
Accelerated Bioconversion of Chemically Solubilized Lignite Solution to Methane by Methanogenic Consortium: Experimental Results and Their Application to the Subsurface Cultivation and Gasification Method

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Preparation of Chemically Solubilized Lignite Solution
2.2. Construction of Methanogenic Enrichment Cultures
2.3. Microcosm Experiments
2.4. Analytical Procedures
2.5. DNA Extraction and Next-Generation Sequencing
2.6. Real-Time Quantitative PCR
2.7. Statistical Analysis
2.8. Sequence Accession Numbers
3. Results
3.1. Methane Production in Microcosms Supplemented with Chemically Solubilized Lignite Solution Using the Microbial Consortium, mMEC
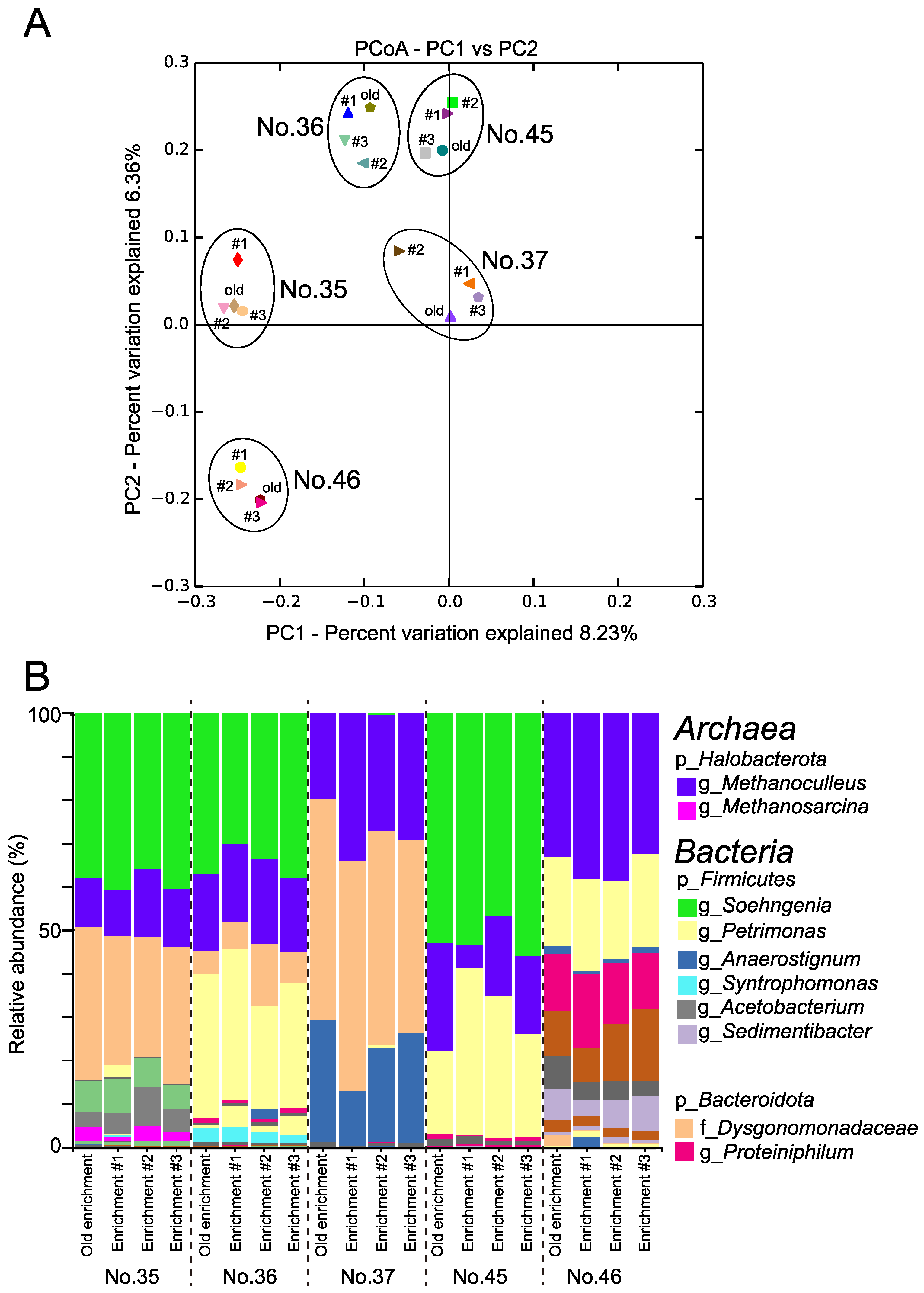
3.1.1. Characteristics of the MECs
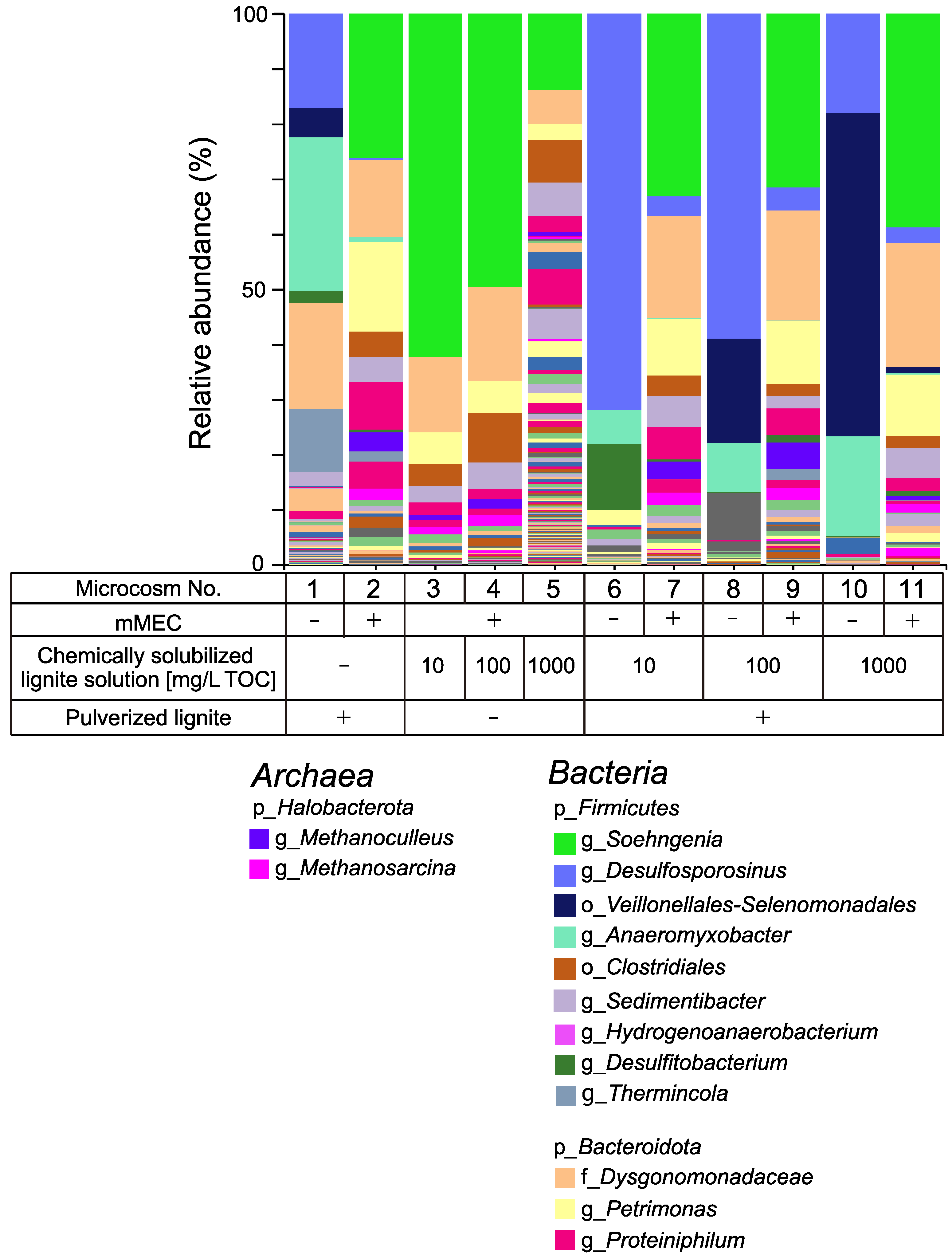
3.1.2. Methane Production Using the mMEC
3.1.3. Microbial Composition in Microcosms
3.2. Methane Production in Microcosms Supplemented with Chemically Solubilized Lignite Solution Using the Microbial Consortium, SAL25-2
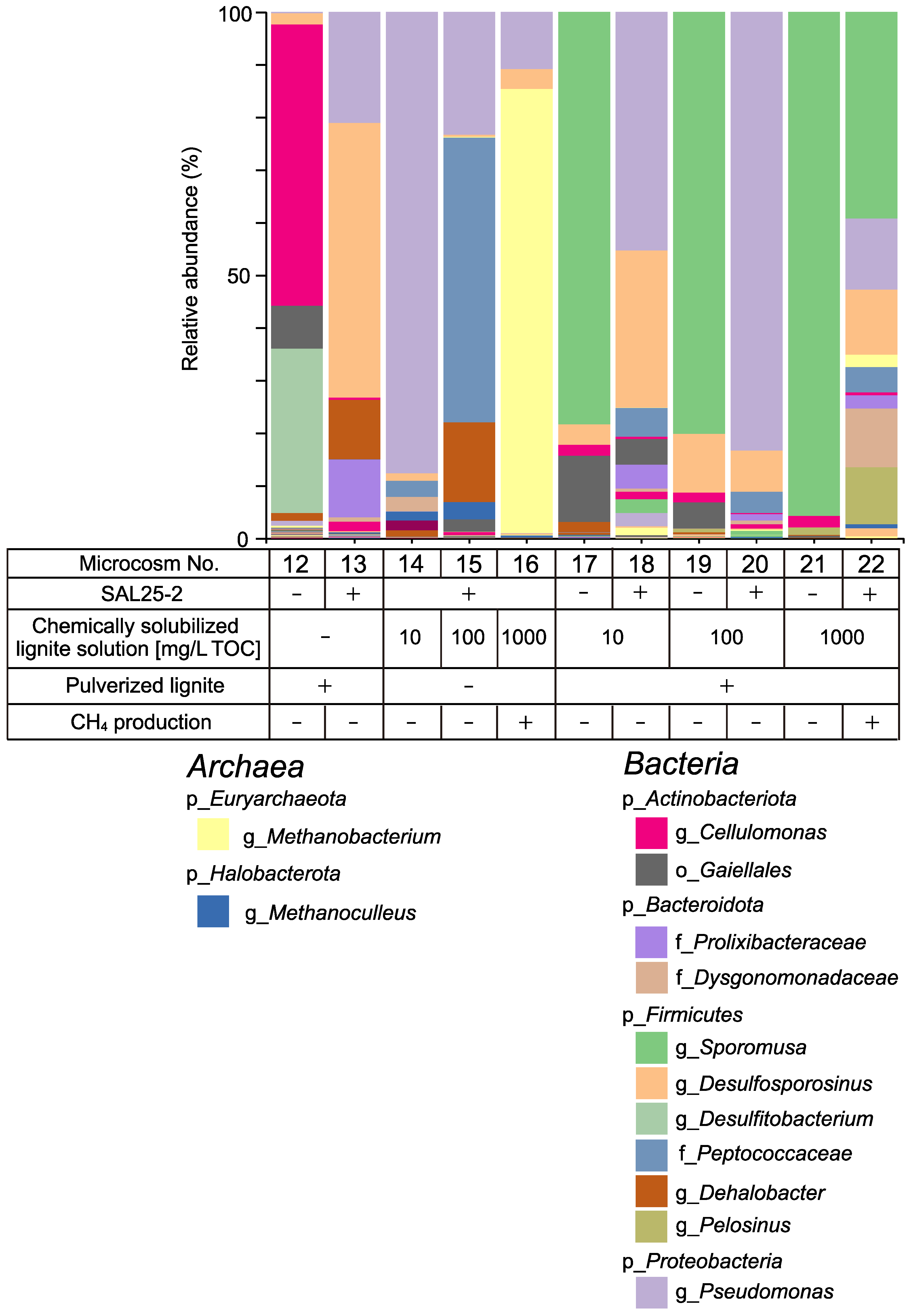
3.2.1. Methane Production Using the SAL25-2
3.2.2. Microbial Composition in Microcosms
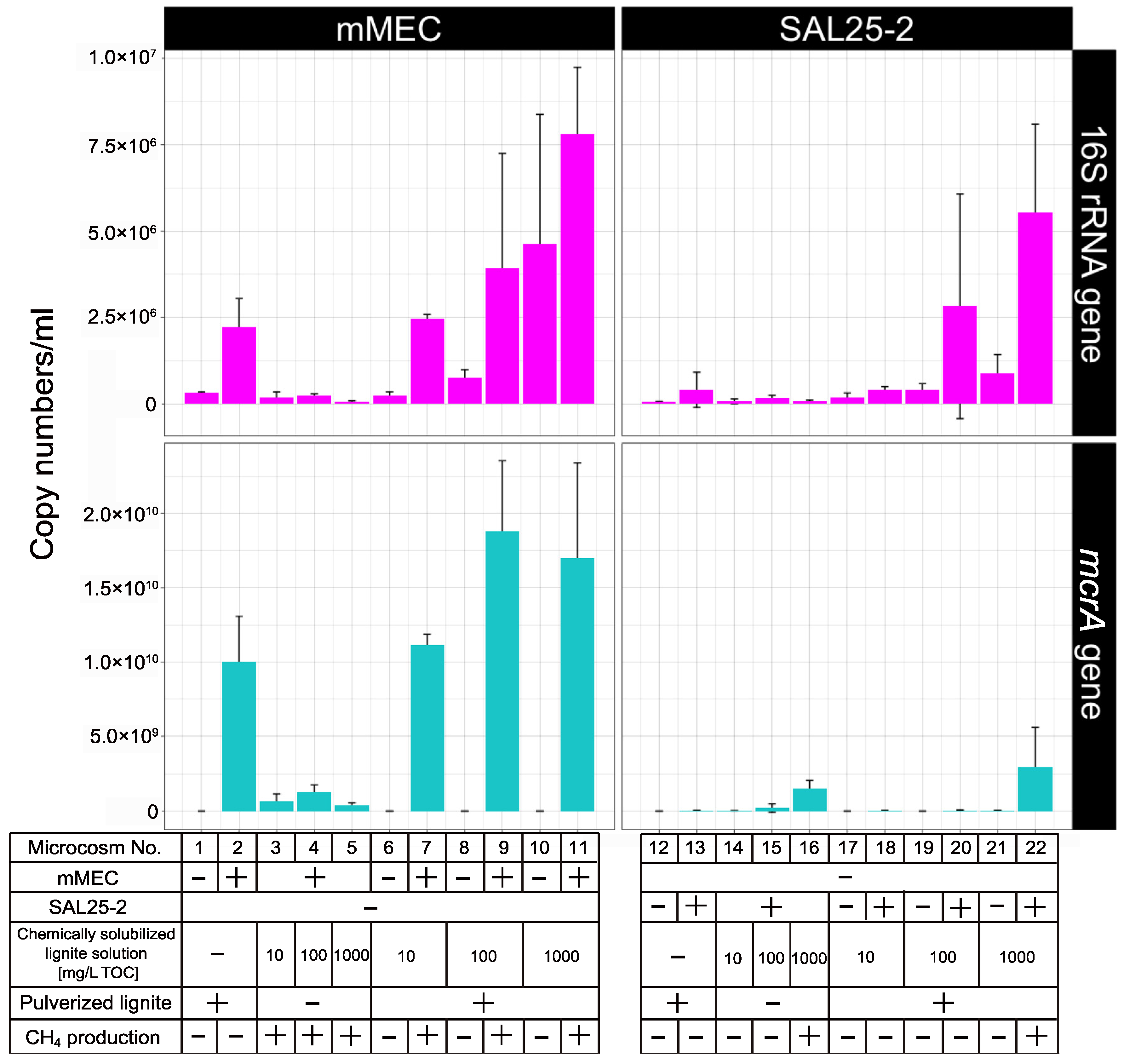
3.3. Real-Time Quantitative PCR Analysis of Microcosms
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ritchie, H.; Roser, M. Energy. Available online: https://ourworldindata.org/energy (accessed on 14 March 2022).
- Faison, B.D. Biological coal conversions. Crit. Rev. Biotechnol. 1991, 11, 347–366. [Google Scholar] [CrossRef]
- Faison, B.D.; Crawford, D.L. The Chemistry of Low Rank Coal and Its Relationship to the Biochemical Mechanisms of Coal Biotransformation; CRC Press: Boca Raton, FL, USA, 1993. [Google Scholar]
- Doskočil, L.; Grasset, L.; Enev, V.; Kalina, L.; Pekař, M. Study of water-extractable fractions from South Moravian lignite. Environ. Earth Sci. 2015, 73, 3873–3885. [Google Scholar] [CrossRef]
- Doskočil, L.; Enev, V.; Grasset, L.; Wasserbauer, J. The characterization of South Moravian lignite in its natural and treated forms using thermal degradation methods. J. Anal. Appl. Pyrol. 2017, 128, 83–91. [Google Scholar] [CrossRef]
- Fakoussa, R.M.; Hofrichter, M. Biotechnology and microbiology of coal degradation. Appl. Microbiol. Biotechnol. 1999, 52, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Hatcher, P.G.; Breger, I.A.; Szeverenyi, N.; Maciel, G.E. Nuclear magnetic resonance studies of ancient buried wood—II. Observations on the origin of coal from lignite to bituminous coal. Org. Geochem. 1982, 4, 9–18. [Google Scholar] [CrossRef]
- Park, S.Y.; Liang, Y. Biogenic methane production from coal: A review on recent research and development on microbially enhanced coalbed methane (MECBM). Fuel 2016, 166, 258–267. [Google Scholar] [CrossRef] [Green Version]
- Bumpus, J.A.; Senko, J.; Lynd, G.; Morgan, R.; Sturm, K.; Stimpson, J.; Roe, S. Biomimetic solubilization of a low rank coal: Implications for its use in methane production. Energy Fuels 1998, 12, 664–671. [Google Scholar] [CrossRef]
- Ritter, D.; Vinson, D.; Barnhart, E.; Akob, D.M.; Fields, M.W.; Cunningham, A.B.; Orem, W.; McIntosh, J.C. Enhanced microbial coalbed methane generation: A review of research, commercial activity, and remaining challenges. Int. J. Coal Geol. 2015, 146, 28–41. [Google Scholar] [CrossRef] [Green Version]
- Strąpoć, D.; Mastalerz, M.; Dawson, K.; Macalady, J.; Callaghan, A.V.; Wawrik, B.; Turich, C.; Ashby, M. Biogeochemistry of microbial coal-bed methane. Annu. Rev. Earth Planet. Sci. 2011, 39, 617–656. [Google Scholar] [CrossRef]
- Wawrik, B.; Mendivelso, M.; Parisi, V.A.; Suflita, J.M.; Davidova, I.A.; Marks, C.R.; Van Nostrand, J.D.; Liang, Y.; Zhou, J.; Huizinga, B.J.; et al. Field and laboratory studies on the bioconversion of coal to methane in the San Juan Basin. FEMS Microbiol. Ecol. 2012, 81, 26–42. [Google Scholar] [CrossRef]
- Green, M.S.; Flanegan, K.C.; Gilcrease, P.C. Characterization of a methanogenic consortium enriched from a coalbed methane well in the Powder River Basin, U.S.A. Int. J. Coal Geol. 2008, 76, 34–45. [Google Scholar] [CrossRef]
- Harris, S.H.; Smith, R.L.; Barker, C.E. Microbial and chemical factors influencing methane production in laboratory incubations of low-rank subsurface coals. Int. J. Coal Geol. 2008, 76, 46–51. [Google Scholar] [CrossRef]
- Ulrich, G.; Bower, S. Active methanogenesis and acetate utilization in Powder River Basin coals, United States. Int. J. Coal Geol. 2008, 76, 25–33. [Google Scholar] [CrossRef]
- Strąpoć, D.; Picardal, F.W.; Turich, C.; Schaperdoth, I.; Macalady, J.L.; Lipp, J.S.; Lin, Y.S.; Ertefai, T.F.; Schubotz, F.; Hinrichs, K.U.; et al. Methane-producing microbial community in a coal bed of the Illinois basin. Appl. Environ. Microbiol. 2008, 74, 2424–2432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Liang, Y.; Pandey, R.; Harpalani, S. Characterizing microbial communities dedicated for conversion of coal to methane in situ and ex situ. Int. J. Coal Geol. 2015, 146, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Papendick, S.L.; Downs, K.R.; Vo, K.D.; Hamilton, S.K.; Dawson, G.K.W.; Golding, S.D.; Gilcrease, P.C. Biogenic methane potential for Surat Basin, Queensland coal seams. Int. J. Coal Geol. 2011, 88, 123–134. [Google Scholar] [CrossRef]
- Krüger, M.; Beckmann, S.; Engelen, B.; Thielemann, T.; Cramer, B.; Schippers, A.; Cypionka, H. Microbial methane formation from hard coal and timber in an abandoned coal mine. Geomicrobiol. J. 2008, 25, 315–321. [Google Scholar] [CrossRef]
- Tang, Y.-Q.; Ji, P.; Lai, G.-L.; Chi, C.-Q.; Liu, Z.-S.; Wu, X.-L. Diverse microbial community from the coalbeds of the Ordos Basin, China. Int. J. Coal Geol. 2012, 90–91, 21–33. [Google Scholar] [CrossRef]
- Wei, M.; Yu, Z.; Jiang, Z.; Zhang, H. Microbial diversity and biogenic methane potential of a thermogenic-gas coal mine. Int. J. Coal Geol. 2014, 134–135, 96–107. [Google Scholar] [CrossRef]
- Guo, H.; Yu, Z.; Thompson, I.P.; Zhang, H. A contribution of hydrogenotrophic methanogenesis to the biogenic coal bed methane reserves of Southern Qinshui Basin, China. Appl. Microbiol. Biotechnol. 2014, 98, 9083–9093. [Google Scholar] [CrossRef]
- Shimizu, S.; Akiyama, M.; Naganuma, T.; Fujioka, M.; Nako, M.; Ishijima, Y. Molecular characterization of microbial communities in deep coal seam groundwater of northern Japan. Geobiology 2007, 5, 423–433. [Google Scholar] [CrossRef]
- Bates, B.L.; McIntosh, J.C.; Lohse, K.A.; Brooks, P.D. Influence of groundwater flowpaths, residence times and nutrients on the extent of microbial methanogenesis in coal beds: Powder River Basin, USA. Chem. Geol. 2011, 284, 45–61. [Google Scholar] [CrossRef]
- Strąpoć, D.; Mastalerz, M.; Eble, C.; Schimmelmann, A. Characterization of the origin of coalbed gases in southeastern Illinois Basin by compound-specific carbon and hydrogen stable isotope ratios. Org. Geochem. 2007, 38, 267–287. [Google Scholar] [CrossRef]
- Hamilton, S.K.; Golding, S.D.; Baublys, K.A.; Esterle, J.S. Conceptual exploration targeting for microbially enhanced coal bed methane (MECoM) in the Walloon Subgroup, Eastern Surat Basin, Australia. Int. J. Coal Geol. 2015, 138, 68–82. [Google Scholar] [CrossRef] [Green Version]
- Orem, W.H.; Voytek, M.A.; Jones, E.J.; Lerch, H.E.; Bates, A.L.; Corum, M.D.; Warwick, P.D.; Clark, A.C. Organic intermediates in the anaerobic biodegradation of coal to methane under laboratory conditions. Org. Geochem. 2010, 41, 997–1000. [Google Scholar] [CrossRef]
- Jones, E.J.; Voytek, M.A.; Corum, M.D.; Orem, W.H. Stimulation of methane generation from nonproductive coal by addition of nutrients or a microbial consortium. Appl. Environ. Microbiol. 2010, 76, 7013–7022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skulachev, V.; Bogachev, A.V.; Kasparinsky, F.O. Principles of Bioenergetics; Springer Science and Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Boone, D.R.; Whitman, W.B.; Rouvière, P. Diversity and Taxonomy of Methanogens. In Methanogenesis: Ecology, Physiology, Biochemistry & Genetics; Ferry, J.G., Ed.; Springer: Boston, MA, USA, 1993; pp. 35–80. [Google Scholar]
- Garcia, J.L.; Patel, B.K.C.; Ollivier, B. Taxonomic, phylogenetic, and ecological diversity of methanogenic Archaea. Anaerobe 2000, 6, 205–226. [Google Scholar] [CrossRef] [PubMed]
- Ferry, J.G.; Kastead, K.A. Methanogenesis. In Archaea: Molecular Cell Biology; Cavicchioli, R., Ed.; American Society of Microbiology Press: Washington, DC, USA, 2007; pp. 288–314. [Google Scholar]
- Thauer, R.K. Biochemistry of methanogenesis: A tribute to Marjory Stephenson: 1998 Marjory Stephenson Prize Lecture. Microbiology 1998, 144, 2377–2406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grahame, D.A.; Gencic, S. Methane Biochemistry. In Encyclopedia of Microbiology, 2nd ed.; Lederberg, J., Ed.; Academic Press: New York, NY, USA, 2000; Volume 3, pp. 188–198. [Google Scholar]
- Colosimo, F.; Thomas, R.; Lloyd, J.R.; Taylor, K.G.; Boothman, C.; Smith, A.D.; Lord, R.; Kalin, R.M. Biogenic methane in shale gas and coal bed methane: A review of current knowledge and gaps. Int. J. Coal Geol. 2016, 165, 106–120. [Google Scholar] [CrossRef] [Green Version]
- Mayumi, D.; Mochimaru, H.; Tamaki, H.; Yamamoto, K.; Yoshioka, H.; Suzuki, Y.; Kamagata, Y.; Sakata, S. Methane production from coal by a single methanogen. Science 2016, 354, 222–225. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Mae, K.; Okutsu, H.; Mizutani, N.-a. New oxidative degradation method for producing fatty acids in high yields and high selectivity from low-rank coals. Energy Fuels 1996, 10, 1196–1201. [Google Scholar] [CrossRef]
- Mae, K.; Shindo, H.; Miura, K. A new two-step oxidative degradation method for producing valuable chemicals from low rank coals under mild conditions. Energy Fuels 2001, 15, 611–617. [Google Scholar] [CrossRef]
- Huang, Z.; Urynowicz, M.A.; Haider, R.; Sattar, H.; Saleem, M.; Hoang, L.; Han, N.T.; To, K.A.; Hung, L.Q.; Ali, M.I.; et al. Biogenic methane generation from Vietnamese coal after pretreatment with hydrogen peroxide. Int. J. Energy Res. 2021, 45, 18713–18721. [Google Scholar] [CrossRef]
- Liu, F.; Guo, H.; Wang, Q.; Haider, R.; Urynowicz, M.A.; Fallgren, P.H.; Jin, S.; Tang, M.; Chen, B.; Huang, Z. Characterization of organic compounds from hydrogen peroxide-treated subbituminous coal and their composition changes during microbial methanogenesis. Fuel 2019, 237, 1209–1216. [Google Scholar] [CrossRef]
- Wang, Q.; Guo, H.; Wang, H.; Urynowicz, M.A.; Hu, A.; Yu, C.-P.; Fallgren, P.; Jin, S.; Zheng, H.; Zeng, R.J.; et al. Enhanced production of secondary biogenic coalbed natural gas from a subbituminous coal treated by hydrogen peroxide and its geochemical and microbiological analyses. Fuel 2019, 236, 1345–1355. [Google Scholar] [CrossRef]
- Chen, T.; Rodrigues, S.; Golding, S.D.; Rudolph, V. Improving coal bioavailability for biogenic methane production via hydrogen peroxide oxidation. Int. J. Coal Geol. 2018, 195, 402–414. [Google Scholar] [CrossRef]
- Huang, Z.; Urynowicz, M.A.; Colberg, P.J.S. Bioassay of chemically treated subbituminous coal derivatives using Pseudomonas putida F1. Int. J. Coal Geol. 2013, 115, 97–105. [Google Scholar] [CrossRef]
- Huang, Z.; Urynowicz, M.A.; Colberg, P.J.S. Stimulation of biogenic methane generation in coal samples following chemical treatment with potassium permanganate. Fuel 2013, 111, 813–819. [Google Scholar] [CrossRef]
- Tamamura, S.; Murakami, T.; Aramaki, N.; Ueno, A.; Badrul, A.A.K.M.; Haq, S.R.; Igarashi, T.; Kaneko, K. Reaction of lignite with dilute hydrogen peroxide to produce substrates for methanogens at in situ subsurface temperatures. Int. J. Coal Geol. 2016, 167, 230–237. [Google Scholar] [CrossRef]
- Aramaki, N.; Tamamura, S.; Ueno, A.; Badrul, A.A.K.M.; Murakami, T.; Tamazawa, S.; Yamaguchi, S.; Aoyama, H.; Kaneko, K. Experimental investigation on the feasibility of industrial methane production in the subsurface environment via microbial activities in northern Hokkaido, Japan—A process involving subsurface cultivation and gasification. Energy Convers. Manag. 2017, 153, 566–575. [Google Scholar] [CrossRef]
- Kaneko, K.; Shimizu, S.; Tamamura, S.; Ueno, A.; Aramaki, N.; Ishijima, Y.; Oomi, Y.; Endo, T. A Method for Producing Methane Gas in a Formation from Coal and/or Diatomite Contained in the Formation. Japan Patent JP6396068, 26 September 2018. [Google Scholar]
- Kato, M.; Katsui, Y.; Kitagawa, Y.; Matsui, M. Regional Geology of Japan Part 1. In Hokkaido; Kyoritsu Shuppan: Tokyo, Japan, 1990; pp. 186–190. (In Japanese) [Google Scholar]
- Aramaki, N.; Tamamura, S.; Alam, A.K.M.B.; Yamamoto, S.; Shigeyoshi, H.; Kaneko, K. Organic matter decomposition of lignite and coaly shale in soya coal-bearing formations using hydrogen peroxide for subsurface cultivation and gasification. J. Min. Mater. Proc Inst. Jpn. 2016, 132, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Haq, S.R.; Tamamura, S.; Ueno, A.; Tamazawa, S.; Aramaki, N.; Murakami, T.; Alam, A.K.M.B.; Igarashi, T.; Kaneko, K. Biogenic methane generation using solutions from column reactions of lignite with hydrogen peroxide. Int. J. Coal Geol. 2018, 197, 66–73. [Google Scholar] [CrossRef]
- Shimizu, S.; Akiyama, M.; Ishijima, Y.; Hama, K.; Kunimaru, T.; Naganuma, T. Molecular characterization of microbial communities in fault-bordered aquifers in the Miocene formation of northernmost Japan. Geobiology 2006, 4, 203–213. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, S.; Ueno, A.; Ishijima, Y. Microbial mmunities associated with acetate-rich gas-petroleum reservoir surface facilities. Biosci. Biotechnol. Biochem. 2011, 75, 1835–1837. [Google Scholar] [CrossRef] [Green Version]
- Ueno, A.; Tamazawa, S.; Tamamura, S.; Aramaki, N.; Alam Badrul, A.K.M.; Murakami, T.; Yamaguchi, S.; Yamagishi, J.; Tamaki, H.; Mayumi, D.; et al. Improvement of terrestrial groundwater sampling method affects microbial community analysis. Geomicrobiol. J. 2019, 36, 303–316. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl Acad. Sci. USA 2011, 108 (Suppl. 1), 4516–4522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estaki, M.; Jiang, L.; Bokulich, N.A.; McDonald, D.; González, A.; Kosciolek, T.; Martino, C.; Zhu, Q.; Birmingham, A.; Vázquez-Baeza, Y.; et al. QIIME 2 enables comprehensive end-to-end analysis of diverse microbiome data and comparative studies with publicly available data. Curr. Protoc. Bioinform. 2020, 70, e100. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Holmes, S.P. Exact sequence variants should replace operational taxonomic units in marker-gene data analysis. ISME J. 2017, 11, 2639–2643. [Google Scholar] [CrossRef]
- Juottonen, H.; Galand, P.E.; Yrjälä, K. Detection of methanogenic archaea in peat: Comparison of PCR primers targeting the mcrA gene. Res. Microbiol. 2006, 157, 914–921. [Google Scholar] [CrossRef] [PubMed]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef] [Green Version]
- Haq, S.R.; Tamamura, S.; Igarashi, T.; Kaneko, K. Characterization of organic substances in lignite before and after hydrogen peroxide treatment: Implications for microbially enhanced coalbed methane. Int. J. Coal Geol. 2018, 185, 1–11. [Google Scholar] [CrossRef]
- Jones, E.J.P.; Voytek, M.A.; Warwick, P.D.; Corum, M.D.; Cohn, A.; Bunnell, J.E.; Clark, A.C.; Orem, W.H. Bioassay for estimating the biogenic methane-generating potential of coal samples. Int. J. Coal Geol. 2008, 76, 138–150. [Google Scholar] [CrossRef]
- Doskočil, L.; Grasset, L.; Válková, D.; Pekař, M. Hydrogen peroxide oxidation of humic acids and lignite. Fuel 2014, 134, 406–413. [Google Scholar] [CrossRef]
- Liu, Z.-X.; Liu, Z.-C.; Zong, Z.-M.; Wei, X.-Y.; Wang, J.; Lee, C.W. GC/MS analysis of water-soluble products from the mild oxidation of Longkou brown coal with H2O2. Energy Fuels 2003, 17, 424–426. [Google Scholar] [CrossRef]
- Pan, X.; Angelidaki, I.; Alvarado-Morales, M.; Liu, H.; Liu, Y.; Huang, X.; Zhu, G. Methane production from formate, acetate and H2/CO2; focusing on kinetics and microbial characterization. Bioresour. Technol. 2016, 218, 796–806. [Google Scholar] [CrossRef]
- Purwantini, E.; Torto-Alalibo, T.; Lomax, J.; Setubal, J.C.; Tyler, B.M.; Mukhopadhyay, B. Genetic resources for methane production from biomass described with the Gene Ontology. Front. Microbiol. 2014, 5, 634. [Google Scholar] [CrossRef] [Green Version]
- Xiao, K.K.; Guo, C.H.; Zhou, Y.; Maspolim, Y.; Wang, J.Y.; Ng, W.J. Acetic acid inhibition on methanogens in a two-phase anaerobic process. Biochem. Eng. J. 2013, 75, 1–7. [Google Scholar] [CrossRef]
- Liu, Y.; Whitman, W.B. Metabolic, phylogenetic, and ecological diversity of the methanogenic archaea. Ann. N. Y. Acad. Sci. 2008, 1125, 171–189. [Google Scholar] [CrossRef] [PubMed]
- Martins, G.; Salvador, A.F.; Pereira, L.; Alves, M.M. Methane production and conductive materials: A critical review. Environ. Sci. Technol. 2018, 52, 10241–10253. [Google Scholar] [CrossRef] [Green Version]
- Salvador, A.F.; Martins, G.; Melle-Franco, M.; Serpa, R.; Stams, A.J.M.; Cavaleiro, A.J.; Pereira, M.A.; Alves, M.M. Carbon nanotubes accelerate methane production in pure cultures of methanogens and in a syntrophic coculture. Environ. Microbiol. 2017, 19, 2727–2739. [Google Scholar] [CrossRef] [Green Version]
- Dang, Y.; Sun, D.; Woodard, T.L.; Wang, L.Y.; Nevin, K.P.; Holmes, D.E. Stimulation of the anaerobic digestion of the dry organic fraction of municipal solid waste (OFMSW) with carbon-based conductive materials. Bioresour. Technol. 2017, 238, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Konieczna, A.; Roman, K.; Borek, K.; Grzegorzewska, E. GHG and NH3 emissions vs. energy efficiency of maize production technology: Evidence from Polish farms; a further Study. Energies 2021, 14, 5574. [Google Scholar] [CrossRef]
- Lovley, D.R. Syntrophy goes electric: Direct interspecies electron transfer. Annu. Rev. Microbiol. 2017, 71, 643–664. [Google Scholar] [CrossRef]
- Lin, R.; Cheng, J.; Zhang, J.; Zhou, J.; Cen, K.; Murphy, J.D. Boosting biomethane yield and production rate with graphene: The potential of direct interspecies electron transfer in anaerobic digestion. Bioresour. Technol. 2017, 239, 345–352. [Google Scholar] [CrossRef]
- Zhuang, L.; Ma, J.; Yu, Z.; Wang, Y.; Tang, J. Magnetite accelerates syntrophic acetate oxidation in methanogenic systems with high ammonia concentrations. Microb. Biotechnol. 2018, 11, 710–720. [Google Scholar] [CrossRef]
- Jin, Z.; Zhao, Z.; Zhang, Y. Potential of direct interspecies electron transfer in synergetic enhancement of methanogenesis and sulfate removal in an up-flow anaerobic sludge blanket reactor with magnetite. Sci. Total Environ. 2019, 677, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Summers, Z.M.; Fogarty, H.E.; Leang, C.; Franks, A.E.; Malvankar, N.S.; Lovley, D.R. Direct exchange of electrons within aggregates of an evolved syntrophic coculture of anaerobic bacteria. Science 2010, 330, 1413–1415. [Google Scholar] [CrossRef] [Green Version]
- Mostafa, A.; Im, S.; Song, Y.-C.; Ahn, Y.; Kim, D.-H. Enhanced anaerobic digestion by stimulating DIET reaction. Processes 2020, 8, 424. [Google Scholar] [CrossRef]
- Guo, H.; Dong, Z.; Liu, X.; Bai, Y.; Gao, Z.; Xia, D. Analysis of methanogens adsorption and biogas production characteristics from different coal surfaces. Environ. Sci. Pollut. Res. Int. 2019, 26, 13825–13832. [Google Scholar] [CrossRef] [PubMed]
- Hazrin-Chong, N.H.; Das, T.; Manefield, M. Surface physico-chemistry governing microbial cell attachment and biofilm formation on coal. Int. J. Coal Geol. 2021, 236, 103671. [Google Scholar] [CrossRef]
- Inagaki, F.; Hinrichs, K.-U.; Kubo, Y.; Bowles, M.W.; Heuer, V.B.; Hong, W.-L.; Hoshino, T.; Ijiri, A.; Imachi, H.; Ito, M.; et al. Exploring deep microbial life in coal-bearing sediment down to ~2.5 km below the ocean floor. Science 2015, 349, 420–424. [Google Scholar] [CrossRef] [Green Version]
- Donlan, R.M. Biofilms: Microbial life on surfaces. Emerg. Infect. Dis. 2002, 8, 881–890. [Google Scholar] [CrossRef]
- Zobell, C.E. The Effect of Solid Surfaces upon Bacterial Activity. J. Bacteriol. 1943, 46, 39–56. [Google Scholar] [CrossRef] [Green Version]
- Jetten, M.S.M.; Stams, A.J.M.; Zehnder, A.J.B. Methanogenesis from acetate: A comparison of the acetate metabolism in Methanothrix soehngenii and Methanosarcina spp. FEMS Microbiol. Lett. 1992, 88, 181–198. [Google Scholar] [CrossRef]
- Beifuss, U.; Tietze, M.; Bäumer, S.; Deppenmeier, U. Methanophenazine: Structure, total synthesis, and function of a new cofactor from methanogenic archaea. Angew. Chem. Int. Ed. Engl. 2000, 39, 2470–2472. [Google Scholar] [CrossRef]
- Fu, L.; Song, T.; Zhang, W.; Zhang, J.; Lu, Y. Stimulatory effect of magnetite nanoparticles on a highly enriched butyrate-oxidizing consortium. Front. Microbiol. 2018, 9, 1480. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Zhou, T.; Wang, J.; You, L.; Lu, Y.; Yu, L.; Zhou, S. NanoFe3O4 as solid electron shuttles to accelerate acetotrophic methanogenesis by Methanosarcina barkeri. Front. Microbiol. 2019, 10, 388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, S.; Hashimoto, K.; Watanabe, K. Methanogenesis facilitated by electric syntrophy via (semi)conductive iron-oxide minerals. Environ. Microbiol. 2012, 14, 1646–1654. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Rotaru, A.-E.; Shrestha, P.M.; Malvankar, N.S.; Nevin, K.P.; Lovley, D.R. Promoting direct interspecies electron transfer with activated carbon. Energy Environ. Sci. 2012, 5, 8982–8989. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Original Solution | Concentrations in Microcosms | ||
|---|---|---|---|---|
| pH | 2.4 | |||
| Dissolved total organic carbon [mg/L TOC] | 2800 | 1000 | 100 | 10 |
| Dissolved nitrogen [mg/L] | 145 | 52.0 | 5.2 | 0.5 |
| Acetate [mg/L] | 937 | 335 | 33.5 | 3.3 |
| Formate [mg/L] | 656 | 235 | 23.5 | 2.3 |
| Cl− [mg/L] | 5.40 | 1.9 | 0.2 | 0.02 |
| NO3− [mg/L] | 7.50 | 2.7 | 0.3 | 0.03 |
| SO42− [mg/L] | 102 | 36.4 | 3.6 | 0.4 |
| Malonate [mg/L] | 1060 | 378 | 37.8 | 3.8 |
| Succinate [mg/L] | 241 | 86.0 | 8.6 | 0.9 |
| Oxalate [mg/L] | 1010 | |||
| H2O2 [%] | <10−4 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ueno, A.; Tamazawa, S.; Tamamura, S.; Murakami, T.; Kiyama, T.; Inomata, H.; Aramaki, N.; Yoshida, K.; Yamaguchi, S.; Aoyama, H.; et al. Accelerated Bioconversion of Chemically Solubilized Lignite Solution to Methane by Methanogenic Consortium: Experimental Results and Their Application to the Subsurface Cultivation and Gasification Method. Microorganisms 2022, 10, 1984. https://doi.org/10.3390/microorganisms10101984
Ueno A, Tamazawa S, Tamamura S, Murakami T, Kiyama T, Inomata H, Aramaki N, Yoshida K, Yamaguchi S, Aoyama H, et al. Accelerated Bioconversion of Chemically Solubilized Lignite Solution to Methane by Methanogenic Consortium: Experimental Results and Their Application to the Subsurface Cultivation and Gasification Method. Microorganisms. 2022; 10(10):1984. https://doi.org/10.3390/microorganisms10101984
Chicago/Turabian StyleUeno, Akio, Satoshi Tamazawa, Shuji Tamamura, Takuma Murakami, Tamotsu Kiyama, Hidenori Inomata, Noritaka Aramaki, Kunihiko Yoshida, Shinji Yamaguchi, Hideo Aoyama, and et al. 2022. "Accelerated Bioconversion of Chemically Solubilized Lignite Solution to Methane by Methanogenic Consortium: Experimental Results and Their Application to the Subsurface Cultivation and Gasification Method" Microorganisms 10, no. 10: 1984. https://doi.org/10.3390/microorganisms10101984
APA StyleUeno, A., Tamazawa, S., Tamamura, S., Murakami, T., Kiyama, T., Inomata, H., Aramaki, N., Yoshida, K., Yamaguchi, S., Aoyama, H., Naganuma, T., & Igarashi, T. (2022). Accelerated Bioconversion of Chemically Solubilized Lignite Solution to Methane by Methanogenic Consortium: Experimental Results and Their Application to the Subsurface Cultivation and Gasification Method. Microorganisms, 10(10), 1984. https://doi.org/10.3390/microorganisms10101984