.; Chen, M. Chlorophyll d and Acaryochloris marina: Current status. Photosynth. Res. 2013, 116, 277–293. [Google Scholar] [CrossRef] [PubMed]
- Taylor, B.L.; Zhulin, I.B. PAS domains: Internal sensors of oxygen, redox potential, and light. Microbiol. Mol. Biol. Rev. 1999, 63, 479–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sankar, P.; Lee, J.H.; Shanmugam, K.T. Gene-product relationships of fhlA and fdv genes of Escherichia coli. J. Bacteriol. 1988, 170, 5440–5445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doello, S.; Burkhardt, M.; Forchhammer, K. The essential role of sodium bioenergetics and ATP homeostasis in the developmental transitions of a cyanobacterium. Curr. Biol. 2021, 31, 1606–1615. [Google Scholar] [CrossRef] [PubMed]
- Harrison, E.; Brockhurst, M.A. Plasmid-mediated horizontal gene transfer is a coevolutionary process. Trends Microbiol. 2012, 20, 262–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- San Millan, A.; MacLean, R.C. Fitness costs of plasmids: A limit to plasmid transmission. Microbiol. Spectr. 2017, 5, 65–79. [Google Scholar] [CrossRef] [Green Version]
- Harrison, E.; Guymer, D.; Spiers, A.J.; Paterson, S.; Brockhurst, M.A. Parallel compensatory evolution stabilizes plasmids across the parasitism-mutualism continuum. Curr. Biol. 2015, 25, 2034–2039. [Google Scholar] [CrossRef] [Green Version]
- Wein, T.; Hülter, N.F.; Mizrahi, I.; Dagan, T. Emergence of plasmid stability under non-selective conditions maintains antibiotic resistance. Nature Comm. 2019, 10, 2595. [Google Scholar] [CrossRef] [Green Version]
- Carroll, A.C.; Wong, A. Plasmid persistence: Costs, benefits, and the plasmid paradox. Can. J. Microbiol. 2018, 64, 293–304. [Google Scholar] [CrossRef]
- Stewart, V. Regulation of nitrate and nitrite reductase synthesis in enterobacteria. Antonie Leeuwenhoek 1994, 66, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Pfreundt, U.; Stal, L.J.; Voß, B.; Hess, W.R. Dinitrogen fixation in a unicellular chlorophyll d-containing cyanobacterium. ISME J. 2012, 6, 1367–1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shih, P.M.; Wu, D.; Latifi, A.; Axen, S.; Fewer, D.; Talla, E.; Calteau, A.; Cai, F.; de Marsac, N.T.; Rippka, R.; et al. Improving the coverage of the cyanobacterial phylum using diversity-driven genome sequencing. Proc. Natl. Acad. Sci. USA 2013, 110, 1053–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watzer, B.; Forchhammer, K. Cyanophycin synthesis optimizes nitrogen utilization in the unicellular cyanobacterium Synechocystis sp. strain PCC 6803. Appl. Environ. Microbiol. 2018, 84, e01298-18. [Google Scholar] [CrossRef] [Green Version]
- Boussiba, S.; Richmond, A.E. C-phycocyanin as a storage protein in the blue-green alga Spirulina platensis. Arch. Microbiol. 1980, 125, 143–147. [Google Scholar] [CrossRef]
- Glazer, A.N. Phycobiliproteins. Methods Enzymol. 1988, 167, 291–303. [Google Scholar]
- Allen, M.M.; Smith, A.J. Nitrogen chlorosis in blue-green algae. Arch. Microbiol. 1969, 69, 114–120. [Google Scholar] [CrossRef]
- Russell, G. Salinity and seaweed vegetation. In Plant Life in Aquatic and Amphibious Habitats; Crawford, R.M.M., Ed.; Blackwell: Oxford, UK, 1987; pp. 35–52. [Google Scholar]
- Karsten, U. Seaweed acclimation to salinity and desiccation stress. In Seaweed Biology; Wiencke, C., Bischof, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 87–107. [Google Scholar]
- Hagemann, M. Molecular biology of cyanobacterial salt acclimation. FEMS Microbiol. Rev. 2011, 35, 87–123. [Google Scholar] [CrossRef]
- Fukaya, F.; Promden, W.; Hibino, T.; Tanaka, Y.; Nakamura, T.; Takabe, T. An Mrp-like cluster in the halotolerant cyanobacterium Aphanothece halophytica functions as a Na+/H+ antiporter. Appl. Environ. Microbiol. 2009, 75, 6626–6629. [Google Scholar] [CrossRef] [Green Version]
- Blanco-Rivero, A.; Leganes, F.; Fernandez-Valiente, E.; Calle, P.; Fernandez-Piñas, F. mrpA, a gene with roles in resistance to Na+ and adaptation to alkaline pH in the cyanobacterium Anabaena sp. PCC7120. Microbiology 2005, 151, 1671–1682. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, T.; Bremer, E. Guardians in a stressful world: The Opu family of compatible solute transporters from Bacillus subtilis. Biol. Chem. 2017, 398, 193–214. [Google Scholar] [CrossRef] [PubMed]
- Emerson, D. The irony of iron–biogenic iron oxides as an iron source to the ocean. Front. Microbiol. 2016, 6, 1502. [Google Scholar] [CrossRef] [PubMed]
- Kranzler, C.; Rudolf, M.; Keren, N.; Schleiff, E. Iron in cyanobacteria. Adv. Bot. Res. 2013, 65, 57–105. [Google Scholar]
- Waterworth, S.C.; Isemonger, E.W.; Rees, E.R.; Dorrington, R.A.; Kwan, J.C. Conserved bacterial genomes from two geographically isolated peritidal stromatolite formations shed light on potential functional guilds. Environ. Microbiol. Rep. 2021, 13, 126–137. [Google Scholar] [CrossRef]
- Gallagher, A.L.; Miller, S.R. Expression of novel gene content drives adaptation to low iron in the cyanobacterium Acaryochloris. Genome Biol. Evol. 2018, 10, 1484–1492. [Google Scholar] [CrossRef] [Green Version]
- Jeanjean, R.; Talla, E.; Latifi, A.; Havaux, M.; Janicki, A.; Zhang, C.C. A large gene cluster encoding peptide synthetases and polyketide synthases is involved in production of siderophores and oxidative stress response in the cyanobacterium Anabaena sp. strain PCC 7120. Environ. Microbiol. 2008, 10, 2574–2585. [Google Scholar] [CrossRef]
- Wheatley, R.M.; MacLean, R.C. CRISPR-Cas systems restrict horizontal gene transfer in Pseudomonas aeruginosa. ISME J. 2021, 15, 1420–1433. [Google Scholar] [CrossRef]
- Jiang, W.; Maniv, I.; Arain, F.; Wang, Y.; Levin, B.R.; Marraffini, L.A. Dealing with the evolutionary downside of CRISPR immunity: Bacteria and beneficial plasmids. PLoS Genet. 2013, 9, e1003844. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Type | Repeat Consensus Sequence |
|---|---|---|
| MU08, MU09 | I-D | GTTGCAAAAACGCTAAAACCCTCcAAGGGATTGAAAC |
| WB4, FH6, FH1, NIES2412 | I-E | GTTGTCCCCACGCCTGTGGGGGTGGTCCG |
| HP10 | I-MYXAN | AGCGGTGATTTAAGGTTTCCGGCCTGAAGCTTTGATGGACTT |
| MU08 | III-A | GTTTCATCACTCATTCCCCGCAAGGGGACGGAAAC |
| HP6 | III-B | GTTTCCAATAATTCCGATTGAAGTCAATCGGTAAAG |
| MU04, HP6, MU13, FH2 | III-B | GTTTCCAATAATTCCGATTGAAGTCAATCGGTAAAG |
| HP11, FH6 | III-B | GTTTTCATTTATTCGCCTTCCTACTGAATAGGAAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miller, S.R.; Abresch, H.E.; Baroch, J.J.; Fishman Miller, C.K.; Garber, A.I.; Oman, A.R.; Ulrich, N.J. Genomic and Functional Variation of the Chlorophyll d-Producing Cyanobacterium Acaryochloris marina. Microorganisms 2022, 10, 569. https://doi.org/10.3390/microorganisms10030569
Miller SR, Abresch HE, Baroch JJ, Fishman Miller CK, Garber AI, Oman AR, Ulrich NJ. Genomic and Functional Variation of the Chlorophyll d-Producing Cyanobacterium Acaryochloris marina. Microorganisms. 2022; 10(3):569. https://doi.org/10.3390/microorganisms10030569
Chicago/Turabian StyleMiller, Scott R., Heidi E. Abresch, Jacob J. Baroch, Caleb K. Fishman Miller, Arkadiy I. Garber, Andrew R. Oman, and Nikea J. Ulrich. 2022. "Genomic and Functional Variation of the Chlorophyll d-Producing Cyanobacterium Acaryochloris marina" Microorganisms 10, no. 3: 569. https://doi.org/10.3390/microorganisms10030569
APA StyleMiller, S. R., Abresch, H. E., Baroch, J. J., Fishman Miller, C. K., Garber, A. I., Oman, A. R., & Ulrich, N. J. (2022). Genomic and Functional Variation of the Chlorophyll d-Producing Cyanobacterium Acaryochloris marina. Microorganisms, 10(3), 569. https://doi.org/10.3390/microorganisms10030569