

Molecular Characterization of Octopamine/Tyramine Receptor Gene of Amitraz-Resistant Rhipicephalus (Boophilus) decoloratus Ticks from Uganda

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Tick Population
2.2. Extraction of Genomic DNA
2.3. Amplification of Octopamine/Tyramine Receptor Gene
2.4. Cloning and Sequencing of Octopamine/Tyramine Receptor Gene
2.5. Sequence Analysis
2.6. Ethical Consideration
3. Results
3.1. Amplification and Sequence of OCT/Tyr Receptor Gene
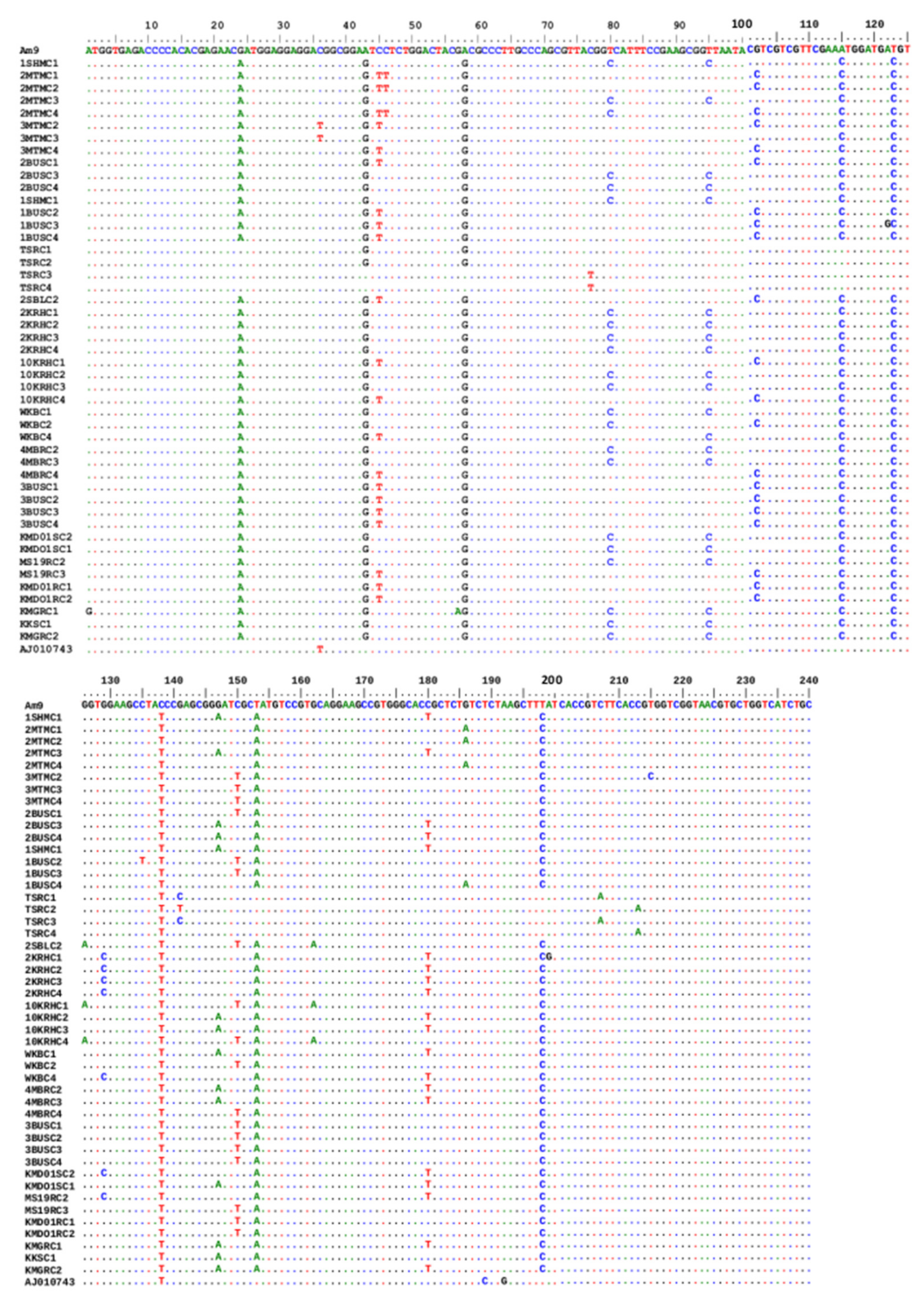
3.2. Analysis of SNPs in the Coding Region of OCT/Tyr Receptor Gene
3.3. OCT/Tyr Gene Amino Acid Mutations in Amitraz-Resistant Ticks
3.4. Evolutionary Characteristics of R. (B.) decoloratus OCT/Tyr Gene from Resistant Ticks
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Estrada-Peña, A.; Bouattour, A.; Camicas, J.-L.; Guglielmone, A.; Horak, I.; Jongejan, F.; Latif, A.; Pegram, R.; Walker, A.R. The Known Distribution and Ecological Preferences of the Tick Subgenus Boophilus (Acari: Ixodidae) in Africa and Latin America. Exp. Appl. Acarol. 2006, 38, 219–235. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.R.; Bouattour, A.; Camicas, J.L.; Estrada-Peña, A.; Horak, I.G.; Latif, A.A.; Pegram, R.G.; Preston, P.M. Ticks of Domestic Animals in Africa: A Guide to Identification of Species; University of Edinburgh: Edinburgh, UK, 2003. [Google Scholar]
- Awa, D.N.; Adakal, H.; Luogbou, N.D.D.; Wachong, K.H.; Leinyuy, I.; Achukwi, M.D. Cattle ticks in Cameroon: Is Rhipicephalus (Boophilus) microplus absent in Cameroon and the Central African region? Ticks Tick Borne Dis. 2015, 6, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Ogo, N.I.; de Mera, I.G.; Galindo, R.C.; Okubanjo, O.O.; Inuwa, H.M.; Agbede, R.I.; Torina, A.; Alongi, A.; Vicente, J.; Gortázar, C.; et al. Molecular identification of tick-borne pathogens in Nigerian ticks. Vet. Parasitol. 2012, 187, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Berggoetz, M.; Schmid, M.; Ston, D.; Wyss, V.; Chevillon, C.; Pretorius, A.-M.; Gern, L. Protozoan and bacterial pathogens in tick salivary glands in wild and domestic animal environments in South Africa. Ticks Tick-Borne Dis. 2014, 5, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Rubaire-Akiiki, C.; Okello-Onen, J.; Nasinyama, G.W.; Vaarst, M.; Kabagambe, E.K.; Mwayi, W.; Musunga, D.; Wandukwa, W. The prevalence of serum antibodies to tick-borne infections in Mbale District, Uganda: The effect of agro-ecological zone, grazing management and age of cattle. J. Insect Sci. 2004, 4, 8. [Google Scholar] [CrossRef]
- Muhanguzi, D.; Matovu, E.; Waiswa, C. Prevalence and characterization of Theileria and Babesia species in cattle under different husbandry systems in western Uganda. Int. J. Anim. Vet. Adv. 2010, 2, 51–58. [Google Scholar]
- Byaruhanga, C.; Collins, N.E.; Knobel, D.; Kabasa, W.; Oosthuizen, M.C. Endemic status of tick-borne infections and tick species diversity among transhumant zebu cattle in Karamoja Region, Uganda: Support for control approaches. Vet. Parasitol. Reg. Stud. Rep. 2015, 1–2, 21–30. [Google Scholar] [CrossRef]
- Bardosh, K.; Waiswa, C.; Welburn, S.C. Conflict of interest: Use of pyrethroids and amidines against tsetse and ticks in zoonotic sleeping sickness endemic areas of Uganda. Parasites Vectors 2013, 6, 204–215. [Google Scholar] [CrossRef]
- Vudriko, P.; Okwee-Acai, J.; Byaruhanga, J.; Tayebwa, D.S.; Okech, S.G.; Tweyongyere, R.; Wampande, E.M.; Okurut, A.R.A.; Mugabi, K.; Muhindo, J.B.; et al. Chemical tick control practices in southwestern and northwestern Uganda. Ticks Tick-Borne Dis. 2018, 9, 945–955. [Google Scholar] [CrossRef]
- Petros, A.; Befekadu, U.W.; Mulisa, M.; Teka, F. In vitro and in vivo acaricidal efficacy study of amitraz and diazinon against some tick species infesting Camelus dromedarius around Jigjiga, Eastern Ethiopia. Afr. J. Pharm. Pharmacol. 2015, 9, 850–855. [Google Scholar] [CrossRef]
- Baron, S.; van der Merwe, N.A.; Madder, M.; Maritz-Olivier, C. SNP Analysis Infers that Recombination Is Involved in the Evolution of Amitraz Resistance in Rhipicephalus microplus. PLoS ONE 2015, 10, e0131341. [Google Scholar] [CrossRef]
- Muyobela, J.; Nkunika, P.O.Y.; Mwase, E.T. Resistance status of ticks (Acari; Ixodidae) to amitraz and cypermethrin acaricides in Isoka District, Zambia. Trop. Anim. Health Prod. 2015, 47, 1599–1605. [Google Scholar] [CrossRef]
- Ahmed, M.A.I.; Vogel, C.F.A.; Matsumura, F. Unique biochemical and molecular biological mechanism of synergistic actions of formamidine compounds on selected pyrethroid and neonicotinoid insecticides on the fourth instar larvae of Aedes aegypti (Diptera: Culicidae). Pestic. Biochem. Physiol. 2015, 120, 57–63. [Google Scholar] [CrossRef]
- Del Pino, J.; Moyano-Cires, P.V.; Anadon, M.J.; Diaz, M.J.; Lobo, M.; Capo, M.A.; Frejo, M.T. Molecular Mechanisms of Amitraz Mammalian Toxicity: A Comprehensive Review of Existing Data. Chem. Res. Toxicol. 2015, 28, 1073–1094. [Google Scholar] [CrossRef]
- Gross, A.D.; Temeyer, K.B.; Day, T.A.; de León, A.A.P.; Kimber, M.J.; Coats, J.R. Pharmacological characterization of a tyramine receptor from the southern cattle tick, Rhipicephalus (Boophilus) microplus. Insect Biochem. Mol. Biol. 2015, 63, 47–53. [Google Scholar] [CrossRef]
- Ahmed, M.A.I.; Vogel, C.F.A. The role of octopamine receptor agonists in the synergistic toxicity of certain insect growth regulators (IGRs) in controlling Dengue vector Aedes aegypti (Diptera: Culicidae) mosquito. Acta Trop. 2016, 155, 1–5. [Google Scholar] [CrossRef]
- Audsley, N.; Down, R.E. G protein coupled receptors as targets for next generation pesticides. Insect Biochem. Mol. Biol. 2015, 67, 27–37. [Google Scholar] [CrossRef]
- Evans, P.D.; Maqueira, B. Insect octopamine receptors: A new classification scheme based on studies of cloned Drosophila G-protein coupled receptors. Invertebr. Neurosci. 2005, 5, 111–118. [Google Scholar] [CrossRef]
- Verlinden, H.; Vleugels, R.; Marchal, E.; Badisco, L.; Pflüger, H.J.; Blenau, W.; Broeck, J.V. The role of octopamine in locusts other arthropods. J. Insect Physiol. 2010, 56, 854–867. [Google Scholar] [CrossRef]
- Farooqui, T. Review of octopamine in insect nervous systems. Open Access Insect Physiol. 2012, 4, 1. [Google Scholar] [CrossRef]
- Cossío-Bayúgar, R.; Miranda-Miranda, E.; Fernández-Rubalcaba, M.; Padilla, V.N.; Reynaud, E. Adrenergic ligands that block oviposition in the cattle tick Rhipicephalus microplus affect ovary contraction. Sci. Rep. 2015, 5, 15109. [Google Scholar] [CrossRef] [PubMed]
- Fernndez-Salas, A.; Rodríguez-Vivas, R.I.; Alonso-Díaz, M.A. Resistance of Rhipicephalus microplus to Amitraz and Cypermethrin in Tropical Cattle Farms in Veracruz, Mexico. J. Parasitol. 2012, 98, 1010–1014. [Google Scholar] [CrossRef] [PubMed]
- Cutullé, C.; Lovis, L.; D’Agostino, B.I.; Balbiani, G.G.; Morici, G.; Citroni, D.; Reggi, J.; Caracostantogolo, J.L. In vitro diagnosis of the first case of amitraz resistance in Rhipicephalus microplus in Santo Tomé (Corrientes), Argentina. Vet. Parasitol. 2012, 192, 296–300. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Sharma, A.K.; Ray, D.D.; Ghosh, S. Determination of discriminating dose and evaluation of amitraz resistance status in different field isolates of Rhipicephalus (Boophilus) microplus in India. Exp. Appl. Acarol. 2014, 63, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.C.; He, H.; Davey, R.B. Mutations in a putative octopamine receptor gene in amitraz-resistant cattle ticks. Vet. Parasitol. 2007, 148, 379–383. [Google Scholar] [CrossRef]
- Vudriko, P.; Okwee-Acai, J.; Tayebwa, D.S.; Byaruhanga, J.; Kakooza, S.; Wampande, E.; Omara, R.; Muhindo, J.B.; Tweyongyere, R.; Owiny, D.O.; et al. Emergence of multi-acaricide resistant Rhipicephalus ticks and its implication on chemical tick control in Uganda. Parasites Vectors 2016, 9, 4. [Google Scholar] [CrossRef]
- Dzemo, W.D.; Thekisoe, O.; Vudriko, P. Development of acaricide resistance in tick populations of cattle: A systematic review and meta-analysis. Heliyon 2022, 8, e08718. [Google Scholar] [CrossRef]
- Donnelly, M.J.; Isaacs, A.T.; Weetman, D. Identification, Validation, and Application of Molecular Diagnostics for Insecticide Resistance in Malaria Vectors. Trends Parasitol. 2016, 32, 197–206. [Google Scholar] [CrossRef]
- Feyereisen, R.; Dermauw, W.; Van Leeuwen, T. Genotype to phenotype, the molecular and physiological dimensions of resistance in arthropods. Pestic. Biochem. Physiol. 2015, 121, 61–77. [Google Scholar] [CrossRef]
- Vudriko, P.; Umemiya-Shirafuji, R.; Okwee-Acai, J.; Tayebwa, D.S.; Byaruhanga, J.; Jirapattharasate, C.; Liu, M.; Moumouni, P.F.A.; Fujisaki, K.; Xuan, X.; et al. Genetic mutations in sodium channel domain II and carboxylesterase genes associated with phenotypic resistance against synthetic pyrethroids by Rhipicephalus (Boophilus) decoloratus ticks in Uganda. Pestic. Biochem. Physiol. 2017, 143, 181–190. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. 2011. Available online: http://www.megasoftware.net (accessed on 16 October 2022).
- de La Canal, L.H.; Dall’Agnol, B.; Webster, A.; Reck, J.; Martins, J.R.; Klafke, G.M. Mechanisms of amitraz resistance in a Rhipicephalus microplus strain from southern Brazil. Ticks Tick Borne Dis. 2021, 12, 101764. [Google Scholar] [CrossRef]
- Jyoti Singh, N.K.; Singh, H.; Singh, N.K.; Rath, S.S. Genotyping amitraz resistance profiles in Rhipicephalus microplus Canestrini (Acari: Ixodidae) ticks from Punjab, India. Ticks Tick Borne Dis. 2021, 12, 101578. [Google Scholar]
- Corley, S.W.; Jonsson, N.N.; Piper, E.K.; Cutullé, C.; Stear, M.J.; Seddon, J.M. Mutation in the RmßAOR gene is associated with amitraz resistance in the cattle tick Rhipicephalus microplus. Proc. Natl. Acad. Sci. USA 2013, 110, 16772–16777. [Google Scholar] [CrossRef]
- Baron, S.; Barrero, R.A.; Black, M.; Bellgard, M.I.; van Dalen, E.; Maritz-Olivier, C. Differentially expressed genes in response to amitraz treatment suggests a proposed model of resistance to amitraz in R. decoloratus ticks. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 361–371. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region (Number of Districts) | District (Number of Farms) | Farm ID/Tick Population | Sub County | Acaricide Application Interval/Week | History of Acaricides Used in the Last 2 Years | Resistance to Amitraz Based on LPT Assay | Number of Oct/Tyr Clones Sequenced for Each Tick Population |
|---|---|---|---|---|---|---|---|
| Western (13) | Bushenyi (03) | 1BUS | Kyeizooba | Twice | AM + OP + SP | Yes (68.1%) | 03 |
| 2BUS | Kyeizooba | - | AM + SP | No | 03 | ||
| 3BUS | Kyeizooba | Twice | AM + SP + COF | No | 04 | ||
| Kiruhura (05) | 2 KRH | Kikatsi | Once | AM | No | 04 | |
| 10 KRH | Kitkatsi | Once | AM + SP + OP + COF | No | 04 | ||
| KKS | Kazo | Twice | AM + COF | Yes (0%) | 01 | ||
| KMD01 | Keishunga | Once | AM + COF | Yes (8.0%) | 04 | ||
| KMG | Keishunga | Twice | AM + COF + OP | Yes (26.7%) | 02 | ||
| Gomba (01) | MS19R | Madu | Once | AM +SP | Yes (14.5%) | 02 | |
| Mbarara (01) | 4 MBR | Ndeija | Once | SP | No | 03 | |
| Mitoma (02) | 2MTM | Mitooma | Once | AM | Yes (45.3%) | 04 | |
| 3MTM | Kashenshero | Once | OP + SP + COF | Yes (45%) | 03 | ||
| Sheema (01) | 1SHM | Kagango | Once | AM + COF | No | 01 | |
| Central (02) | Sembabule (01) | 2 SBL | Lugusulu | - | AM + OP + COF | No | 01 |
| Wakiso (01) | WKB | Wakiso | Twice | AM + COF | No | 03 | |
| East (01) | Serere (01) | TSR Þ | Serere | Once | SP | No | 04 |
| North (01) | Adjumani (01) | AM9 Þ | Pachara | Occasional | AM | No | 01 |
| Total | Farms (17) | 47 |
| Category | SNP Loci in ORF | SNP in ORF | SNP Loci Based on Previous Nomenclature by Chen et al. [26] and Baron et al. [12] | Amino Acid Substitution | Phenotype Status |
|---|---|---|---|---|---|
| Low amitraz pressure area | Locus 43 | A43G | A178G | I15V | Susceptible |
| Locus 58 | A58G | A193G | T20A | Susceptible | |
| Locus 77 | C77T | C212T | T26M | Susceptible | |
| High amitraz pressure area | Locus 1 | A1G | A135G | M1V | Resistant |
| Locus 46 | C46T | C181T | L16F | Resistant | |
| Locus 80 | T80C | T215C | V27A | Susceptible | |
| Locus 95 | T95C | T230C | V32A | Susceptible | |
| Locus 115 | A115C | A115C | M39L | Susceptible | |
| Locus 122 | A122G | A257G | D41G | Resistant | |
| Locus 199 | A199G | A334G | I67V | Susceptible | |
| Locus 215 | T215C | T350C | V72A | Resistant |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vudriko, P.; Umemiya-Shirafuji, R.; Tayebwa, D.S.; Byaruhanga, J.; Byamukama, B.; Tumwebaze, M.; Xuan, X.; Suzuki, H. Molecular Characterization of Octopamine/Tyramine Receptor Gene of Amitraz-Resistant Rhipicephalus (Boophilus) decoloratus Ticks from Uganda. Microorganisms 2022, 10, 2384. https://doi.org/10.3390/microorganisms10122384
Vudriko P, Umemiya-Shirafuji R, Tayebwa DS, Byaruhanga J, Byamukama B, Tumwebaze M, Xuan X, Suzuki H. Molecular Characterization of Octopamine/Tyramine Receptor Gene of Amitraz-Resistant Rhipicephalus (Boophilus) decoloratus Ticks from Uganda. Microorganisms. 2022; 10(12):2384. https://doi.org/10.3390/microorganisms10122384
Chicago/Turabian StyleVudriko, Patrick, Rika Umemiya-Shirafuji, Dickson Stuart Tayebwa, Joseph Byaruhanga, Benedicto Byamukama, Maria Tumwebaze, Xuenan Xuan, and Hiroshi Suzuki. 2022. "Molecular Characterization of Octopamine/Tyramine Receptor Gene of Amitraz-Resistant Rhipicephalus (Boophilus) decoloratus Ticks from Uganda" Microorganisms 10, no. 12: 2384. https://doi.org/10.3390/microorganisms10122384
APA StyleVudriko, P., Umemiya-Shirafuji, R., Tayebwa, D. S., Byaruhanga, J., Byamukama, B., Tumwebaze, M., Xuan, X., & Suzuki, H. (2022). Molecular Characterization of Octopamine/Tyramine Receptor Gene of Amitraz-Resistant Rhipicephalus (Boophilus) decoloratus Ticks from Uganda. Microorganisms, 10(12), 2384. https://doi.org/10.3390/microorganisms10122384