
Phytoplankton Community Structure in Highly-Mineralized Small Gypsum Karst Lake (Russia)

Abstract
1. Introduction
2. Materials and Methods
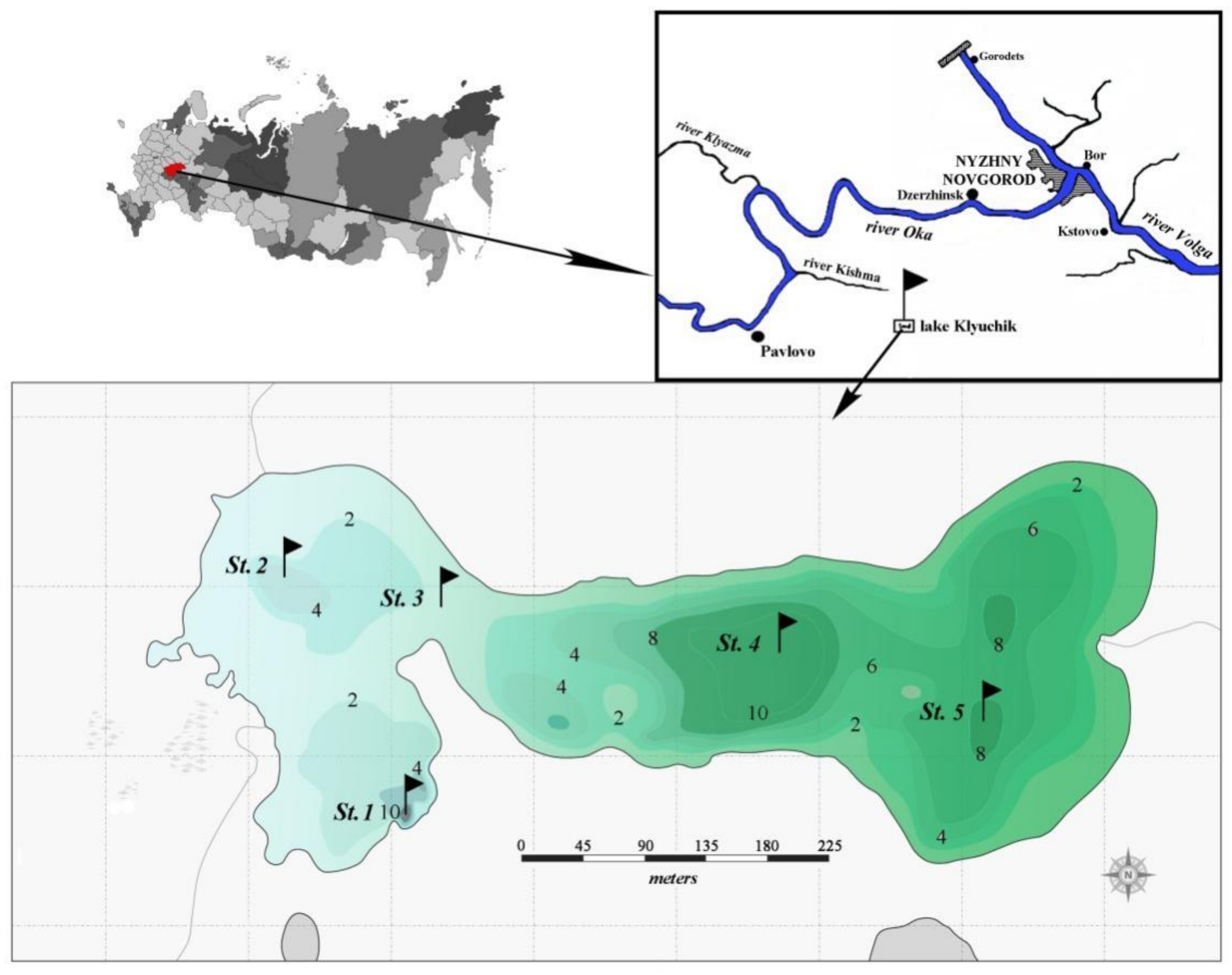
2.1. Study Area and Sampling
2.2. Data Analysis
2.3. Statistical Analysis
3. Results
3.1. Environmental Conditions
3.2. Phytoplankton Community Composition
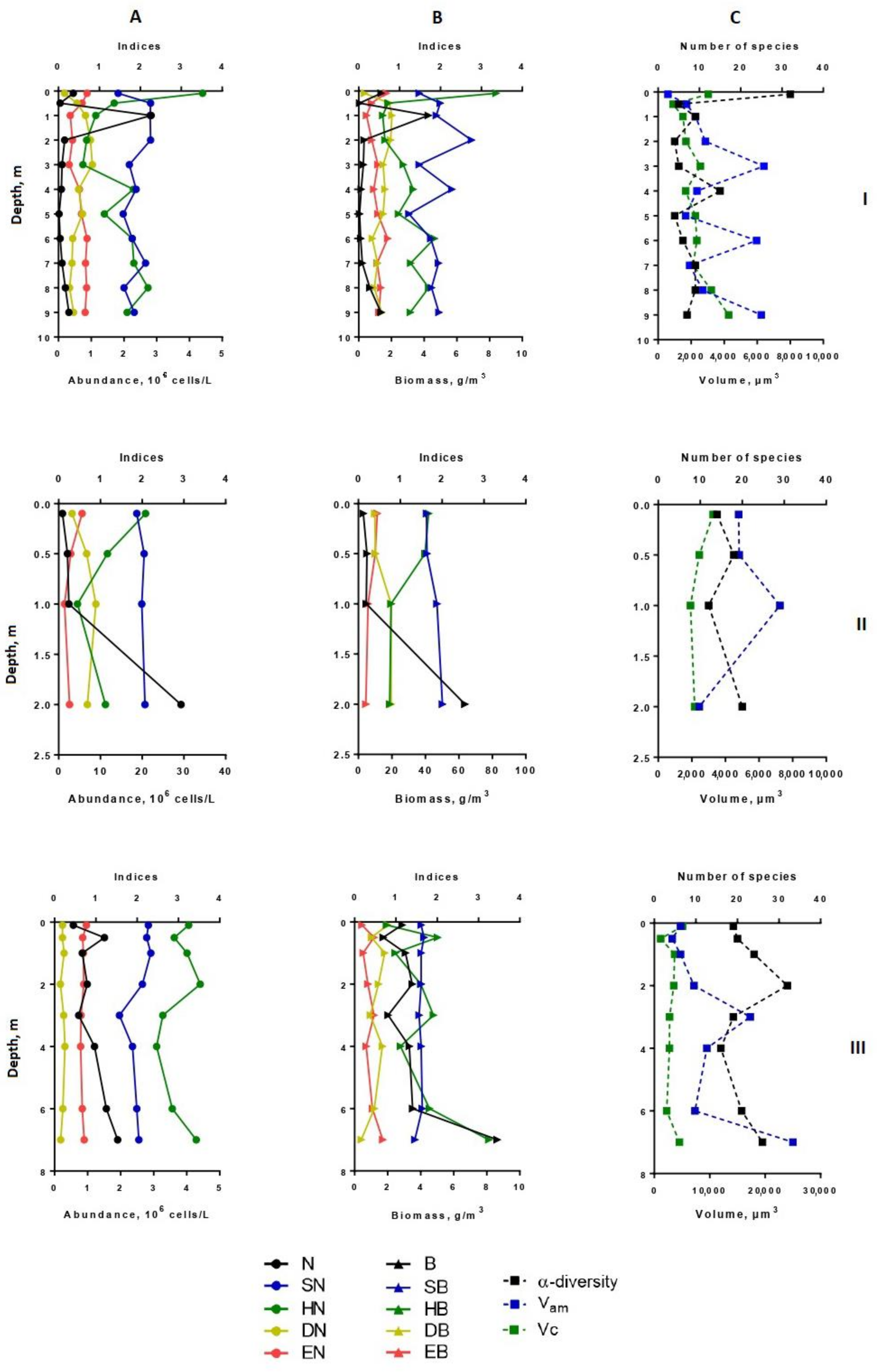
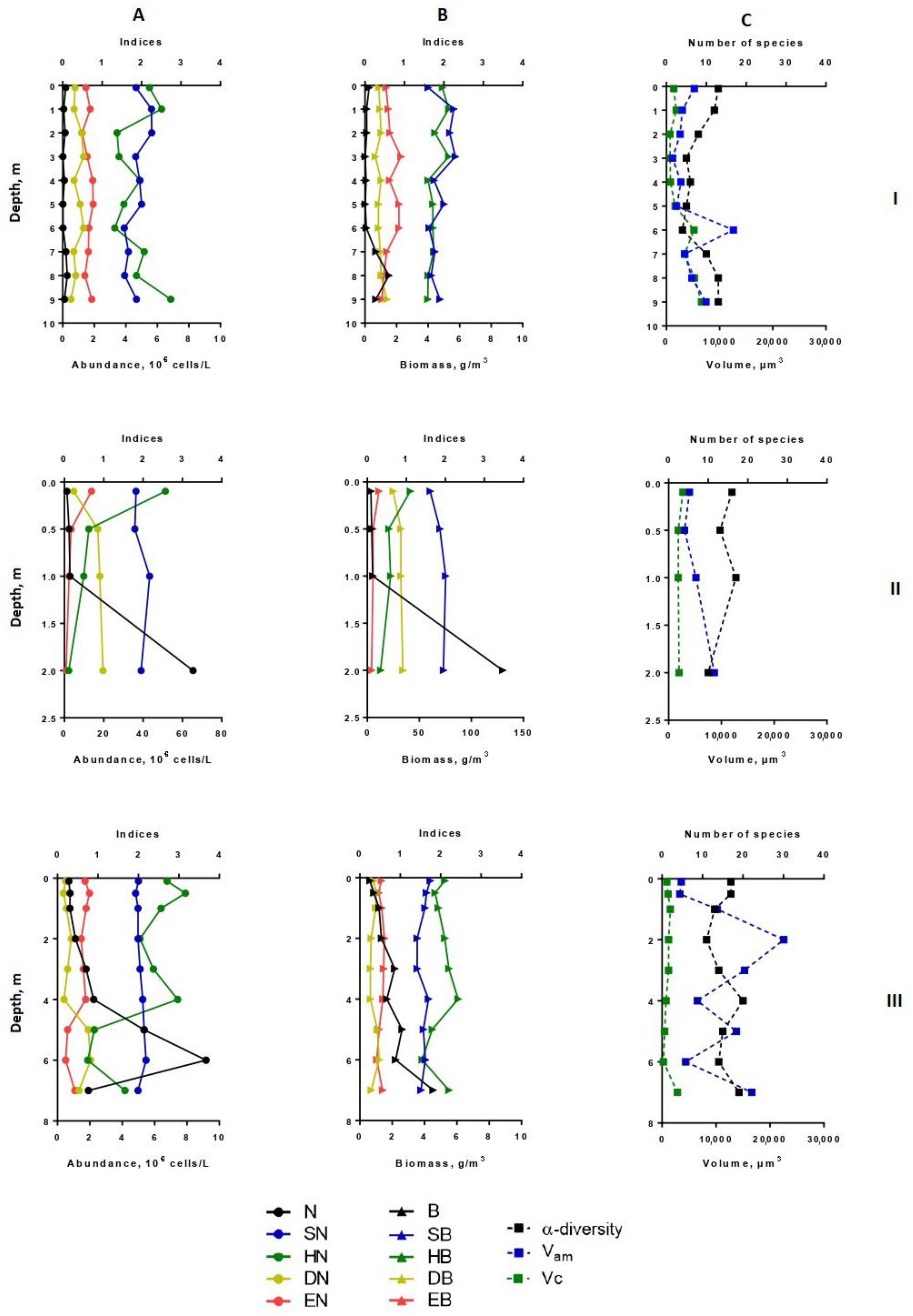
3.3. The Spatial Distribution of Phytoplankton Abundance, Biomass, Diversity Indices, Size Structure
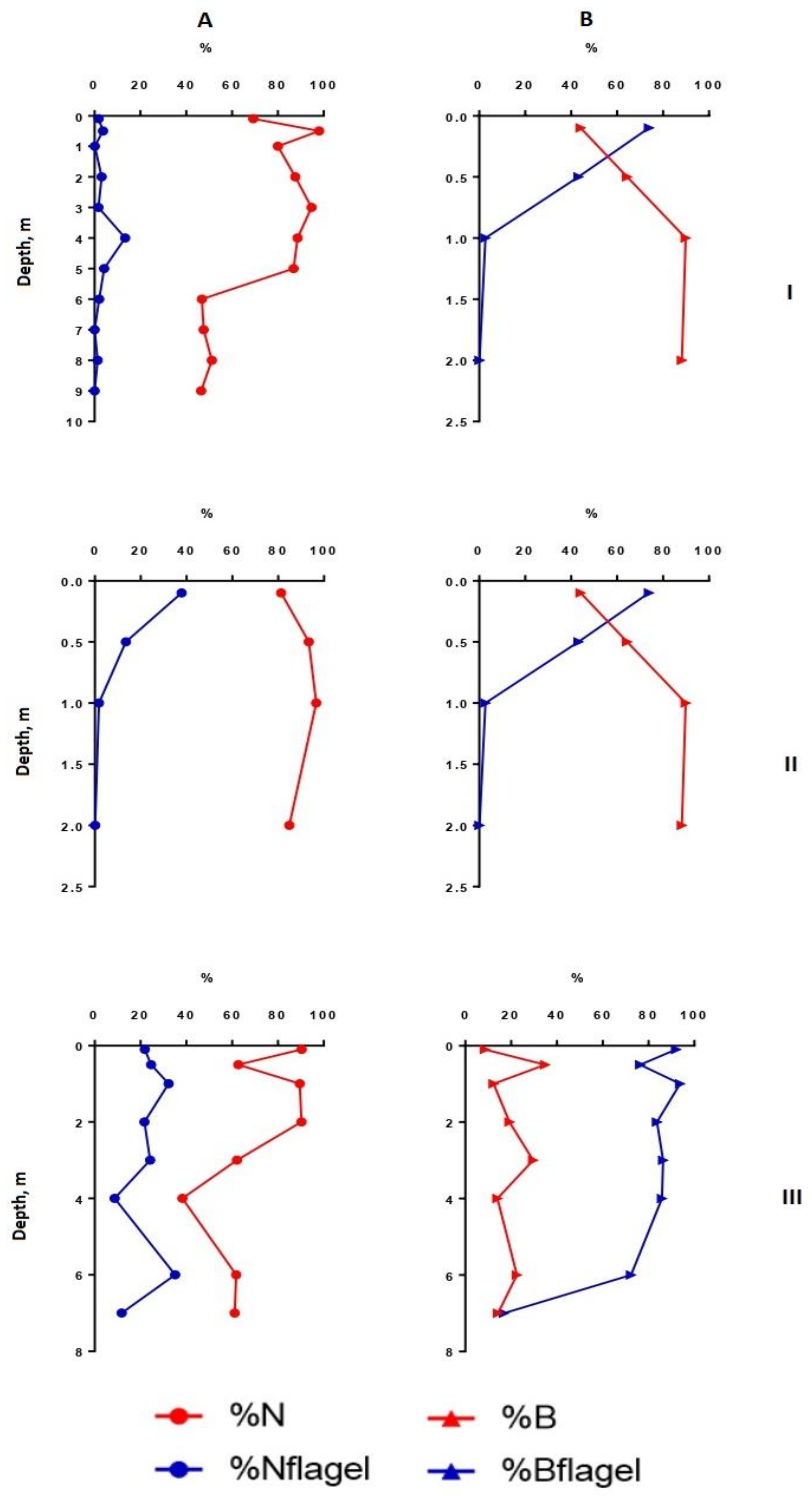
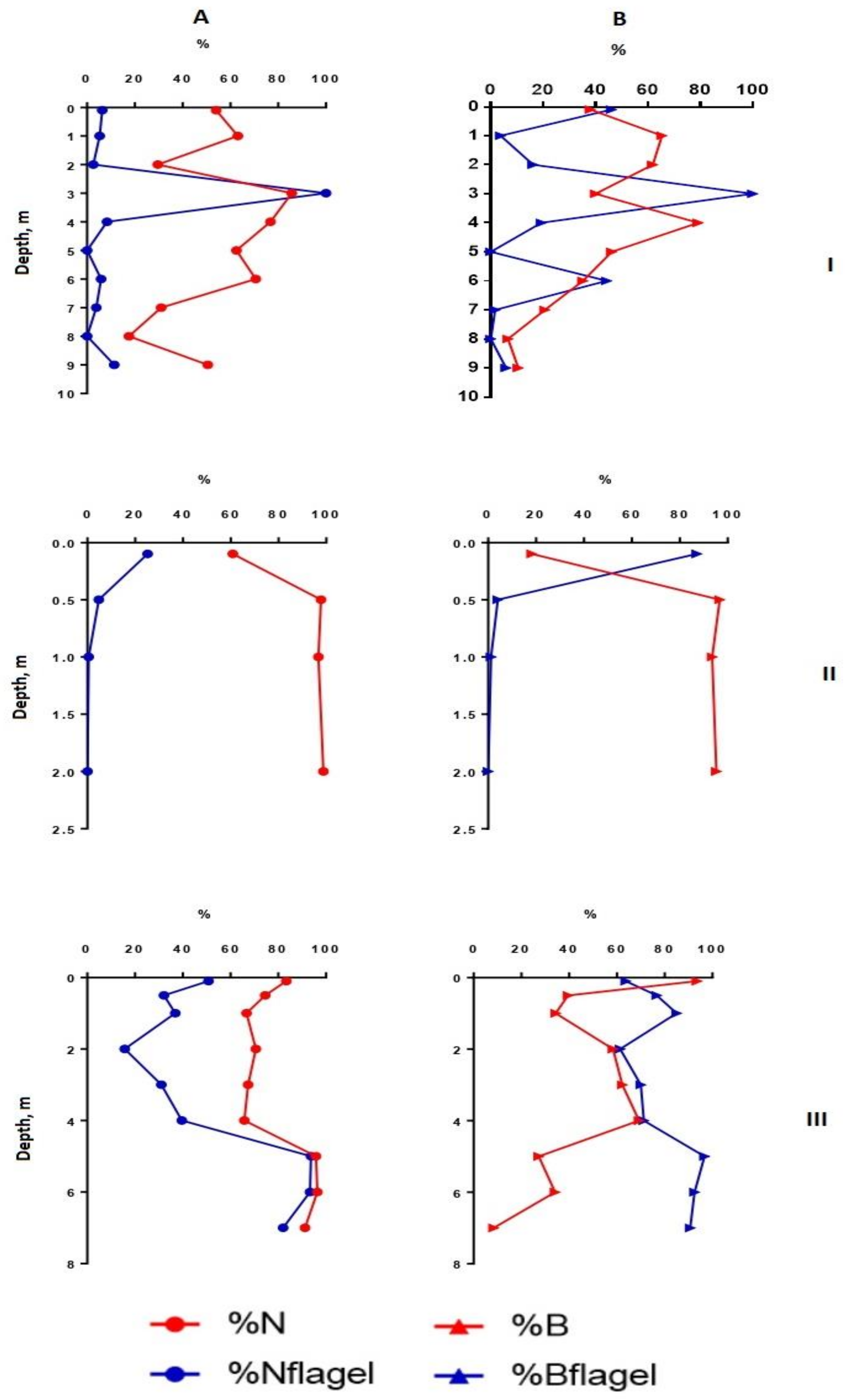
3.4. The Vertical Distribution of Phytoplankton Abundance, Biomass, Dominant Species, Diversity Indices, Size Structure
3.5. Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ryanzhin, S.V. Size distribution of world lakes and rivers derived from WORLDLAKE database. In Proceedings of the 4th International Lake Ladoga Symposium, Saint-Petersburg, Russia, 2–6 September 2002; pp. 435–441. [Google Scholar]
- Ciorca, A.M.; Momeu, L.; Battes, K.P. Same karstic substratum, different aquatic communities? Case study: Three water bodies from western Romania. Studia Univ. Babeş Bolyai Biol. 2017, 62, 67–85. [Google Scholar] [CrossRef]
- Maksimovich, G.A. Basics of the Karst Studies; Perm Book Publishing House: Perm, Russia, 1963; Volume 1, p. 444. (In Russian) [Google Scholar]
- Krevs, A.; Kucinskiene, A. Vertical distribution of bacteria and intensity of microbiological processes in two stratified gypsum Karst Lakes in Lithuania. Knowl. Manag. Aquat. Ecosyst. 2011, 4, 1–12. [Google Scholar] [CrossRef][Green Version]
- Chalkia, E.; Zacharias, I.; Thomatou, A.A.; Kehayias, G. Zooplankton dynamics in a gypsum karst lake and interrelation with the abiotic environment. Biologia 2012, 67, 151–163. [Google Scholar] [CrossRef]
- Alimova, A.F.; Mingazova, N.M. (Eds.) Unique Ecosystems of Brackish-Water Karst Lakes in the Middle Volga Region; Kazan University Publishing House: Kazan, Russia, 2001; p. 256. (In Russian) [Google Scholar]
- Klimchouk, A.; Fotri, P.; Cooper, A. Gypsum karst of the world: A brief overview. Int. J. Speleol. 1996, 25, 159–181. [Google Scholar] [CrossRef]
- Šulčius, S.; Alzbutas, G.; Juknevičiūtė, V.; Šimoliūnas, E.; Venckus, P.; Šimoliūnienė, M.; Paškauskas, R. Exploring viral diversity in a Gypsum Karst lake ecosystem using targeted single-cell genomics. Genes 2021, 12, 886. [Google Scholar] [CrossRef]
- Stachura, M.; Wieczorek, D.; Zieliński, A. An attempt at a typology of karst lakes in the Połaniec Basin (Małopolska Upland). Bull. Geography. Phys. Geogr. Ser. 2018, 15, 63–74. [Google Scholar] [CrossRef]
- Palagushkina, O.V. Ecology of Phytoplankton of Karst Lakes in the Middle Volga Region. Extended Abstract of Cand. Sci. (Biol.). PhD Thesis, Kazan State University, Kazan, Russia, 14 December 2004; p. 25. (In Russian). [Google Scholar]
- Gusev, E.S. Phytoplankton primary production in several karst lakes in central Russia. Inland Water Biol. 2008, 1, 356–361. [Google Scholar] [CrossRef]
- Danielidis, D.B.; Spartinou, M.; Economou-Amilli, A. Limnological survey of Lake Amvrakia, western Greece. Hydrobiologia 1996, 318, 207–218. [Google Scholar] [CrossRef]
- Meng, F.; Li, Z.; Li, L.; Lu, F.; Liu, Y.; Lu, H.; Fan, Y. Phytoplankton alpha diversity indices response the trophic state variation in hydrologically connected aquatic habitats in the Harbin Section of the Songhua River. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Sevindik, T.O.; Altundal, E.; Kucuk, F. The seasonal and spatial distribution of the phytoplankton of a Mesotrophic Lake related to certain physical and chemical parameters. Ekoloji 2015, 24, 14–23. [Google Scholar] [CrossRef]
- Pollingher, U. Effects of latitude on phytoplankton composition and abundance in large lakes. In Large Lakes. Brock; Tilzer, M.M., Serruya, C., Eds.; Springer Series in Contemporary Bioscience; Springer: Berlin/Heidelberg, Germany, 1990; pp. 368–402. [Google Scholar] [CrossRef]
- Udovič, M.G.; Cvetkoska, A.; Žutinić, P.; Bosak, S.; Stanković, I.; Špoljarić, I.; Mršić, G.; Kralj Borojević, K.; Ćukurin, A.; Plenković-Morja, A. Defining centric diatoms of most relevant phytoplankton functional groups in deep karst lakes. Hydrobiologia 2017, 788, 169–191. [Google Scholar] [CrossRef]
- Tarasova, N.G. The composition of the algal flora of the plankton of Lake Goluboe (Samara region). Samara Luka Probl. Reg. Glob. Ecol. 2010, 19, 157–161. (In Russian) [Google Scholar]
- Valadez, F.; Rosiles-González, G.; Almazán-Becerril, A.; Merino-Ibarra, M. Planktonic cyanobacteria of the tropical karstic lake Lagartos from the Yucatan Peninsula, Mexico. Rev. Biol. Trop. 2013, 61, 971–979. [Google Scholar] [CrossRef]
- Miracle, M.R.; Vicente, E.; Pedrós-Alió, C. Biological studies of Spanish meromictic and stratified karstic lakes. Limnetica 1992, 8, 59–77. [Google Scholar] [CrossRef]
- Petar, Ž.; Marija, G.U.; Koraljka, K.B.; Anđelka, P.M.; Judit, P. Morpho-functional classifications of phytoplankton assemblages of two deep karstic lakes. Hydrobiologia 2014, 740, 147–166. [Google Scholar] [CrossRef]
- Momeu, L.; Ciorca, A.; László, O.T.; Segedi, C.; Battes, K.P.; Cîmpean, M. The karstic lake Iezerul Ighiel (Transylvania, Romania): Its first limnological study. Studia Univ. Babes Bolyai Biol. LX 2015, 2, 39–60. [Google Scholar]
- Korneva, L.G. Diversity and Structure of Phytoplankton of Some Low-Mineralized Forest Lakes in the Vologda Oblast, in Gidrobiologicheskie Voprosy (Hydrobiological Issues); Yakutsk University: Yakutsk, Russia, 2000; p. 94, Part 2. (In Russian) [Google Scholar]
- Okhapkin, A.G. Dynamics of the species structure of potamophytoplankton in watercourses of different types. Inland Water Biol. 2000, 1, 53–61. (In Russian) [Google Scholar]
- Okhapkin, A.G.; Startseva, N.A. Dynamics of species structure of phytoplankton in small waterbodies of urbanized territories: Species diversity and size structure of communities. Inland Water Biol. 2005, 2, 29–33. (In Russian) [Google Scholar]
- Korneva, L.G. Phytoplankton of Volga River Basin Reservoirs; Kostroma Printing House: Kostroma, Russia, 2015; p. 284. (In Russian) [Google Scholar]
- Alimov, A.F. Elements of Aquatic Ecosystem Function Theory; Nauka: Saint Petersburg, Russia, 2000; p. 147. (In Russian) [Google Scholar]
- Carvalho, L.; Poikane, S.; Solheim, A.L.; Phillips, G.; Borics, G.; Catalan, J.; de Hoyos, C.; Drakare, S.; Dudley, B.J.; Järvinen, M.; et al. Strength and uncertainty of phytoplankton metrics for assessing eutrophication impacts in lakes. Hydrobiologia 2013, 704, 127–140. [Google Scholar] [CrossRef]
- Borics, G.; Abonyi, A.; Salmaso, N.; Ptacnik, R. Freshwater phytoplankton diversity: Models, drivers and implications for ecosystem properties. Hydrobiologia 2021, 848, 53–75. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, A.V.; Tarasov, I.A.; Dedyk, V.E. Ecological and hydrochemical characteristics of the water area of the Klyuchik Lake in Pavlovsky district of the Nizhny Novgorod region. Mod. Probl. Sci. Educ. 2017, 1, 126–134. (In Russian) [Google Scholar]
- Bayanov, N.G. Klyuchik Lake is a unique natural object of the Nizhny Novgorod Volga region. Work. State Nat. Biosph. «Kerzhnesky» 2019, 9, 65–72. (In Russian) [Google Scholar]
- Bakka, S.V.; Kiseleva, N.Y. Specially Protected Natural Areas of the Nizhny Novgorod Region; Ministry of Ecology Natural Resources: Nizhny Novgorod, Russia, 2009; p. 560. [Google Scholar]
- Vodeneeva, E.L.; Okhapkin, A.G.; Genkal, S.I.; Kulizin, P.V.; Sharagina, E.M.; Skamejkina, K.O. Composition, structure and distribution of phytoplankton of a highly mineralized karst lake. Inland Water Biol. 2020, 13, 573–582. [Google Scholar] [CrossRef]
- Genkal, S.I.; Okhapkin, A.G.; Vodeneeva, E.L. On the morphology and taxonomy of Cyclotella distinguenda (Bacillariophyta). Nov. Sist. Nizsh. Rast. 2019, 5, 247–253. [Google Scholar] [CrossRef]
- Petrov, M.S.; Astashin, A.E. Dynamics of the consumption of water in the biggest in the Nizhny Novgorod region spring Surin. Orfanov readings—2017. In Proceedings of the Materials All-Russian Scientific and Practical Conference Dedicated to the 80th Anniversary of Higher Geographical Education in the Nizhny Novgorod Region and the 70th Anniversary of the Nizhny Novgorod Branch of the All-Russian Public Organization Russian Geographical Society, Minin University, Nizhny Novgorod, Russia, 24–25 November 2017; pp. 29–32. (In Russian). [Google Scholar]
- Bakanina, F.M.; Vorotnikov, V.P.; Lukina, E.V.; Fridman, B.I. Lakes of the Nizhny Novgorod Region; VOOP Publishing House: Nizhny Novgorod, Russia, 2001; p. 165. (In Russian) [Google Scholar]
- Okhapkin, A.G.; Genkal, S.I.; Scharagina, E.M.; Vodeneeva, E.L. Structure and dynamics of phytoplankton in the Oka river mouth at the beginning of the 21th century. Inland Water Biol. 2014, 7, 357–365. [Google Scholar] [CrossRef]
- Sun, J.; Liu, D. Geometric models for calculating cell biovolume and surface area for phytoplankton. J. Plankton Res. 2003, 25, 1331–1346. [Google Scholar] [CrossRef]
- Hillebrand, H.; Dürselen, C.D.; Kirschtel, D.; Pollingher, D.; Zohary, T. Biovolume calculation for pelagic and benthic microalgae. J. Phycol. 1999, 35, 403–424. [Google Scholar] [CrossRef]
- Abakumov, V.A. (Ed.) Guide on Methods for Hydrobiological Analysis of Surface Waters and Bottom Sediments; Gidrometeoizdat: Leningrad, Russia, 1983; p. 239. (In Russian) [Google Scholar]
- USSR Science Academy. Methods for Studying Biogeocenoses of Inland Water Bodies; Morduhai-Boltovskoi, F.D., Ed.; Nauka: Moscow, Russia, 1975; p. 240. (In Russian) [Google Scholar]
- Malashenkov, D.V.; Dashkova, V.; Zhakupova, K.; Vorobjev, I.A.; Barteneva, N.S. Comparative analysis of freshwater phytoplankton communities in two lakes of Burabay National Park using morphological and molecular approaches. Sci. Rep. 2021, 11, 16130. [Google Scholar] [CrossRef]
- Vodeneeva, E.L.; Kulizin, P.V. Algae of the Mordovian Nature Reserve. Annotated List of Species; Flora and Fauna of Nature Reserves: Moscow, Russia, 2019; Volume 134, p. 62. (In Russian) [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication; National University of Ireland: Galway, Ireland, 2021; Available online: http://www.algaebase.org (accessed on 29 July 2021).
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; The University of Illinois Press: Urbana, IL, USA, 1949; p. 117. [Google Scholar]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Sládeček, V. System of water quality from biological point of view. Arch. Hydrobiol. Erg. Limnol. 1973, 7, 218. [Google Scholar]
- Reynolds, C.S. Ecology of Phytoplankton; Cambrige University Press: New York, NY, USA, 2006; p. 535. [Google Scholar]
- Okhapkin, A.G. Species composition of phytoplankton as an indicator of living conditions in streams of different types. Bot. Zh. 1998, 83, 1–13. (In Russian) [Google Scholar]
- Mann, H.B.; Whitney, D.R. On a test of whether one of two random variables is stochastically larger than the other. Ann. Math. Stat. 1947, 18, 50–60. [Google Scholar] [CrossRef]
- Stevens, L.E.; Schenk, E.R.; Springer, A.E. Springs ecosystem classification. Ecol. Appl. 2021, 31, e2218. [Google Scholar] [CrossRef]
- Kitaev, S.P. Basics of Limnology for Hydrobiologists and Ichthyologists; Karelian Science Center Press RAN: Petrozavodsk, Russia, 2007; p. 394. (In Russian) [Google Scholar]
- Bigon, M.; Harper, D.; Tausend, K. Ecology. Individuals, populations and communities; Mir Publishers: Moscow, Russia, 1989; Volume 2, p. 477. (In Russian) [Google Scholar]
- Weckström, J.; Korhola, A.; Blom, T. Diatoms as quantitative indicators of ph and water temperature in subarctic Fennoscandian lakes. Hydrobiologia 1997, 347, 171–184. [Google Scholar] [CrossRef]
- Okhapkin, A.G. Phytoplankton of the system of Pustynskie Lakes of the Gorky Region. In Biological Bases for Increasing the Productivity and Protection of Plant Communities in the Volga Region; Gorky State University Press: Gorky, Russia, 1981; pp. 106–109. (In Russian) [Google Scholar]
- Gusev, E.S. Vertical Distribution of Phytoplankton in Small Karst Lakes in Central Russia. Algae: Taxonomy, Ecology, Use in Monitoring; Ural Branch of the Russian Academy of Sciences: Ekaterinburg, Russia, 2011; pp. 160–167. (In Russian) [Google Scholar]
- Odum, U. Ecology; Mir Publishing: Moscow, Russia, 1986; Volume 1, p. 328. (In Russian) [Google Scholar]
- Magarran, E. Ecological Diversity and Its Measurement; Mir Publishing: Moscow, Russia, 1992; p. 181. (In Russian) [Google Scholar]
- Korneva, L.G. Impact of acidification on structural organization of phytoplankton community in the forest lakes of the north-western Russia. Water Sci. Technol. 1996, 33, 291–296. [Google Scholar] [CrossRef]
- Lessmann, D.; Fyson, A.; Nixdorf, B. Experimental eutrophication of a shallow acidic mining lake and effects on the phytoplankton. Hydrobiologia 2003, 506, 753–758. [Google Scholar] [CrossRef]
- Okhapkin, A.G.; Vodeneeva, E.L.; Yulova, G.A. Phytoplankton of reservoirs of the “Kerzhensky” nature reserve (Nizhny Novgorod region). Bot. Zh. 2004, 89, 1264–1275. [Google Scholar]
- Okhapkin, A.G.; Sharagina, E.M.; Bondarev, O.O. Phytoplankton of the Cheboksary reservoir at the present state of its existence. Povolzhskiy J. Ecol. 2013, 2, 190–199. (In Russian) [Google Scholar]
- Kiss, K.T.; Klee, R.; Ector, L.; Ács, É. Centric diatoms of large rivers and tributaries in Hungary: Morphology and biogeographic distribution. Acta Bot. Croat. 2012, 71, 311–363. [Google Scholar] [CrossRef]
- Padisák, J.; Crossetti, L.; Naselli-Flores, L. Use and misuse in the application of the phytoplankton functional classification: A critical review with updates. Hydrobiologia 2009, 621, 1–19. [Google Scholar] [CrossRef]
- Dokulil, M.T.; Teubner, K. Cyanobacterial dominance in lakes. Hydrobiologia 2000, 438, 1–12. [Google Scholar] [CrossRef]
- Downing, J.A.; Watson, S.B.; McCauley, E. Predicting cyanobacteria dominance in lakes. Can. J. Fish. Aquat. Sci. 2001, 58, 1905–1908. [Google Scholar] [CrossRef]
- Bouterfas, R.; Belkoura, M.; Dauta, A. Light and temperature effects on the growth rate of the three freshwater algae isolated from a eutrophic lake. Hydrobiologia 2002, 489, 207–217. [Google Scholar] [CrossRef]
- Padisák, J. Phytoplankton. The Lakes Handbook. Limnology and Limnetic Ecology; O’Sullivan, P.E., Reynolds, C.S., Eds.; Blackwell Publishing: Hoboken, NJ, USA, 2004; Volume 1, pp. 251–308. [Google Scholar]
- Trifonova, I.S. Ecology and Succession of Lake Phytoplankton; Science: Leningrad, Russia, 1990; p. 183. (In Russian) [Google Scholar]
- Rottberger, J.; Gruber, A.; Boenigk, J.; Kroth, P.G. Influence of nutrients and light on autotrophic, mixotrophic and heterotrophic freshwater chrysophytes. Aquat. Microb. Ecol. 2013, 71, 179–191. [Google Scholar] [CrossRef]
- Saad, J.F.; Unrein, F.; Tribelli, P.M.; López, N.; Izaguirre, I. Influence of lake trophic conditions on the dominant mixotrophic algal assemblages. J. Plankton Res. 2016, 38, 818–829. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | St. 1 | St. 2 | St. 3 | St. 4 | St. 5 |
|---|---|---|---|---|---|
| Depth (m) | 10.7 | 3.0 | 2.0 | 9.4 | 7.0 |
| Transparency (m) | 8.0–8.5 | to the bottom | to the bottom | 3.4–5.4 | 3.0–4.5 |
| Temperature (°C) | 7.5–9.5 8.8 ± 0.9 | 8.0–10.5 9.2 ± 0.7 | 8.1–13.6 10.6 ± 1.6 | 11.0– 15.8 13.8 ± 1.3 | 13.2–17.3 15.6±1.2 |
| pH | 7.0–7.1 7.1 ± 0.04 | 7.1–7.2 7.1 ± 0.03 | 7.1–7.6 7.3 ± 0.16 | 7.3–7.4 7.4 ± 0.02 | 7.4–7.6 7.5 ± 0.06 |
| Structural Variables | St. 1 | St. 2 | St. 3 | St. 4 | St. 5 | U Criterion St. 1 × St. 5 |
|---|---|---|---|---|---|---|
| N | 0.31 ± 0.07 | 3.57 ± 2.10 | 25.80 ± 11.40 | 1.90 ± 0.41 | 1.53 ± 0.65 | |
| B | 0.94 ± 0.12 | 11.20 ± 6.00 | 55.50 ± 21.90 | 8.32 ± 0.48 | 6.82 ± 2.96 | p ≤ 0.05 |
| SN | 1.59 ± 0.04 | 1.54 ± 0.02 | 1.94 ± 0.20 | 1.52 ± 0.01 | 1.64 ± 0.07 | |
| SB | 1.55 ± 0.04 | 1.75 ± 0.13 | 1.87 ± 0.05 | 1.62 ± 0.07 | 1.49 ± 0.08 | |
| HN | 2.25 ± 0.46 | 1.56 ± 0.21 | 1.01 ± 0.56 | 2.83 ± 0.02 | 2.22 ± 0.07 | |
| HB | 2.19 ± 0.42 | 1.61 ± 0.23 | 1.31 ± 0.70 | 3.31 ± 0.17 | 2.23 ± 0.18 | |
| DN | 0.37 ± 0.10 | 0.47 ± 0.04 | 0.71 ± 0.16 | 0.22 ± 0.01 | 0.34 ± 0.03 | |
| DB | 0.36 ± 0.11 | 0.50 ± 0.08 | 0.64 ± 0.20 | 0.14 ± 0.02 | 0.36 ± 0.06 | |
| EN | 0.58 ± 0.06 | 0.38 ± 0.04 | 0.21 ± 0.10 | 0.66 ± 0.01 | 0.51 ± 0.03 | |
| EB | 0.57 ± 0.09 | 0.40 ± 0.06 | 0.27 ± 0.13 | 0.77 ± 0.02 | 0.53 ± 0.07 | |
| %N | 74.00 ± 3.50 | 94.60 ± 1.60 | 91.60 ± 3.70 | 74.5 ± 2.60 | 73.40 ± 7.20 | p ≤ 0.05 |
| %B | 59.80 ± 8.50 | 79.70 ± 9.40 | 82.80 ± 9.00 | 38.80 ± 5.10 | 44.60 ± 18.50 | p ≤ 0.05 |
| %Nflagel | 0.60 ± 0.30 | 1.10 ± 0.20 | 0.20 ± 0.10 | 8.40 ± 5.90 | 11.30 ± 4.70 | p ≤ 0.05 |
| %Bflagel | 0.40 ± 0.30 | 6.10 ± 3.70 | 1.00 ± 0.90 | 0.90 ± 0.70 | 14.30 ± 12.30 | p ≤ 0.05 |
| SDN | 1.70 ± 0.30 | 2.00 ± 0.00 | 1.30 ± 0.30 | 3.00 ± 0.60 | 3.00 ± 0.60 | |
| SDB | 2.70 ± 0.30 | 2.30 ± 0.30 | 1.00 ± 0.00 | 2.70 ± 0.30 | 1.70 ± 0.30 | |
| SMN | 3.70 ± 0.30 | 2.30 ± 0.30 | 2.00 ± 0.60 | 3.30 ± 0.30 | 3.70 ± 0.30 | |
| SMB | 3.70 ± 0.30 | 3.70 ± 0.90 | 1.70 ± 0.30 | 6.70 ± 0.30 | 3.30 ± 0.70 | |
| Vam | 4.60 ± 1.10 | 12.50 ± 4.10 | 7.80 ± 1.90 | 16.20 ± 4.90 | 19.90 ± 4.70 | |
| Vc | 3.30 ± 0.50 | 3.30 ± 0.20 | 2.30 ± 0.50 | 4.80 ± 0.90 | 5.60 ± 1.90 | |
| Sp | 18.30 ± 5.30 | 17.30 ± 1.80 | 26.00 ± 4.60 | 20.00 ± 1.70 | 21.00 ± 3.20 |
| HB | HN | DB | DN | EB | EN | |
|---|---|---|---|---|---|---|
| HB | 1.00 | 0.88 | −0.99 | −0.89 | 0.86 | 0.88 |
| HN | 0.88 | 1.00 | −0.87 | −0.96 | 0.74 | 0.95 |
| DB | −0.99 | −0.87 | 1.00 | 0.87 | −0.88 | −0.85 |
| DN | −0.89 | −0.96 | 0.87 | 1.00 | −0.73 | −0.91 |
| EB | 0.86 | 0.74 | −0.88 | −0.73 | 1.00 | 0.78 |
| EN | 0.85 | 0.95 | −0.85 | −0.91 | 0.78 | 1.00 |
| HB | HN | DB | DN | EB | EN | |
|---|---|---|---|---|---|---|
| N | p ≥ 0.05 | p ≥ 0.05 | p ≥ 0.05 | p ≥ 0.05 | p ≥ 0.05 | −0.54 |
| SB | p ≥ 0.05 | −0.52 | p ≥ 0.05 | p ≥ 0.05 | p ≥ 0.05 | −0.55 |
| %N | −0.62 | −0.76 | 0.59 | 0.76 | −0.66 | −0.82 |
| %B | −0.71 | −0.85 | 0.73 | 0.74 | −0.67 | −0.81 |
| %Nflagel | 0.57 | 0.64 | −0.54 | −0.77 | p ≥ 0.05 | 0.61 |
| SDN | 0.70 | 0.64 | −0.65 | −0.78 | 0.56 | 0.57 |
| SMN | 0.56 | 0.77 | p ≥ 0.05 | −0.75 | p ≥ 0.05 | 0.74 |
| SDB | p≥0.05 | p ≥ 0.05 | p ≥ 0.05 | p ≥ 0.05 | p ≥ 0.05 | 0.53 |
| SMB | 0.74 | 0.67 | −0.77 | −0.74 | 0.68 | 0.76 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okhapkin, A.; Sharagina, E.; Kulizin, P.; Startseva, N.; Vodeneeva, E. Phytoplankton Community Structure in Highly-Mineralized Small Gypsum Karst Lake (Russia). Microorganisms 2022, 10, 386. https://doi.org/10.3390/microorganisms10020386
Okhapkin A, Sharagina E, Kulizin P, Startseva N, Vodeneeva E. Phytoplankton Community Structure in Highly-Mineralized Small Gypsum Karst Lake (Russia). Microorganisms. 2022; 10(2):386. https://doi.org/10.3390/microorganisms10020386
Chicago/Turabian StyleOkhapkin, Alexander, Ekaterina Sharagina, Pavel Kulizin, Natalja Startseva, and Ekaterina Vodeneeva. 2022. "Phytoplankton Community Structure in Highly-Mineralized Small Gypsum Karst Lake (Russia)" Microorganisms 10, no. 2: 386. https://doi.org/10.3390/microorganisms10020386
APA StyleOkhapkin, A., Sharagina, E., Kulizin, P., Startseva, N., & Vodeneeva, E. (2022). Phytoplankton Community Structure in Highly-Mineralized Small Gypsum Karst Lake (Russia). Microorganisms, 10(2), 386. https://doi.org/10.3390/microorganisms10020386