Occurrence of Chlamydiaceae in Raptors and Crows in Switzerland

 ,
, 
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Chlamydiaceae 23S rRNA qPCR
2.1.1. Species
2.1.2. Geographical Origin
2.1.3. Swab Type
2.2. C. Psittaci qPCR and C. Buteonis qPCR
2.3. 16S rRNA Conventional PCR and Sequencing
2.4. Outer Membrane Protein A (ompA) Genotyping
3. Discussion
3.1. Corvids
3.2. Raptors
3.3. Geographical Distribution
3.4. Swab Types
3.5. Public Health Concerns
4. Materials and Methods
4.1. Samples
4.2. DNA Extraction
4.3. Chlamydiaceae 23S rRNA qPCR
4.4. C. Psittaci OmpA qPCR
4.5. C. Buteonis OxaA qPCR
4.6. 16S rRNA PCR and Sequencing
4.7. OmpA Genotyping PCR
4.8. Statistical Analysis
4.9. Ethical Statement
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sachse, K.; Laroucau, K.; Vanrompay, D. Avian chlamydiosis. Curr. Clin. Micro. Rpt. 2015, 2, 10–21. [Google Scholar] [CrossRef] [Green Version]
- Cheong, H.C.; Lee, C.Y.Q.; Cheok, Y.Y.; Tan, G.M.Y.; Looi, C.Y.; Wong, W.F. Chlamydiaceae: Diseases in primary hosts and zoonosis. Microorganisms 2019, 7, 146. [Google Scholar] [CrossRef] [Green Version]
- Laroucau, K.; Vorimore, F.; Aaziz, R.; Solmonson, L.; Hsia, R.C.; Bavoil, P.M.; Fach, P.; Hölzer, M.; Wuenschmann, A.; Sachse, K. Chlamydia buteonis, a new Chlamydia species isolated from a red-shouldered hawk. Syst. Appl. Microbiol. 2019, 42, 125997. [Google Scholar] [CrossRef]
- Sachse, K.; Bavoil, P.M.; Kaltenboeck, B.; Stephens, S.S.; Kuo, C.C.; Rosselló-Móra, R.; Horn, M. Emendation of the family Chlamydiaceae: Proposal of a single genus, Chlamydia, to include all currently recognized species. Syst. Appl. Microbiol. 2015, 38, 99–103. [Google Scholar] [CrossRef]
- Kaleta, E.F.; Taday, E.M. Avian host range of Chlamydophila spp. based on isolation, antigen detection and serology. Avian Pathol. 2003, 32, 435–461. [Google Scholar] [CrossRef] [Green Version]
- Sachse, K.; Kuehlewind, S.; Ruettger, A.; Schubert, E.; Rohde, G. More than classical Chlamydia psittaci in urban pigeons. Vet. Microbiol. 2012, 157, 476–480. [Google Scholar] [CrossRef]
- Vorimore, F.; Hsia, R.C.; Huot-Creasy, H.; Bastian, S.; Deruyter, L.; Passet, A.; Sachse, K.; Bavoil, P.; Myers, G.; Laroucau, K. Isolation of a new Chlamydia species from the feral sacred ibis (Threskiornis aethiopicus): Chlamydia ibidis. PLoS ONE 2013, 8, e74823. [Google Scholar] [CrossRef] [Green Version]
- Di Francesco, A.; Donati, M.; Laroucau, K.; Balboni, A.; Galuppi, R.; Merialdi, G.; Salvatore, D.; Renzi, M. Chlamydiae in corvids. Vet. Rec. 2015, 177, 466. [Google Scholar] [CrossRef]
- Guo, W.; Li, J.; Kaltenboeck, B.; Gong, J.; Fan, W.; Wang, C. Chlamydia gallinacea, not C. psittaci, is the endemic chlamydial species in chicken (Gallus gallus). Sci. Rep. 2016, 6, 19638. [Google Scholar] [CrossRef]
- Szymańska-Czerwińska, M.; Mitura, A.; Niemczuk, K.; Zaręba, K.; Jodełko, A.; Pluta, A.; Scharf, S.; Vitek, B.; Aaziz, R.; Vorimore, F.; et al. Dissemination and genetic diversity of chlamydial agents in Polish wildfowl: Isolation and molecular characterisation of avian Chlamydia abortus strains. PLoS ONE 2017, 12, e0174599. [Google Scholar] [CrossRef] [Green Version]
- Stokes, H.S.; Martens, J.M.; Chamings, A.; Walder, K.; Berg, M.L.; Segal, Y.; Bennet, A. Identification of Chlamydia gallinacea in a parrot and in free-range chickens in Australia. Aust. Vet. J. 2019, 97, 398–400. [Google Scholar] [CrossRef]
- Bundesamt für Lebensmittelsicherheit und Veterinärwesen (BLV). Informationssystem Seuchenmeldungen InfoSM. Available online: https://www.infosm.blv.admin.ch (accessed on 3 April 2020).
- Borel, N.; Polkinghorne, A.; Pospischil, A. A review on chlamydial diseases in animals: Still a challenge for pathologists? Vet. Pathol. 2018, 55, 374–390. [Google Scholar] [CrossRef] [Green Version]
- Knittler, M.R.; Sachse, K. Chlamydia psittaci: Update on an underestimated zoonotic agent. Pathog. Dis. 2015, 73, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Laroucau, K.; Vorimore, F.; Aaziz, R.; Berndt, A.; Schubert, E.; Sachse, K. Isolation of a new chlamydial agent from infected domestic poultry coincided with cases of atypical pneumonia among slaughterhouse workers in France. Infect. Genet. Evol. 2009, 9, 1240–1247. [Google Scholar] [CrossRef]
- Sachse, K.; Laroucau, K.; Riege, K.; Wehner, S.; Dilcher, M.; Creasy, H.H.; Weidmann, M.; Myers, G.; Vorimore, F.; Vicari, N.; et al. Evidence of the existence of two new members of the family Chlamydiaceae and proposal of Chlamydia avium sp. nov. and Chlamydia gallinacea sp. nov. Syst. Appl. Microbiol. 2014, 37, 79–88. [Google Scholar] [CrossRef]
- Zweifel, D.; Hoop, R.; Sachse, K.; Pospischil, A.; Borel, N. Prevalence of Chlamydophila psittaci in wild birds—Potential risk for domestic poultry, pet birds, and public health? Eur. J. Wildlife Res. 2009, 55, 575–581. [Google Scholar] [CrossRef]
- Mattmann, P.; Marti, H.; Borel, N.; Jelocnik, M.; Albini, S.; Vogler, B.R. Chlamydiaceae in wild, feral and domestic pigeons in Switzerland and insight into population dynamics by Chlamydia psittaci multilocus sequence typing. PLoS ONE 2019, 14, e0226088. [Google Scholar] [CrossRef] [Green Version]
- Blomqvist, M.; Christerson, L.; Waldenström, J.; Lindberg, P.; Helander, B.; Gunnarsson, G.; Herrmann, B.; Olsen, B. Chlamydia psittaci in birds of prey, Sweden. Infect. Ecol. Epidemiol. 2012, 2. [Google Scholar] [CrossRef]
- Gerbermann, H.; Korbel, R. The occurrence of Chlamydia psittaci infections in raptors from wildlife preserves. Tierarztl. Prax. 1993, 21, 217–224. [Google Scholar]
- Schettler, E.; Fickel, J.; Hotzel, H.; Sachse, K.; Streich, W.J.; Wittstatt, U.; Frölich, K. Newcastle disease virus and Chlamydia psittaci in free-living raptors from eastern Germany. J. Wildl. Dis. 2003, 39, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Fukushi, H.; Hirai, K. Immunochemical diversity of the major outer membrane protein of avian and mammalian Chlamydia psittaci. J. Clin. Microbiol. 1988, 26, 675–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanrompay, D.; Butaye, P.; Sayada, C.; Ducatelle, R.; Haesebrouck, F. Characterization of avian Chlamydia psittaci strains using omp1 restriction mapping and serovar-specific monoclonal antibodies. Res. Microbiol. 1997, 148, 327–333. [Google Scholar] [CrossRef]
- Herrmann, B.; Rahman, R.; Bergström, S.; Bonnedahl, J.; Olsen, B. Chlamydophila abortus in a brown skua (Catharacta antarctica lonnbergi) from a subantarctic island. Appl. Environ. Microbiol. 2000, 66, 3654–3656. [Google Scholar] [CrossRef] [Green Version]
- van Loock, M.; Vanrompay, D.; Herrmann, B.; vander Stappen, J.; Volckaert, G.; Goddeeris, B.M.; Everett, K.D.E. Missing links in the divergence of Chlamydophila abortus from Chlamydophila psittaci. Int. J. Syst. Evol. Microbiol. 2003, 53, 761–770. [Google Scholar] [CrossRef]
- Madani, S.A.; Peighambari, S.M. PCR-based diagnosis, molecular characterization and detection of atypical strains of avian Chlamydia psittaci in companion and wild birds. Avian Pathol. 2013, 42, 38–44. [Google Scholar] [CrossRef] [Green Version]
- Aaziz, R.; Gourlay, P.; Vorimore, F.; Sachse, K.; Siarkou, V.I.; Laroucau, K. Chlamydiaceae in North Atlantic seabirds admitted to a wildlife rescue center in western France. Appl. Environ. Microbiol. 2015, 81, 4581–4590. [Google Scholar] [CrossRef] [Green Version]
- Krawiec, M.; Piasecki, T.; Wieliczko, A. Prevalence of Chlamydia psittaci and other Chlamydia species in wild birds in Poland. Vector Borne Zoonotic Dis. 2015, 15, 652–655. [Google Scholar] [CrossRef] [Green Version]
- Luján-Vega, C.; Hawkins, M.G.; Johnson, C.K.; Briggs, C.; Vennum, C.; Bloom, P.H.; Hull, J.M.; Cray, C.; Pesti, D.; Johnson, L.; et al. Atypical Chlamydiaceae in wild populations of hawks (Buteo spp.) in California. J. Zoo. Wildl. Med. 2018, 49, 108–115. [Google Scholar] [CrossRef]
- Liu, S.Y.; Li, K.P.; Hsieh, M.K.; Chang, P.C.; Shien, J.H.; Ou, S.C. Prevalence and genotyping of Chlamydia psittaci from domestic waterfowl, companion birds, and wild birds in Taiwan. Vector Borne Zoonotic Dis. 2019, 19, 666–673. [Google Scholar] [CrossRef]
- Mirandé, L.A.; Howerth, E.W.; Poston, R.P. Chlamydiosis in a red-tailed hawk (Buteo jamaicensis). J. Wildl. Dis. 1992, 28, 284–287. [Google Scholar] [CrossRef]
- Joseph, S.J.; Marti, H.; Didelot, X.; Castillo-Ramirez, S.; Read, T.D.; Dean, D. Chlamydiaceae genomics reveals interspecies admixture and the recent evolution of Chlamydia abortus infecting lower mammalian species and humans. Genome Biol. Evol. 2015, 7, 3070–3084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrmann, B.; Persson, H.; Jensen, J.K.; Joensen, H.D.; Klint, M.; Olsen, B. Chlamydophila psittaci in Fulmars, the Faroe Islands. Emerg. Infect. Dis. 2006, 12, 330–332. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; An, I.; Oem, J.K.; Wang, S.J.; Kim, Y.; Shin, J.H.; Woo, C.; Kim, Y.; Jo, S.D.; Son, K.; et al. Molecular prevalence and genotyping of Chlamydia spp. in wild birds from South Korea. J. Vet. Med. Sci. 2017, 79, 1204–1209. [Google Scholar] [CrossRef] [Green Version]
- Yatsentyuk, S.P.; Obukhov, I.L. Molecular genetic characterization of avian Chlamydophila psittaci isolates. Russ. J. Genet. 2007, 43, 1454–1460. [Google Scholar] [CrossRef]
- Dickx, V.; Geens, T.; Deschuyffeleer, T.; Tyberghien, L.; Harkinezhad, T.; Beeckman, D.S.A.; Braeckman, L.; Vanrompay, D. Chlamydophila psittaci zoonotic risk assessment in a chicken and turkey slaughterhouse. J. Clin. Microbiol. 2010, 48, 3244–4250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konicek, C.; Vodrážka, P.; Barták, P.; Knotek, Z.; Hess, C.; Račka, K.; Hess, M.; Troxler, S. Detection of zoonotic pathogens in wild birds in the cross-border region Austria—Czech Republic. J. Wildl. Dis. 2016, 52, 850–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pannekoek, Y.; Dickx, V.; Beeckman, D.S.A.; Jolley, K.A.; Keijzers, W.C.; Vretou, E.; Maiden, M.C.J.; Vanrompay, D.; van der Ende, A. Multi locus sequence typing of Chlamydia reveals an association between Chlamydia psittaci genotypes and host species. PLoS ONE 2010, 5, e14179. [Google Scholar] [CrossRef] [PubMed]
- Sachse, K.; Ruettger, A. Rapid microarray-based genotyping of Chlamydia spp. strains from clinical tissue samples. Methods Mol. Biol. 2015, 1247, 391–400. [Google Scholar] [CrossRef]
- Andersen, A.A. Comparison of pharyngeal, fecal, and cloacal samples for the isolation of Chlamydia psittaci from experimentally infected cockatiels and turkeys. J. Vet. Diagn. Investig. 1996, 8, 448–450. [Google Scholar] [CrossRef] [Green Version]
- Čechová, L.; Halánová, M.; Babinská, I.; Danišová, O.; Bartkovský, M.; Marcinčák, S.; Marcinčáková, D.; Valenčáková, A.; Čisláková, L. Chlamydiosis in farmed chickens in Slovakia and zoonotic risk for humans. Ann. Agric. Environ. Med. 2018, 25, 320–325. [Google Scholar] [CrossRef]
- Yin, L.; Lagae, S.; Kalmar, I.; Borel, N.; Pospischil, A.; Vanrompay, D. Pathogenicity of low and highly virulent Chlamydia psittaci isolates for specific-pathogen-free chickens. Avian Dis. 2013, 57, 242–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Buuren, C.E.; Dorrestein, G.M.; van Dijk, J.E. Chlamydia psittaci infections in birds: A review on the pathogenesis and histopathological features. Vet. Q. 1994, 16, 38–41. [Google Scholar] [CrossRef] [PubMed]
- Beeckman, D.S.; Vanrompay, D.C. Zoonotic Chlamydophila psittaci infections from a clinical perspective. Clin. Microbiol. Infect. 2009, 15, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Bavoil, P.; Kaltenboeck, B.; Greub, G. In Chlamydia veritas. Pathog. Dis. 2013, 67, 89–90. [Google Scholar] [CrossRef]
- Heddema, E.R.; van Hannen, E.J.; Bongaerts, M.; Dijkstra, F.; Ten Hove, R.J.; de Wever, B.; Vanrompay, D. Typing of Chlamydia psittaci to monitor epidemiology of psittacosis and aid disease control in the Netherlands, 2008 to 2013. Euro Surveill. 2015, 20, 21026. [Google Scholar] [CrossRef] [Green Version]
- Carlier, L.; Kempf, M.; Aaziz, R.; Jolivet-Gougeon, A.; Laroucau, K. A severe case of pneumopathy in a duck breeder due to Chlamydia psittaci diagnosed by 16S rDNA sequencing. JMM Case Rep. 2014, 1. [Google Scholar] [CrossRef] [Green Version]
- Radomski, N.; Einenkel, R.; Müller, A.; Knittler, M.R. Chlamydia-host cell interaction not only from a bird’s eye view: Some lessons from Chlamydia psittaci. FEBS Lett. 2016, 590, 3920–3940. [Google Scholar] [CrossRef]
- Spoorenberg, S.M.; Bos, W.J.; van Hannen, E.J.; Dijkstra, F.; Heddema, E.R.; van Velzen-Blad, H.; Heijligenberg, R.; Grutters, J.C.; de Jongh, B.M.; Ovidius Study Group. Chlamydia psittaci: A relevant cause of community-acquired pneumonia in two Dutch hospitals. Neth. J. Med. 2016, 74, 75–81. [Google Scholar]
- Hogerwerf, L.; de Gier, B.; Baan, B.; van der Hoek, W. Chlamydia psittaci (psittacosis) as a cause of community-acquired pneumonia: A systematic review and meta-analysis. Epidemiol. Infect. 2017, 145, 3096–3105. [Google Scholar] [CrossRef] [Green Version]
- De Gier, B.; Hogerwerf, L.; Dijkstra, F.; van der Hoek, W. Disease burden of psittacosis in the Netherlands. Epidemiol. Infect. 2018, 146, 303–305. [Google Scholar] [CrossRef] [Green Version]
- Rybarczyk, J.; Versteele, C.; Lernout, T.; Vanrompay, D. Human psittacosis: A review with emphasis on surveillance in Belgium. Acta Clin. Belg. 2020, 75, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Telfer, B.L.; Moberley, S.A.; Hort, K.P.; Branley, J.M.; Dwyer, D.E.; Muscatello, D.J.; Correll, P.K.; England, J.; McAnulty, J.M. Probable psittacosis outbreak linked to wild birds. Emerg. Infect. Dis. 2005, 11, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Rehn, M.; Ringberg, H.; Runehagen, A.; Herrmann, B.; Olsen, B.; Petersson, A.C.; Hjertqvist, M.; Kühlmann-Berenzon, S.; Wallensten, A. Unusual increase of psittacosis in southern Sweden linked to wild bird exposure, January to April 2013. Euro Surveill. 2013, 18, 20478. [Google Scholar] [PubMed]
- Chereau, F.; Rehn, M.; Pini, A.; Kühlmann-Berenzon, S.; Ydring, E.; Ringberg, H.; Runehagen, A.; Ockborn, G.; Dotevall, L.; Wallensten, A. Wild and domestic bird faeces likely source of psittacosis transmission-a case-control study in Sweden, 2014-2016. Zoonoses Public Health 2018, 65, 790–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haagen, E.; Maurer, G. Ueber eine auf den Menschen übertragbare Viruskrankheit bei Sturmvögeln und ihre Beziehung zur Psittakose. Zentralblatt für Bakteriologie, Parasitenkunde und Infektionskrankheiten. Erste Abt. Orig. 1938, 143, 81–88. [Google Scholar]
- Filstein, M.R.; Ley, A.B.; Vernon, M.S.; Gaffney, K.A.; Glickman, L.T. Epidemic of psittacosis in college of veterinary medicine. J. Am. Vet. Med. Assoc. 1981, 179, 569–572. [Google Scholar]
- Schlossberg, D.; Delgado, J.; Moore, M.M.; Wishner, A.; Mohn, J. An epidemic of avian and human psittacosis. Arch. Intern. Med. 1993, 153, 2594–2596. [Google Scholar] [CrossRef]
- Davies, A.; Collins, T. Respiratory Chlamydia: The management of an outbreak. Public Health 1995, 109, 207–211. [Google Scholar] [CrossRef]
- Gosbell, I.B.; Ross, A.D.; Turner, I.B. Chlamydia psittaci infection and reinfection in a veterinarian. Aust. Vet. J. 1999, 77, 511–513. [Google Scholar] [CrossRef]
- Saito, T.; Ohnishi, J.; Mori, Y.; Iinuma, Y.; Ichiyama, S.; Kohi, F. Infection by Chlamydophila avium in an elderly couple working in a pet shop. J. Clin. Microbiol. 2005, 43, 3011–3013. [Google Scholar] [CrossRef] [Green Version]
- Raso, T.F.; Carrasco, A.O.; Silva, J.C.; Marvulo, M.F.; Pinto, A.A. Seroprevalence of antibodies to Chlamydophila psittaci in zoo workers in Brazil. Zoonoses Public Health 2010, 57, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Tolba, H.M.N.; Abou Elez, R.M.M.; Elsohaby, I. Risk factors associated with Chlamydia psittaci infections in psittacine birds and bird handlers. J. Appl. Microbiol. 2019, 126, 402–410. [Google Scholar] [CrossRef]
- Ehricht, R.; Slickers, P.; Goellner, S.; Hotzel, H.; Sachse, K. Optimized DNA microarray assay allows detection and genotyping of single PCR-amplifiable target copies. Mol. Cell. Probes. 2006, 20, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, B.; Depner, K.; Schirrmeier, H.; Beer, M. A universal heterologous internal control system for duplex real-time RT-PCR assays used in a detection system for pestiviruses. J. Virol. Methods. 2006, 136, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Blumer, S.; Greub, G.; Waldvogel, A.; Hässig, M.; Thoma, R.; Tschuor, A.; Pospischil, A.; Borel, N. Waddlia, Parachlamydia and Chlamydiaceae in bovine abortion. Vet. Microbiol. 2011, 152, 385–393. [Google Scholar] [CrossRef] [Green Version]
- Pantchev, A.; Sting, R.; Bauerfeind, R.; Tyczka, J.; Sachse, K. New real-time PCR tests for species-specific detection of Chlamydophila psittaci and Chlamydophila abortus from tissue samples. Vet. J. 2009, 181, 145–150. [Google Scholar] [CrossRef]
- Hoffmann, B.; Beer, M.; Schelp, C.; Schirrmeier, H.; Depner, K. Validation of a real-time RT-PCR assay for sensitive and specific detection of classical swine fever. J. Virol. Methods 2005, 130, 36–44. [Google Scholar] [CrossRef]
- Everett, K.D.; Bush, R.M.; Andersen, A.A. Emended description of the order Chlamydiales, proposal of Parachlamydiaceae fam. nov. and Simkaniaceae fam. nov., each containing one monotypic genus, revised taxonomy of the family Chlamydiaceae, including a new genus and five new species, and standards for the identification of organisms. Int. J. Syst. Bacteriol. 1999, 49, 415–440. [Google Scholar] [CrossRef]
- Blumer, C.; Zimmermann, D.R.; Weilenmann, R.; Vaughan, L.; Pospischil, A. Chlamydiae in free-ranging and captive frogs in Switzerland. Vet. Pathol. 2007, 44, 144–150. [Google Scholar] [CrossRef] [Green Version]
- Taylor-Brown, A.; Rüegg, S.; Polkinghorne, A.; Borel, N. Characterisation of Chlamydia pneumoniae and other novel chlamydial infections in captive snakes. Vet. Microbiol. 2015, 178, 88–93. [Google Scholar] [CrossRef]
- Hosokawa, T.; Kikuchi, Y.; Nikoh, N.; Shimada, M.; Fukatsu, T. Strict host-symbiont cospeciation and reductive genome evolution in insect gut bacteria. PLoS Biol. 2006, 4, e337. [Google Scholar] [CrossRef]
- Sachse, K.; Laroucau, K.; Hotzel, H.; Schubert, E.; Ehricht, R.; Slickers, P. Genotyping of Chlamydophila psittaci using a new DNA microarray assay based on sequence analysis of ompA genes. BMC Microbiol. 2008, 8, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]


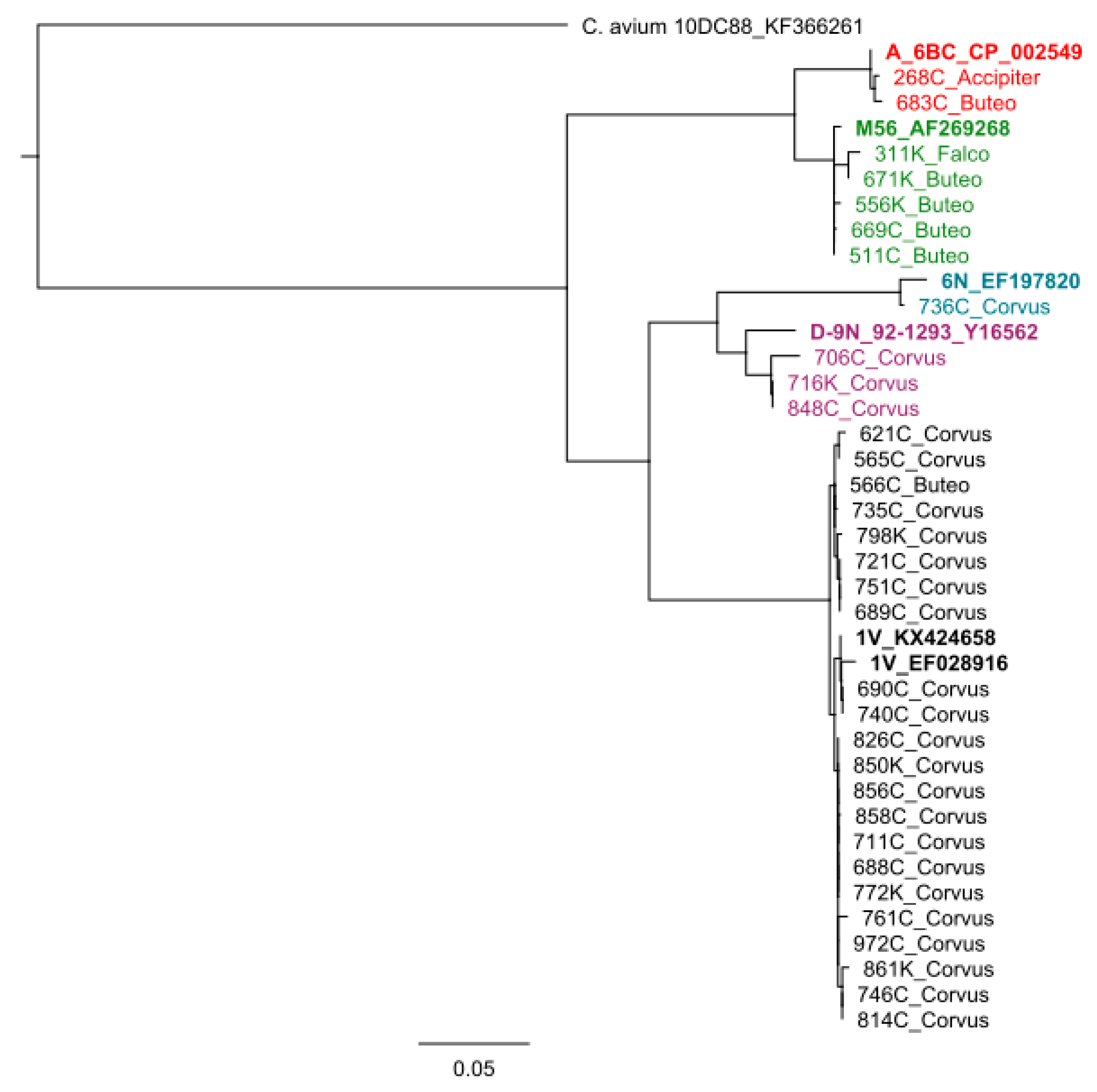{kind=link}
| Species Name | Chlamydiaceae qPCR Pos. (%) | Final Classification | ||||||
|---|---|---|---|---|---|---|---|---|
| C. Abortus/ C. Psittaci (%) | C. Psittaci M56 (%) | C. Psittaci A (%) | C. Psittaci 6N (%) | C. Psittaci 1V (%) | C. Psittaci D (%) | Not Further Identified (%) | ||
| Bearded vulture | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Black kite | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Common buzzard | 13 (9.2%) | 0 | 5 (38.5%) | 1 (7.7%) | 0 | 1 (7.7%) | 0 | 6 (46.2%) |
| Eurasian sparrowhawk | 3 (9.4%) | 0 | 0 | 1 (33.3%) | 0 | 0 | 0 | 2 (66.7%) |
| European honey-buzzard | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Golden eagle | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Montagu’s harrier | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Northern goshawk | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Red kite | 1 (4.3%) | 0 | 0 | 0 | 0 | 0 | 0 | 1 (100%) |
| Accipitridae subtotal | 17 (7.9%) | 0 | 5 (29.4%) | 2 (11.8%) | 0 | 1 (5.9%) | 0 | 9 (52.9%) |
| Common kestrel | 1 (1.5%) | 0 | 1 (100%) | 0 | 0 | 0 | 0 | 0 |
| Eurasian hobby | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Peregrine falcon | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Falconidae subtotal | 1 (1.4%) | 0 | 1 (100%) | 0 | 0 | 0 | 0 | 0 |
| Eurasian eagle-owl | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Long-eared owl | 1 (5.9%) | 0 | 1 (100%) | 0 | 0 | 0 | 0 | 0 |
| Tawny owl | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Strigidae subtotal | 1 (2.6%) | 0 | 1 (100%) | 0 | 0 | 0 | 0 | 0 |
| Barn owl | 1 (5.9%) | 0 | 0 | 0 | 0 | 0 | 0 | 1 (100%) |
| Tytonidae subtotal | 1 (5.9%) | 0 | 0 | 0 | 0 | 0 | 0 | 1 (100%) |
| Carrion crow | 59 (28.5%) | 23 (39.0%) | 0 | 0 | 1 (1.7%) | 21 (35.6%) | 3 (5.1%) | 11 (18.6%) |
| Eurasian jay | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Eurasian magpie | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Hooded crow | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Rook | 1 (33.3%) | 0 | 0 | 0 | 0 | 1 (100%) | 0 | 0 |
| Western jackdaw | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Corvidae subtotal | 60 (23.7%) | 23 (38.3%) | 0 | 0 | 1 (1.7%) | 22 (36.7%) | 3 (5.0%) | 11 (18.3%) |
| Greater Region | Swiss Canton | Number of Birds | Chlamydiaceae Positive (%) |
|---|---|---|---|
| Lake Geneva | Geneva | 15 | 0 |
| Valais | 11 | 0 | |
| Espace Mittelland | Bern | 56 | 5 (8.9%) |
| Fribourg | 18 | 1 (5.6%) | |
| Solothurn | 13 | 0 | |
| Northwestern Switzerland | Aargau | 31 | 1 (3.2%) |
| Basel District | 4 | 0 | |
| Zurich | Zurich | 132 | 32 (24.2%) |
| Eastern Switzerland | Glarus | 1 | 0 |
| Grisons | 10 | 0 | |
| Schaffhausen | 9 | 1 (11.1%) | |
| St. Gallen | 6 | 0 | |
| Thurgau | 14 | 1 (7.1%) | |
| Central Switzerland | Lucerne | 115 | 2 (1.7%) |
| Nidwalden | 3 | 0 | |
| Obwalden | 5 | 1 (20.0%) | |
| Uri | 1 | 0 | |
| Zug | 51 | 27 (52.9%) | |
| Ticino | Ticino | 6 | 0 |
| Unknown | Unknown | 93 | 9 (9.7%) |
| Choanal Swabs Positive/Total (%) | Cloacal Swabs Positive/Total (%) | Fecal Swabs Positive/Total (%) | |
|---|---|---|---|
| Raptors | 15/299 (5.0%) | 16/304 (5.3%) | 0/42 (0%) |
| Corvids | 57/220 (25.9%) | 30/216 (13.9%) | 1/47 (2.1%) |
| Total | 72/519 (13.9%) | 46/520 (8.8%) | 1/89 (1.1%) |
| Sample Nr. | Species Name (English) | Sequence Length (bp) | Sequence Quality (%) | First Hit | Nucleotide Identity (%) | Accession Number |
|---|---|---|---|---|---|---|
| Raptors | ||||||
| 14C | Common buzzard | 245 | 26.1 | C. psittaci M56 | 99.59 | MT423441 |
| 311K | Common kestrel | 269 | 80.3 | C. psittaci M56 | 100 | MT423442 |
| 556C | Common buzzard | 239 | 30.1 | C. psittaci M56 | 100 | MT423443 |
| 556K | Common buzzard | 278 | 83.1 | C. psittaci M56 | 100 | MT423444 |
| 566C | Common buzzard | 278 | 80.9 | C. abortus 15-58d44 | 100 | MT423446 |
| 669K | Common buzzard | 271 | 83.0 | C. psittaci M56 | 100 | MT423448 |
| 671C | Common buzzard | 260 | 32.7 | C. psittaci M56 | 98.85 | MT423449 |
| 671K | Common buzzard | 269 | 77.3 | C. psittaci M56 | 100 | MT423450 |
| Corvids | ||||||
| 565C | Carrion crow | 253 | 82.2 | C. abortus 15-58d44 | 100 | MT423445 |
| 621C | Rook | 274 | 75.2 | C. abortus 15-58d44 | 100 | MT423447 |
| 686C | Carrion crow | 271 | 40.6 | C. abortus 15-58d44 | 99.63 | MT423451 |
| 688C | Carrion crow | 252 | 80.2 | C. abortus 15-58d44 | 100 | MT423452 |
| 688K | Carrion crow | 279 | 73.8 | C. abortus 15-58d44 | 100 | MT423453 |
| 689C | Carrion crow | 271 | 79.3 | C. abortus 15-58d44 | 100 | MT423454 |
| 689K | Carrion crow | 249 | 25.7 | C. abortus 15-58d44 | 95.58 | MT423455 |
| 690C | Carrion crow | 253 | 80.6 | C. abortus 15-58d44 | 100 | MT423456 |
| 696C | Carrion crow | 276 | 78.3 | C. abortus 15-58d44 | 100 | MT423457 |
| 702C | Carrion crow | 275 | 80.7 | C. abortus 15-58d44 | 100 | MT423458 |
| 705C | Carrion crow | 278 | 71.6 | C. abortus 15-58d44 | 100 | MT423459 |
| 706C | Carrion crow | 256 | 80.1 | C. abortus 15-58d44 | 100 | MT423460 |
| 711C | Carrion crow | 260 | 85.4 | C. abortus 15-58d44 | 100 | MT423461 |
| 716K | Carrion crow | 272 | 84.2 | C. abortus 15-58d44 | 100 | MT423462 |
| 721C | Carrion crow | 253 | 81.4 | C. abortus 15-58d44 | 100 | MT423463 |
| 725C | Carrion crow | 267 | 81.3 | C. abortus 15-58d44 | 100 | MT423464 |
| 735C | Carrion crow | 267 | 76.8 | C. abortus 15-58d44 | 100 | MT423465 |
| 736C | Carrion crow | 253 | 80.6 | C. abortus 15-58d44 | 100 | MT423466 |
| 736K | Carrion crow | 271 | 76.8 | C. abortus 15-58d44 | 100 | MT423467 |
| 737C | Carrion crow | 275 | 77.5 | C. abortus 15-58d44 | 100 | MT423468 |
| 737K | Carrion crow | 315 | 63.8 | C. abortus 15-58d44 | 96.96 | MT423469 |
| 740C | Carrion crow | 260 | 82.7 | C. abortus 15-58d44 | 100 | MT423470 |
| 740K | Carrion crow | 260 | 73.5 | C. abortus 15-58d44 | 100 | MT423471 |
| 744C | Carrion crow | 278 | 77.3 | C. abortus 15-58d44 | 100 | MT423472 |
| 746C | Carrion crow | 266 | 84.6 | C. abortus 15-58d44 | 100 | MT423473 |
| 750C | Carrion crow | 276 | 76.4 | C. abortus 15-58d44 | 100 | MT423474 |
| 750K | Carrion crow | 250 | 21.2 | C. abortus 15-58d44 | 98.40 | MT423475 |
| 751C | Carrion crow | 269 | 85.9 | C. abortus 15-58d44 | 100 | MT423476 |
| 751K | Carrion crow | 226 | 32.7 | C. abortus 15-58d44 | 100 | MT423477 |
| 752C | Carrion crow | 278 | 62.2 | C. abortus 15-58d44 | 100 | MT423478 |
| 752K | Carrion crow | 253 | 51.0 | C. abortus 15-58d44 | 100 | MT423479 |
| 753C | Carrion crow | 274 | 75.9 | C. abortus 15-58d44 | 100 | MT423480 |
| 754C | Carrion crow | 267 | 82.8 | C. abortus 15-58d44 | 100 | MT423481 |
| 756C | Carrion crow | 278 | 75.5 | C. abortus 15-58d44 | 100 | MT423482 |
| 756K | Carrion crow | 240 | 28.3 | C. abortus 15-58d44 | 99.17 | MT423483 |
| 759C | Carrion crow | 278 | 72.7 | C. abortus 15-58d44 | 100 | MT423484 |
| 760C | Carrion crow | 279 | 68.1 | C. abortus 15-58d44 | 100 | MT423485 |
| 760K | Carrion crow | 250 | 53.2 | C. abortus 15-58d44 | 100 | MT423486 |
| 761C | Carrion crow | 256 | 81.3 | C. abortus 15-58d44 | 100 | MT423487 |
| 764C | Carrion crow | 276 | 71.7 | C. abortus 15-58d44 | 100 | MT423488 |
| 765C | Carrion crow | 278 | 78.8 | C. abortus 15-58d44 | 100 | MT423489 |
| 769C | Carrion crow | 276 | 81.2 | C. abortus 15-58d44 | 100 | MT423490 |
| 770C | Carrion crow | 256 | 75.4 | C. abortus 15-58d44 | 100 | MT423491 |
| 772C | Carrion crow | 267 | 82.8 | C. abortus 15-58d44 | 100 | MT423492 |
| 772K | Carrion crow | 270 | 80.4 | C. abortus 15-58d44 | 100 | MT423493 |
| 773C | Carrion crow | 269 | 76.6 | C. abortus 15-58d44 | 100 | MT423494 |
| 774C | Carrion crow | 277 | 75.1 | C. abortus 15-58d44 | 100 | MT423495 |
| 797C | Carrion crow | 275 | 77.1 | C. abortus 15-58d44 | 100 | MT423496 |
| 797K | Carrion crow | 265 | 64.2 | C. abortus 15-58d44 | 100 | MT423497 |
| 798C | Carrion crow | 277 | 75.8 | C. abortus 15-58d44 | 100 | MT423498 |
| 798K | Carrion crow | 271 | 81.2 | C. abortus 15-58d44 | 100 | MT423499 |
| 814C | Carrion crow | 266 | 82.7 | C. abortus 15-58d44 | 100 | MT423500 |
| 814K | Carrion crow | 278 | 74.8 | C. abortus 15-58d44 | 100 | MT423501 |
| 826C | Carrion crow | 266 | 77.8 | C. abortus 15-58d44 | 100 | MT423502 |
| 846C | Carrion crow | 276 | 71.7 | C. abortus 15-58d44 | 100 | MT423503 |
| 847C | Carrion crow | 238 | 43.3 | C. abortus 15-58d44 | 99.58 | MT423504 |
| 848C | Carrion crow | 267 | 82.8 | C. abortus 15-58d44 | 100 | MT423505 |
| 850C | Carrion crow | 256 | 75.4 | C. abortus 15-58d44 | 100 | MT423506 |
| 850K | Carrion crow | 278 | 65.5 | C. abortus 15-58d44 | 100 | MT423507 |
| 851C | Carrion crow | 264 | 82.6 | C. abortus 15-58d44 | 100 | MT423508 |
| 856C | Carrion crow | 267 | 80.1 | C. abortus 15-58d44 | 100 | MT423509 |
| 858C | Carrion crow | 255 | 74.9 | C. abortus 15-58d44 | 100 | MT423510 |
| 858K | Carrion crow | 271 | 77.1 | C. abortus 15-58d44 | 100 | MT423511 |
| 861C | Carrion crow | 276 | 80.1 | C. abortus 15-58d44 | 100 | MT423512 |
| 861K | Carrion crow | 270 | 40.0 | C. abortus 15-58d44 | 99.63 | MT423513 |
| 972C | Carrion crow | 260 | 83.8 | C. abortus 15-58d44 | 100 | MT423514 |
| Sample Nr. | Species Name (English) | Sequence Length (bp) | Sequence Quality (%) | First Hit | Nucleotide Identity (%) | Accession Number |
|---|---|---|---|---|---|---|
| Raptors | ||||||
| 268C | Eurasian sparrowhawk | 1000 | 86.1 | C. psittaci Ful127 | 97.99 | MT430892 |
| 311K | Common kestrel | 921 | 78.8 | C. psittaci M56 | 99.57 | MT429304 |
| 556K | Common buzzard | 1395 | 95.8 | C. psittaci M56 | 100 | MT430893 |
| 566C | Common buzzard | 1147 | 94.9 | C. psittaci nier_A113 | 100 | MT430895 |
| Corvids | ||||||
| 565C | Carrion crow | 996 | 98.4 | C. psittaci nier_A113 | 99.90 | MT430894 |
| 621C | Rook | 1357 | 91.4 | C. psittaci nier_A113 | 100 | MT430896 |
| 746C | Carrion crow | 1218 | 95.7 | C. psittaci nier_A113 | 99.92 | MT430897 |
| 769C | Carrion crow | 1370 | 95.5 | C. psittaci nier_A113 | 99.85 | MT430898 |
| 814C | Carrion crow | 1379 | 93.8 | C. psittaci nier_A97 | 99.93 | MT430899 |
| 972C | Carrion crow | 1071 | 93.8 | C. psittaci nier_A113 | 99.72 | MT430900 |
| Sample Nr. | Species Name (English) | Canton of Origin | Year of Sampling | Mean Cq Value Chlamydiaceae qPCR | OmpA Genotype | Accession Number |
|---|---|---|---|---|---|---|
| Raptors | ||||||
| 268C | Eurasian sparrowhawk | Unknown | 2018 | 26.3 | A | MT450242 |
| 311K | Common kestrel | Unknown | 2018 | 26.8 | M56 | MT450243 |
| 511C | Common buzzard | Unknown | 2019 | 38.3 | M56 | MT450244 |
| 556K | Common buzzard | Zurich | 2019 | 14.1 | M56 | MT450245 |
| 566C | Common buzzard | Obwalden | 2019 | 29.8 | 1V | MT450247 |
| 669C | Common buzzard | Unknown | 2019 | 27.9 | M56 | MT450249 |
| 671K | Common buzzard | Thurgau | 2019 | 27.1 | M56 | MT450250 |
| 683C | Common buzzard | Unknown | 2019 | 33.3 | A | MT450251 |
| 877K | Long-eared owl | Bern | 2019 | 31.3 | M56 | MT450275 |
| Corvids | ||||||
| 565C | Carrion crow | Aargau | 2019 | 27.9 | 1V | MT450246 |
| 621C | Rook | Unknown | 2019 | 23.9 | 1V | MT450248 |
| 688C | Carrion crow | Zurich | 2019 | 28.2 | 1V | MT450252 |
| 689C | Carrion crow | Zurich | 2019 | 26.8 | 1V | MT450253 |
| 690C | Carrion crow | Zurich | 2019 | 29.4 | 1V | MT450254 |
| 706C | Carrion crow | Zurich | 2019 | 30.2 | D | MT450255 |
| 711C | Carrion crow | Bern | 2019 | 31.6 | 1V | MT450256 |
| 716K | Carrion crow | Bern | 2019 | 30.6 | D | MT450257 |
| 721C | Carrion crow | Zurich | 2019 | 29.9 | 1V | MT450258 |
| 735C | Carrion crow | Zug | 2019 | 28.4 | 1V | MT450259 |
| 736C | Carrion crow | Zug | 2019 | 28.7 | 6N | MT450260 |
| 740C | Carrion crow | Zug | 2019 | 27.7 | 1V | MT450261 |
| 746C | Carrion crow | Zug | 2019 | 24.9 | 1V | MT450262 |
| 751C | Carrion crow | Zug | 2019 | 20.2 | 1V | MT450263 |
| 752C | Carrion crow | Zug | 2019 | 30.4 | 1V | MT450264 |
| 761C | Carrion crow | Zug | 2019 | 29.7 | 1V | MT450265 |
| 772K | Carrion crow | Zug | 2019 | 26.8 | 1V | MT450266 |
| 798K | Carrion crow | Zurich | 2019 | 26.7 | 1V | MT450267 |
| 814C | Carrion crow | Zurich | 2019 | 25.6 | 1V | MT450268 |
| 826C | Carrion crow | Zurich | 2019 | 29.3 | 1V | MT450269 |
| 848C | Carrion crow | Zurich | 2019 | 26.6 | D | MT450270 |
| 850K | Carrion crow | Zurich | 2019 | 27.9 | 1V | MT450271 |
| 856C | Carrion crow | Zurich | 2019 | 29.3 | 1V | MT450272 |
| 858C | Carrion crow | Zurich | 2019 | 27.8 | 1V | MT450273 |
| 861K | Carrion crow | Unknown | 2019 | 23.6 | 1V | MT450274 |
| 972C | Carrion crow | Fribourg | 2019 | 27.4 | 1V | MT450276 |
| Order | Family | Species Name (Latin) | Species Name (English) | Number of Birds | Number of Choanal Swabs | Number of Cloacal Swabs | Number of Fecal Swabs |
|---|---|---|---|---|---|---|---|
| Accipitriformes | Accipitridae | Gypaetus barbatus | Bearded vulture | 1 | 1 | 1 | 0 |
| Milvus migrans | Black kite | 6 | 4 | 4 | 3 | ||
| Buteo buteo | Common buzzard | 142 | 127 | 128 | 14 | ||
| Accipiter nisus | Eurasian sparrowhawk | 32 | 32 | 32 | 1 | ||
| Pernis apivorus | European honey-buzzard | 1 | 1 | 1 | 0 | ||
| Aquila chrysaetos | Golden eagle | 6 | 3 | 6 | 0 | ||
| Circus pygargus | Montagu’s harrier | 1 | 1 | 1 | 0 | ||
| Accipiter gentilis | Northern goshawk | 2 | 2 | 2 | 0 | ||
| Milvus milvus | Red kite | 23 | 20 | 21 | 2 | ||
| Falconiformes | Falconidae | Falco tinnunculus | Common kestrel | 66 | 54 | 55 | 12 |
| Falco subbuteo | Eurasian hobby | 4 | 3 | 3 | 1 | ||
| Falco peregrinus | Peregrine falcon | 1 | 1 | 1 | 0 | ||
| Passeriformes | Corvidae | Corvus corone | Carrion crow | 207 | 190 | 187 | 19 |
| Garrulus glandarius | Eurasian jay | 9 | 9 | 8 | 1 | ||
| Pica pica | Eurasian magpie | 30 | 16 | 16 | 22 | ||
| Corvus cornix | Hooded crow | 1 | 1 | 1 | 0 | ||
| Corvus frugilegus | Rook | 3 | 3 | 3 | 2 | ||
| Corvus monedula | Western jackdaw | 3 | 1 | 1 | 3 | ||
| Strigiformes | Strigidae | Bubo bubo | Eurasian eagle-owl | 4 | 4 | 4 | 0 |
| Asio otus | Long-eared owl | 17 | 15 | 14 | 2 | ||
| Strix aluco | Tawny owl | 18 | 14 | 14 | 6 | ||
| Tytonidae | Tyto alba | Barn owl | 17 | 17 | 17 | 1 | |
| Total | 594 | 519 | 520 | 89 |
| Method | Target | Final Concentration | Primer & Probe | Sequence (5′–3′) | Amplicon Size (Base Pairs) | Annealing Temperature (°C) | References |
|---|---|---|---|---|---|---|---|
| Chlamydiaceae 23S rRNA qPCR | 23S rRNA | 500 nM | Ch23S-F Ch23S-R | CTGAAACCAGTAGCTTATAAGCGGT ACCTCGCCGTTTAACTTAACTCC | 111 | 60 | Ehricht et al. (2006) [64] |
| 200 nM | Ch23S-P | FAM-CTCATCATGCAAAAGGCACGCCG-TAMRA | |||||
| Internal amplification control | eGFP | 200 nM | eGFP-1-F | GACCACTACCAGCAGAACAC | 177 | Hoffmann et al. (2006) [65] PCR modified by Blumer et al. (2011) [66] | |
| eGFP-10-R | CTTGTACAGCTCGTCCATGC | ||||||
| eGFP-HEX | HEX-AGCACCCAGTCCGCCCTGAGCA-BHQ1 | ||||||
| C. psittaci ompA qPCR | ompA | 900 nM | CppsOMP1-F | CACTATGTGGGAAGGTGCTTCA | 76 | 60 | Pantchev et al. (2009) [67] |
| CppsOMP1-R | CTGCGCGGATGCTAATGG | ||||||
| 200 nM | CppsOMP1-S | FAM-CGCTACTTGGTGTGAC-TAMRA | |||||
| Internal amplification control | eGFP | 900 nM | eGFP-1-F | GACCACTACCAGCAGAACAC | 132 | Hoffmann et al. (2005) [68] | |
| eGFP-2-R | GAACTCCAGCAGGACCATG | ||||||
| 200 nM | eGFP-HEX | HEX-AGCACCCAGTCCGCCCTGAGCA-BHQ1 | |||||
| 16S rRNA PCR (partial) | 16S rRNA | 300 nM | 16S IGF (short) 16S IGR (short) | GATGAGGCATGCAAGTCGAACG CCAGTGTTGGCGGTCAATCTCTC | 278 | 65 | Blumer et al. (2007) [70], Modified from Everett et al. (1999) [75] |
| 16S rRNA PCR (near-full length) | 16S rRNA | 300 nM | 16S-IGF 16S-B1 | CGGCGTGGATGAGGCAT TACGGYTACCTTGTTACGACTT | 1481 | 57.5 | Everett et al. (1999) [69] Hosokawa et al. (2006) [72] |
| C. buteonis oxaA qPCR | oxaA | 600 nM | RSHA-F | ATTTCCAACACGCACTGCAT | 80 | 60 | Laroucau et al. (2019) [3] |
| RSHA-R | TGGGACTAGGTGTTCTCCCT | ||||||
| 200 nM | RSHA-P | FAM-GGACAACATGCCTAGATGAAGA-TAMRA | |||||
| Internal amplification control | eGFP | 400 nM | eGFP-1-F | GACCACTACCAGCAGAACAC | 132 | Hoffmann et al. (2005) [68] | |
| eGFP-2-R | GAACTCCAGCAGGACCATG | ||||||
| 200 nM | eGFP-HEX | HEX-AGCACCCAGTCCGCCCTGAGCA-BHQ1 | |||||
| ompA typing PCR | ompA | 200 nM | ompA F (CTU) | ATGAAAAAACTCTTGAAATCGG | 1212 | 49 | Sachse et al. (2008) [73] |
| ompA rev | TCCTTAGAATCTGAATTGAGC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stalder, S.; Marti, H.; Borel, N.; Sachse, K.; Albini, S.; Vogler, B.R. Occurrence of Chlamydiaceae in Raptors and Crows in Switzerland. Pathogens 2020, 9, 724. https://doi.org/10.3390/pathogens9090724
Stalder S, Marti H, Borel N, Sachse K, Albini S, Vogler BR. Occurrence of Chlamydiaceae in Raptors and Crows in Switzerland. Pathogens. 2020; 9(9):724. https://doi.org/10.3390/pathogens9090724
Chicago/Turabian StyleStalder, Sandro, Hanna Marti, Nicole Borel, Konrad Sachse, Sarah Albini, and Barbara Renate Vogler. 2020. "Occurrence of Chlamydiaceae in Raptors and Crows in Switzerland" Pathogens 9, no. 9: 724. https://doi.org/10.3390/pathogens9090724
APA StyleStalder, S., Marti, H., Borel, N., Sachse, K., Albini, S., & Vogler, B. R. (2020). Occurrence of Chlamydiaceae in Raptors and Crows in Switzerland. Pathogens, 9(9), 724. https://doi.org/10.3390/pathogens9090724