
Characterization and Biological Activity of a Novel Exopolysaccharide Produced by Pediococcus pentosaceus SSC–12 from Silage


Abstract
:1. Introduction
2. Materials and Methods
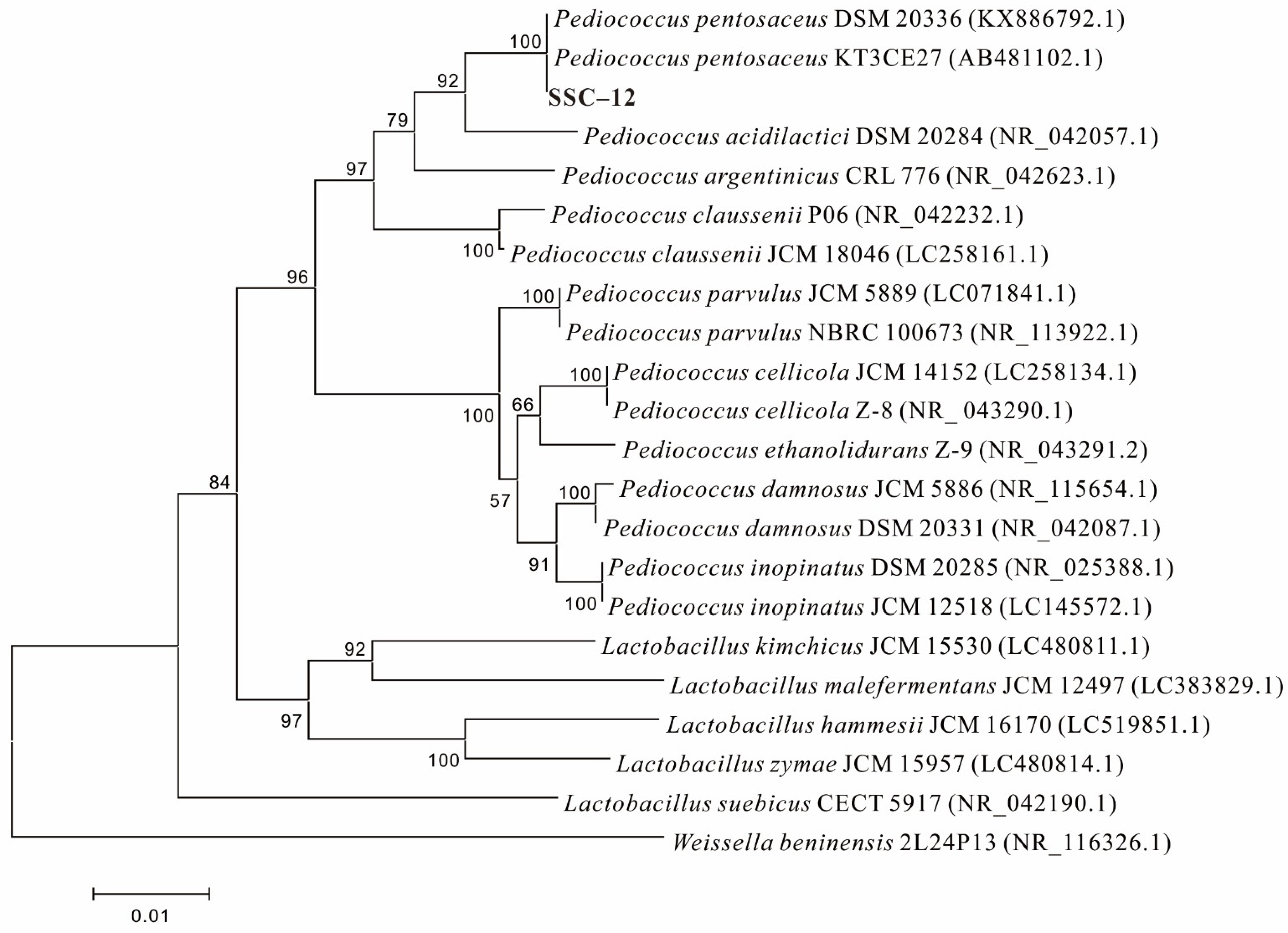
2.1. Screening and Identification of EPS-Producing LAB
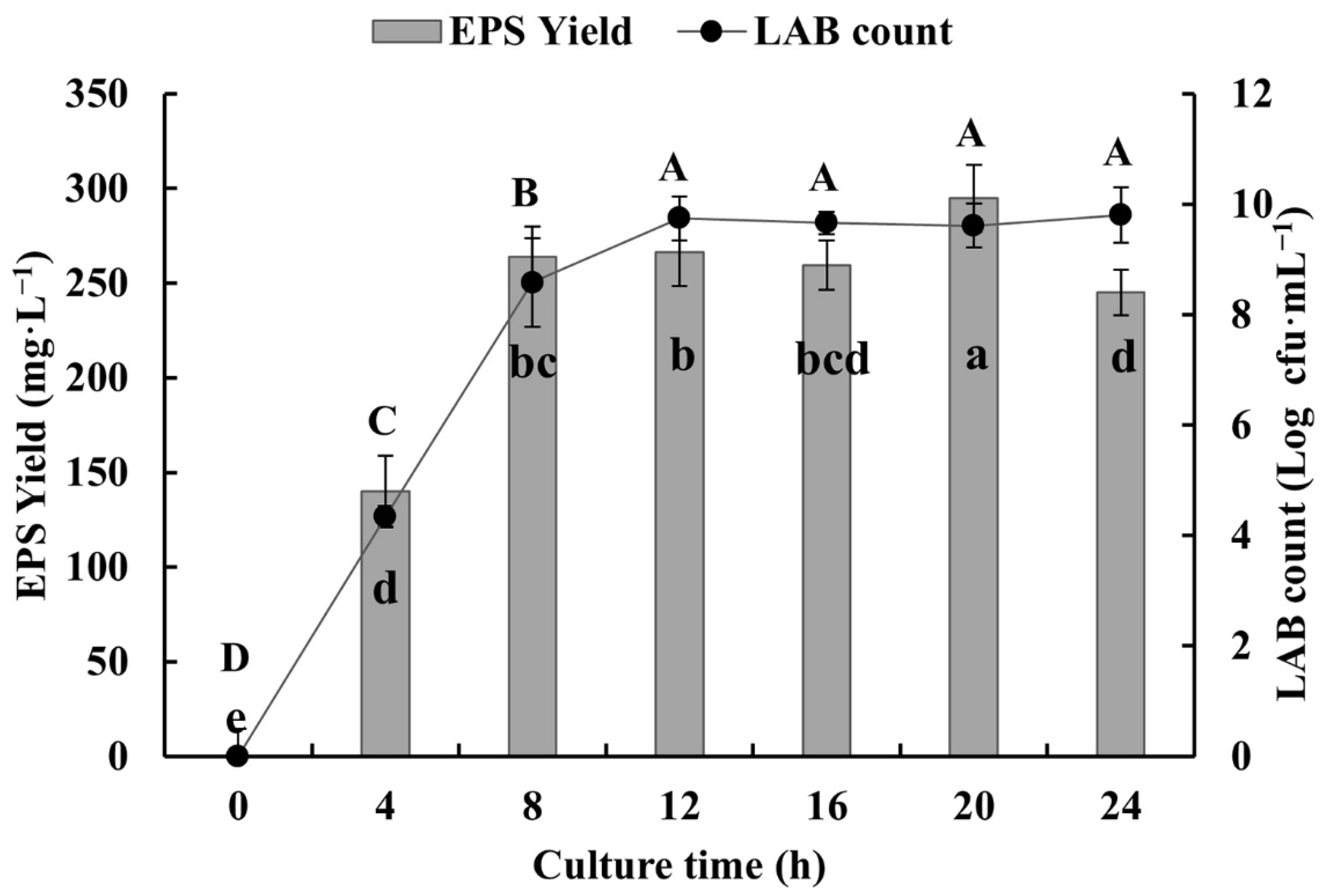
2.2. Analysis of Strain Growth and EPS Production
2.3. Molecular Weight (Mw) Determination of EPS
2.4. Determination of EPS Monosaccharide Composition
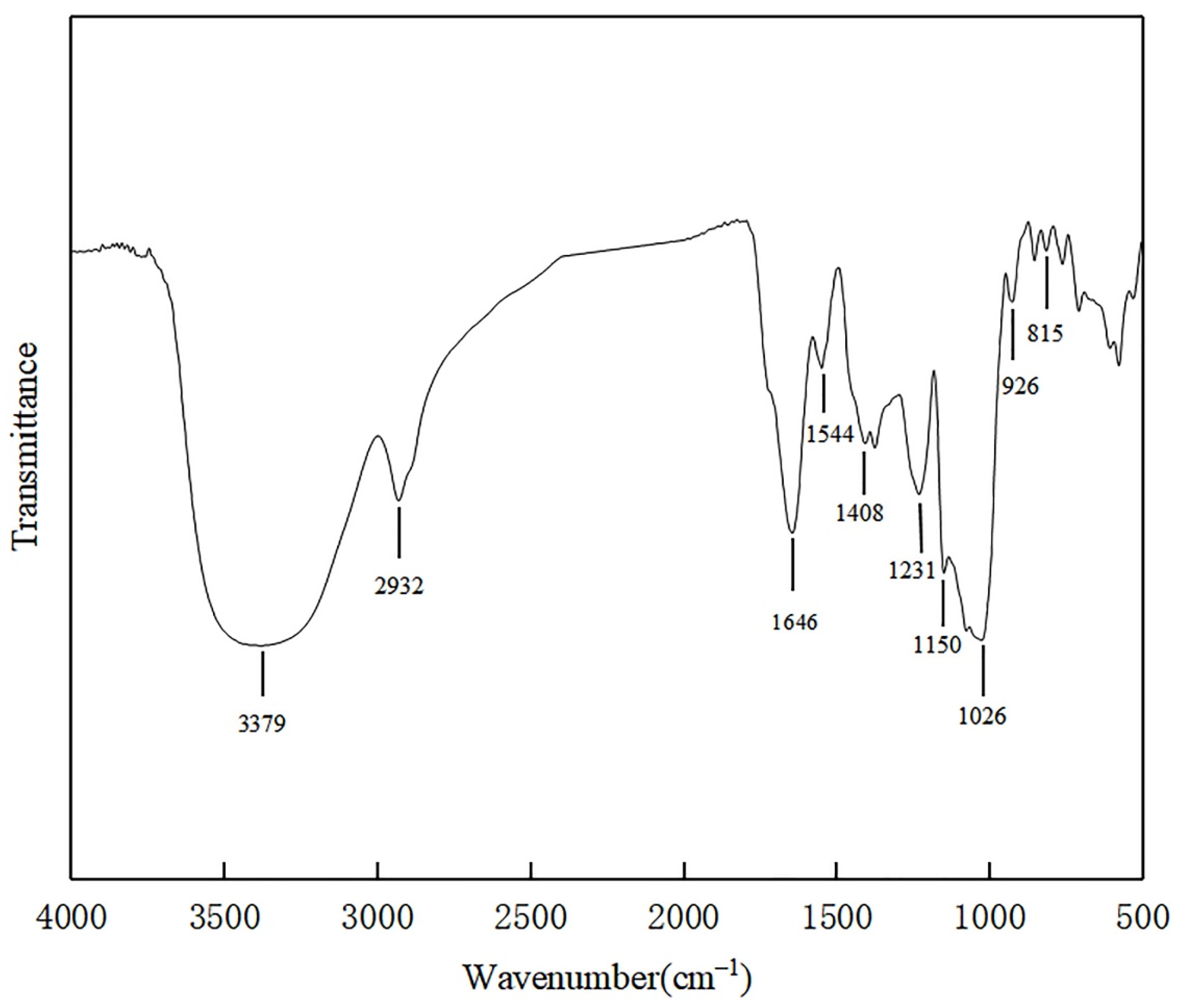
2.5. Fourier Transform-Infrared (FT-IR) Spectroscopic Analysis of EPS
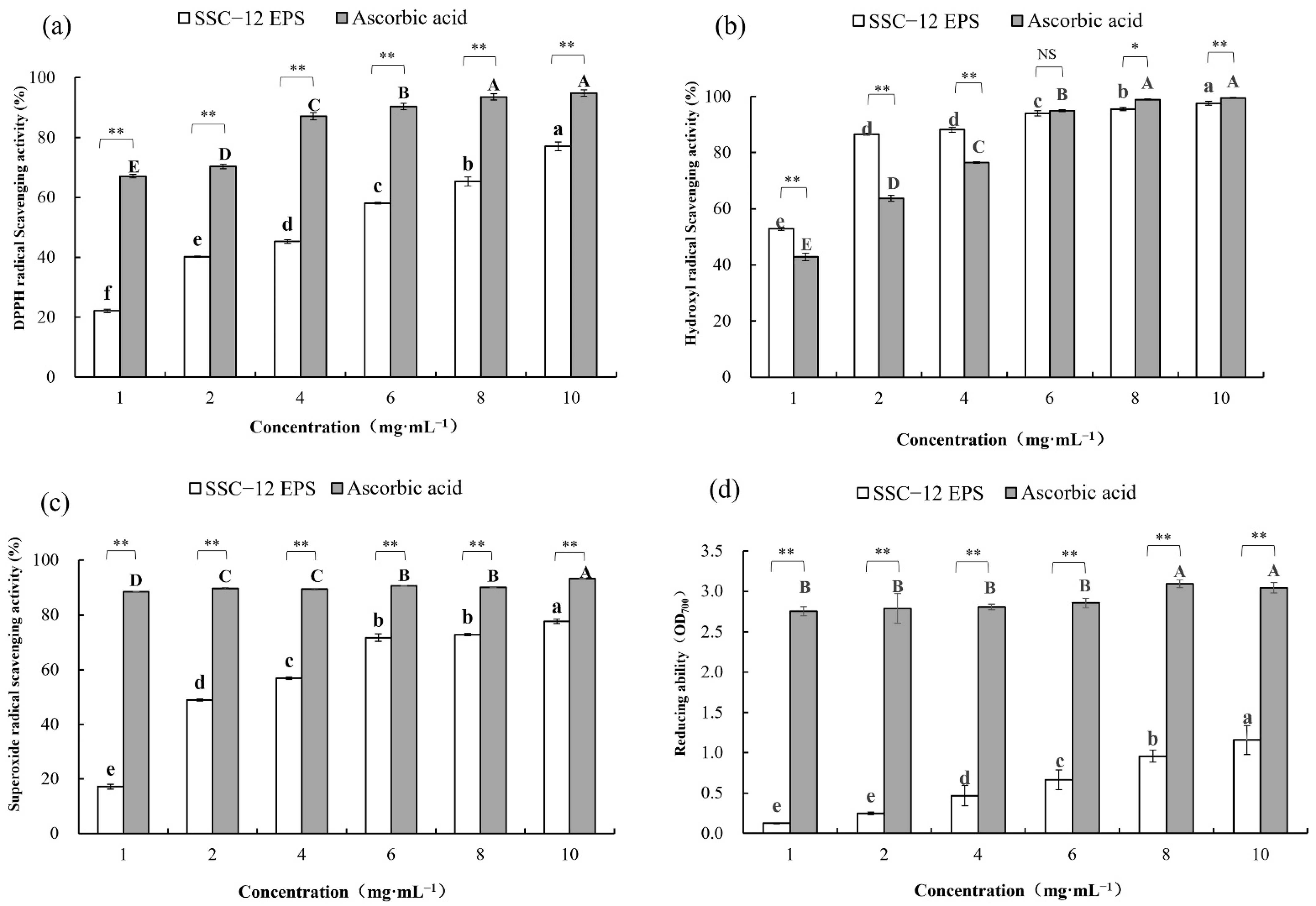
2.6. Assessment of EPS Antioxidant Activity
2.6.1. DPPH Radical Scavenging Ability
2.6.2. Hydroxyl Radical Scavenging Ability
2.6.3. Superoxide Radical Scavenging Activity
2.6.4. Reducing Ability
2.7. Measurements of EPS Antibacterial Ability
2.8. Statistical Analysis
3. Results and Discussion
3.1. EPS-Producing LAB and EPS Yield
3.2. EPS-Producing LAB and EPS Yield
3.3. Molecular Weight of EPS
3.4. Monosaccharide Composition of EPS
3.5. Fourier Transform Infrared (FT-IR) Analysis of EPS
3.6. Antioxidant Activity of EPS
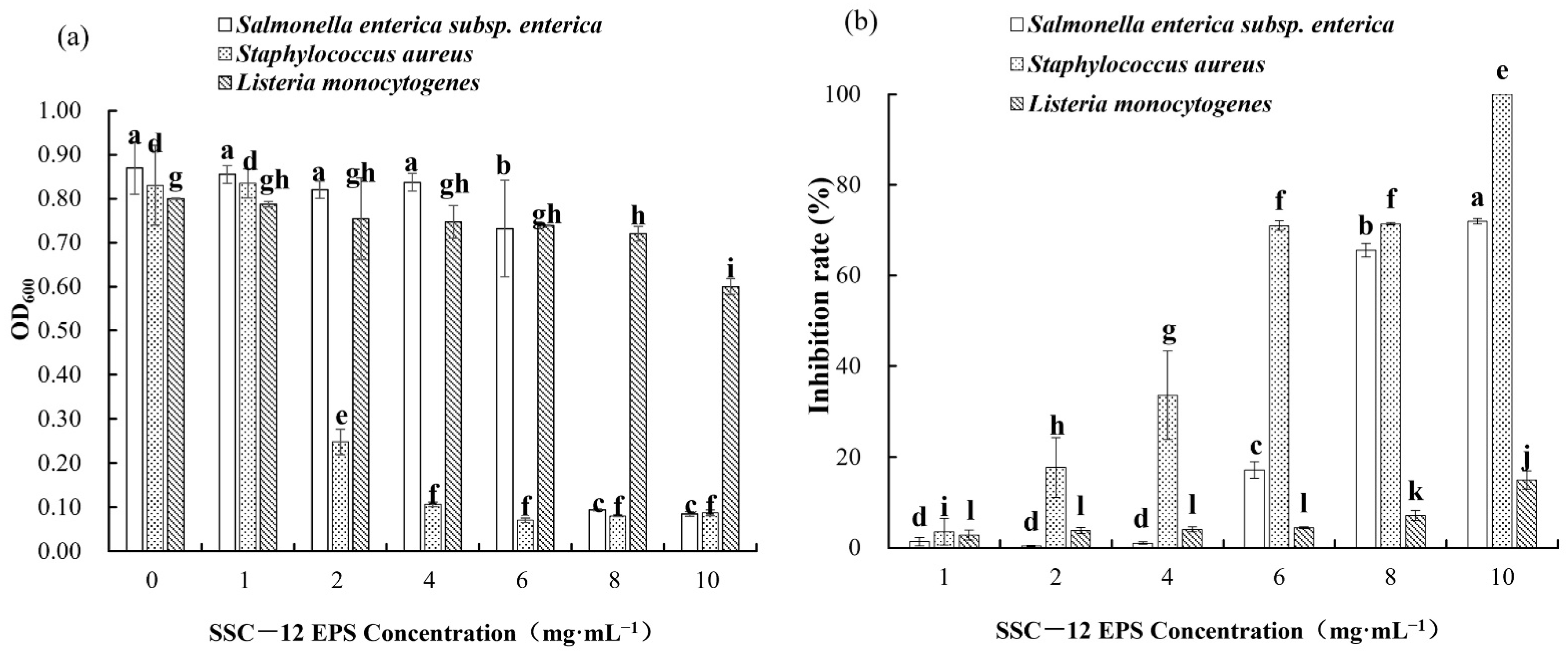
3.7. Antibacterial Ability
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, Z.Q.; Zhang, Z.H.; Qiu, L.; Zhang, F.; Xu, X.P.; Wei, H.; Tao, X.Y. Characterization and bioactivities of the exopolysaccharide from a probiotic strain of Lactobacillus plantarum WLPL04. J. Dairy Sci. 2017, 100, 6895–6905. [Google Scholar] [CrossRef]
- Adesulu-Dahunsi, A.T.; Sanni, A.I.; Jeyaram, K.; Ojediran, J.O.; Ogunsakin, A.O.; Banwo, K. Extracellular polysaccharide from Weissella confusa OF126: Production, optimization, and characterization. Int. J. Biol. Macromol. 2018, 111, 514–525. [Google Scholar] [CrossRef] [Green Version]
- Li, S.J.; Huang, R.H.; Shah, N.P.; Tao, X.Y.; Xiong, Y.H.; Wei, H. Antioxidant and antibacterial activities of exopolysaccharides from Bifidobacterium bifidum WBINO3 and Lactobacillus plantarum R315. J. Dairy Sci. 2014, 97, 7334–7343. [Google Scholar] [CrossRef] [Green Version]
- Nacher-Vazquez, M.; Ballesteros, N.; Canales, A.; Saint-Jean, S.R.; Perez-Prieto, S.I.; Prieto, A.; Aznar, R.; Lopez, P. Dextrans produced by lactic acid bacteria exhibit antiviral and immunomodulatory activity against salmonid viruses. Carbohydr. Polym. 2015, 124, 292–301. [Google Scholar] [CrossRef]
- Surayot, U.; Wang, J.G.; Seesuriyachan, P.; Kuntiya, A.; Tabarsa, M.; Lee, Y.; Kim, J.K.; Park, W.; You, S. Exopolysaccharides from lactic acid bacteria: Structural analysis, molecular weight effect on immunomodulation. Int. J. Biol. Macromol. 2014, 68, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.; Kim, D.H.; Kang, I.B.; Kim, H.; Song, K.Y.; Kim, H.S.; Seo, K.H. Characterization and antibacterial activity of a novel exopolysaccharide produced by Lactobacillus kefiranofaciens DN1 isolated from kefir. Food Control 2017, 78, 436–442. [Google Scholar] [CrossRef]
- Abd El Ghany, K.; Hamouda, R.; Abd Elhafez, E.; Mahrous, H.; Salem-Bekhit, M.; Hamza, H.A. A potential role of Lactobacillus acidophilus LA1 and its exopolysaccharides on cancer cells in male albino mice. Biotechnol. Biotechnol. Equip. 2015, 29, 977–983. [Google Scholar] [CrossRef] [Green Version]
- Silva, L.A.; Neto, J.; Cardarelli, H.R. Exopolysaccharides produced by Lactobacillus plantarum: Technological properties, biological activity, and potential application in the food industry. Ann. Microbiol. 2019, 69, 321–328. [Google Scholar] [CrossRef]
- Xu, Y.M.; Cui, Y.L.; Yue, F.F.; Liu, L.H.; Shan, Y.Y.; Liu, B.F.; Zhou, Y.; Lu, X. Exopolysaccharides produced by lactic acid bacteria and Bifidobacteria: Structures, physiochemical functions and applications in the food industry. Food Hydrocoll. 2019, 94, 475–499. [Google Scholar] [CrossRef]
- Rajoka, M.S.R.; Wu, Y.G.; Mehwish, H.M.; Bansal, M.; Zhao, L.Q. Lactobacillus exopolysaccharides: New perspectives on engineering strategies, physiochemical functions, and immunomodulatory effects on host health. Trends Food Sci. Technol. 2020, 103, 36–48. [Google Scholar] [CrossRef]
- Zhou, Y.; Cui, Y.H.; Qu, X.J. Exopolysaccharides of lactic acid bacteria: Structure, bioactivity and associations: A review. Carbohydr. Polym. 2019, 207, 317–332. [Google Scholar] [CrossRef]
- AlKalbani, N.S.; Turner, M.S.; Ayyash, M.M. Isolation, identification, and potential probiotic characterization of isolated lactic acid bacteria and in vitro investigation of the cytotoxicity, antioxidant, and antidiabetic activities in fermented sausage. Microb. Cell. Fact. 2019, 18, 188. [Google Scholar] [CrossRef] [Green Version]
- Dilna, S.V.; Surya, H.; Aswathy, R.G.; Varsha, K.K.; Sakthikumar, D.N.; Pandey, A.; Nampoothiri, K.M. Characterization of an exopolysaccharide with potential health-benefit properties from a probiotic Lactobacillus plantarum RJF(4). LWT-Food Sci. Technol. 2015, 64, 1179–1186. [Google Scholar] [CrossRef]
- Bomfim, V.B.; Neto, J.; Leite, K.S.; Vieira, E.D.; Iacomini, M.; Silva, C.M.; dos Santos, K.M.O.; Cardarelli, H.R. Partial characterization and antioxidant activity of exopolysaccharides produced by Lactobacillus plantarum CNPC003. LWT-Food Sci. Technol. 2020, 127, 109349. [Google Scholar] [CrossRef]
- Xu, X.Q.; Peng, Q.; Zhang, Y.W.; Tian, D.D.; Zhang, P.B.; Huang, Y.; Ma, L.; Qiao, Y.; Shi, B. A novel exopolysaccharide produced by Lactobacillus coryniformis NA-3 exhibits antioxidant and biofilm-inhibiting properties in vitro. Food Nutr. Res. 2020, 64, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polak-Berecka, M.; Wasko, A.; Szwajgier, D.; Choma, A. Bifidogenic and antioxidant activity of exopolysaccharides produced by Lactobacillus rhamnosus E/N cultivated on different carbon sources. Pol. J. Microbiol. 2013, 62, 181–188. [Google Scholar] [CrossRef]
- Zheng, B.A.; Zhu, Y.K.; Zheng, S.H.; Mo, Y.; Sun, S.L.; Ren, J.L.; Li, Y.H.; Wu, A.M.; Li, H.L. Upgrade the torrefaction process of bamboo based on autohydrolysis pretreatment. Ind. Crop Prod. 2021, 166, 113470. [Google Scholar] [CrossRef]
- Huang, J.W.; Zhu, Y.K.; Liu, T.Y.; Sun, S.L.; Ren, J.L.; Wu, A.M.; Li, H.L. A novel wet-mechanochemical pretreatment for the efficient enzymatic saccharification of lignocelluloses: Small dosage dilute alkali assisted ball milling. Energy Convers. Manag. 2019, 194, 46–54. [Google Scholar] [CrossRef]
- Rajoka, M.S.R.; Mehwish, H.M.; Hayat, H.F.; Hussain, N.; Sarwar, S.; Aslam, H.; Nadeem, A.; Shi, J.L. Characterization, the antioxidant and antimicrobial activity of exopolysaccharide isolated from poultry origin lactobacilli. Probiot. Antimicrob. Proteins 2019, 11, 1132–1142. [Google Scholar] [CrossRef]
- Smitinont, T.; Tansakul, C.; Tanasupawat, S.; Keeratipibul, S.; Navarini, L.; Bosco, M.; Cescutti, P. Exopolysaccharide-producing lactic acid bacteria strains from traditional thai fermented foods: Isolation, identification and exopolysaccharide characterization. Int. J. Food Microbiol. 1999, 51, 105–111. [Google Scholar] [CrossRef]
- Li, S.Y.; Ma, Y.L.; Ji, T.T.; Sameen, D.E.; Ahmed, S.; Qin, W.; Dai, J.W.; Li, S.Q.; Liu, Y.W. Cassava starch/carboxymethylcellulose edible films embedded with lactic acid bacteria to extend the shelf life of banana. Carbohydr. Polym. 2020, 248, 116805. [Google Scholar] [CrossRef]
- Abedfar, A.; Hosseininezhad, M.; Sadeghi, A.; Raeisi, M.; Feizy, J. Investigation on “spontaneous fermentation” and the productivity of microbial exopolysaccharides by Lactobacillus plantarum and Pediococcus pentosaceus isolated from wheat bran sourdough. LWT-Food Sci. Technol. 2018, 96, 686–693. [Google Scholar] [CrossRef]
- Jiang, S.M.; Cai, L.Z.; Lv, L.X.; Li, L.J. Pediococcus pentosaceus, a future additive or probiotic candidate. Microb. Cell. Fact. 2021, 20, 45. [Google Scholar] [CrossRef] [PubMed]
- Ayyash, M.; Abu-Jdayil, B.; Olaimat, A.; Esposito, G.; Itsaranuwat, P.; Osaili, T.; Obaid, R.; Kizhakkayil, J.; Liu, S.Q. Physicochemical, bioactive and rheological properties of an exopolysaccharide produced by a probiotic Pediococcus pentosaceus M41. Carbohydr. Polym. 2020, 229, 115462. [Google Scholar] [CrossRef] [PubMed]
- Abid, Y.; Casillo, A.; Gharsallah, H.; Joulak, I.; Lanzetta, R.; Corsaro, M.M.; Attia, H.; Azabou, S. Production and structural characterization of exopolysaccharides from newly isolated probiotic lactic acid bacteria. Int. J. Biol. Macromol. 2018, 108, 719–728. [Google Scholar] [CrossRef]
- Hussain, A.; Zia, K.M.; Tabasum, S.; Noreen, A.; Ali, M.; Iqbal, R.; Zuber, M. Blends and composites of exopolysaccharides; properties and applications: A review. Int. J. Biol. Macromol. 2017, 94, 10–27. [Google Scholar] [CrossRef] [PubMed]
- Kanmani, P.; Kumar, R.S.; Yuvaraj, N.; Paari, K.A.; Pattukumar, V.; Arul, V. Production and purification of a novel exopolysaccharide from lactic acid bacterium Streptococcus phocae PI80 and its functional characteristics activity in vitro. Bioresour. Technol. 2011, 102, 4827–4833. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, X.; Yang, Y.W.; Zhao, A.M.; Yang, Z.N. Characterization and bioactivities of an exopolysaccharide produced by Lactobacillus plantarum YW32. Int. J. Biol. Macromol. 2015, 74, 119–126. [Google Scholar] [CrossRef]
- Ahmed, Z.; Wang, Y.P.; Anjum, N.; Ahmad, H.; Ahmad, A.; Raza, M. Characterization of new exopolysaccharides produced by coculturing of L. kefiranofaciens with yoghurt strains. Int. J. Biol. Macromol. 2013, 59, 377–383. [Google Scholar] [CrossRef]
- Chen, Y.L.; Mao, W.J.; Wang, J.F.; Zhu, W.M.; Zhao, C.Q.; Li, N.; Wang, C.Y.; Yan, M.X.; Guo, T.; Liu, X. Preparation and structural elucidation of a glucomannogalactan from marine fungus Penicillium commune. Carbohydr. Polym. 2013, 97, 293–299. [Google Scholar] [CrossRef]
- Wang, K.; Li, W.; Rui, X.; Chen, X.H.; Jiang, M.; Dong, M.S. Structural characterization and bioactivity of released exopolysaccharides from Lactobacillus plantarum 70810. Int. J. Biol. Macromol. 2014, 67, 71–78. [Google Scholar] [CrossRef]
- Gomez-Pastor, R.; Perez-Torrado, R.; Cabiscol, E.; Ros, J.; Matallana, E. Reduction of oxidative cellular damage by overexpression of the thioredoxin TRX2 gene improves yield and quality of wine yeast dry active biomass. Microb. Cell. Fact. 2010, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.L.; Chen, X.Y.; Hu, P.; Liao, Q.W.; Luo, Y.; Li, J.; Feng, D.D.; Zhang, J.; Wu, Z.Q.; Xu, H.X. Extraction, purification, and antioxidant activity of exopolysaccharides produced by Lactobacillus kimchi SR8 from sour meat in vitro and in vivo. CyTA-J. Food 2021, 19, 228–237. [Google Scholar] [CrossRef]
- Zhang, S.W.; Liu, L.; Su, Y.L.; Li, H.J.; Sun, Q.; Liang, X.; Lv, J.P. Antioxidative activity of lactic acid bacteria in yogurt. Afr. J. Microbiol. Res. 2011, 5, 5194–5201. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Li, W.; Rui, X.; Li, T.; Chen, X.H.; Jiang, M.; Dong, M.S. Chemical modification, characterization and bioactivity of a released exopolysaccharide (r-EPS1) from Lactobacillus plantarum 70810. Glycoconj. J. 2015, 32, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Rani, R.P.; Anandharaj, M.; Ravindran, A.D. Characterization of a novel exopolysaccharide produced by Lactobacillus gasseri FR4 and demonstration of its in vitro biological properties. Int. J. Biol. Macromol. 2018, 109, 772–783. [Google Scholar] [CrossRef] [PubMed]
- Seo, B.J.; Bajpai, V.K.; Rather, I.A.; Park, Y.H. Partially Purified exopolysaccharide from Lactobacillus plantarum YML009 with total phenolic content, antioxidant and free radical scavenging efficacy. Indian J. Pharm. Educ. Res. 2015, 49, 282–292. [Google Scholar] [CrossRef] [Green Version]
- Xiao, L.Y.; Han, S.; Zhou, J.Z.; Xu, Q.; Dong, M.S.; Fan, X.; Rui, X.; Chen, X.H.; Zhang, Q.Q.; Li, W. Preparation, characterization and antioxidant activities of derivatives of exopolysaccharide from Lactobacillus helveticus MB2-1. Int. J. Biol. Macromol. 2020, 145, 1008–1017. [Google Scholar] [CrossRef]
- Tang, W.Z.; Dong, M.S.; Wang, W.L.; Han, S.; Rui, X.; Chen, X.H.; Jiang, M.; Zhang, Q.Q.; Wu, J.J.; Li, W. Structural characterization and antioxidant property of released exopolysaccharides from Lactobacillus delbrueckii ssp bulgaricus SRFM-1. Carbohydr. Polym. 2017, 173, 654–664. [Google Scholar] [CrossRef]
- Nambiar, R.B.; Sellamuthu, P.S.; Perumal, A.B.; Sadiku, E.R.; Phiri, G.; Jayaramudu, J. Characterization of an exopolysaccharide produced by Lactobacillus plantarum HM47 isolated from human breast milk. Process Biochem. 2018, 73, 15–22. [Google Scholar] [CrossRef]
- Wang, K.; Niu, M.M.; Song, D.W.; Song, X.J.; Zhao, J.; Wu, Y.; Lu, B.X.; Niu, G.C. Preparation, partial characterization and biological activity of exopolysaccharides produced from Lactobacillus fermentum S1. J. Biosci. Bioeng. 2020, 129, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Xing, K.; Chen, X.G.; Kong, M.; Liu, C.S.; Cha, D.S.; Park, H.J. Effect of oleoyl-chitosan nanoparticles as a novel antibacterial dispersion system on viability, membrane permeability and cell morphology of Escherichia coli and Staphylococcus aureus. Carbohydr. Polym. 2009, 76, 17–22. [Google Scholar] [CrossRef]
- Spano, A.; Lagana, P.; Visalli, G.; Maugeri, T.L.; Gugliandolo, C. In vitro antibiofilm activity of an exopolysaccharide from the marine thermophilic bacillus licheniformis T14. Curr. Microbiol. 2016, 72, 518–528. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source Silage | Strains | Yield (mg/L in MRS Broth, Mean ± SD) |
|---|---|---|
| Napier grass | SPP–1 | 59.3 ± 2.9 l |
| SPP–2 | 67.9 ± 4.9 kl | |
| SPP–3 | 62.4 ± 5.1 l | |
| SPP–5 | 87.5 ± 6.5 jk | |
| SPP–6 | 202.5 ± 17.9 bc | |
| SSP–9 | 172.6 ± 27.2 def | |
| Corn | SSC–1 | 51.0 ± 5.0 l |
| SSC–3 | 200.5 ± 16.3 bc | |
| SSC–12 | 276.6 ± 12.1 a | |
| SSC–16 | 110.6 ± 6.3 i | |
| SSC–23 | 212.3 ± 10.5 b | |
| SSC–48 | 143.7 ± 16.7 gh | |
| Orchardgrass | SDJ–3 | 30.5 ± 5.2 m |
| SDJ–6 | 188.5 ± 9.0 cd | |
| SDJ–8 | 203.7 ± 8.2 bc | |
| SDJ–16 | 131.1 ± 10.0 h | |
| Stylo | SS–9 | 90.5 ± 4.9 j |
| SS–17 | 181.7 ± 10.2 cde | |
| Soybean | SGM–9 | 202.9 ± 11.5 bc |
| SGM–16 | 164.8 ± 15.8 ef | |
| SGM–18 | 176.8 ± 19.9 de | |
| SGM–20 | 153.2 ± 7.3 fg |
| Monosaccharide Name | Molar Ratio (%) |
|---|---|
| Glucose | 42.7 ± 0.28 |
| Mannose | 28.9 ± 0.39 |
| Galactose | 16.3 ± 0.06 |
| Arabinose | 9.4 ± 0.36 |
| Rhamnose | 2.9 ± 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, Y.; Li, X.; Tian, R.; Tang, R.; Zhang, J. Characterization and Biological Activity of a Novel Exopolysaccharide Produced by Pediococcus pentosaceus SSC–12 from Silage. Microorganisms 2022, 10, 18. https://doi.org/10.3390/microorganisms10010018
Fan Y, Li X, Tian R, Tang R, Zhang J. Characterization and Biological Activity of a Novel Exopolysaccharide Produced by Pediococcus pentosaceus SSC–12 from Silage. Microorganisms. 2022; 10(1):18. https://doi.org/10.3390/microorganisms10010018
Chicago/Turabian StyleFan, Yang, Xinqin Li, Rong Tian, Ruxue Tang, and Jianguo Zhang. 2022. "Characterization and Biological Activity of a Novel Exopolysaccharide Produced by Pediococcus pentosaceus SSC–12 from Silage" Microorganisms 10, no. 1: 18. https://doi.org/10.3390/microorganisms10010018
APA StyleFan, Y., Li, X., Tian, R., Tang, R., & Zhang, J. (2022). Characterization and Biological Activity of a Novel Exopolysaccharide Produced by Pediococcus pentosaceus SSC–12 from Silage. Microorganisms, 10(1), 18. https://doi.org/10.3390/microorganisms10010018