Inhibition of Asaia in Adult Mosquitoes Causes Male-Specific Mortality and Diverse Transcriptome Changes

 ,
,  ,
, 
 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
3. Materials and Methods
3.1. An. stephensi Rearing
3.2. Production of Monoclonal anti-Asaia IgG
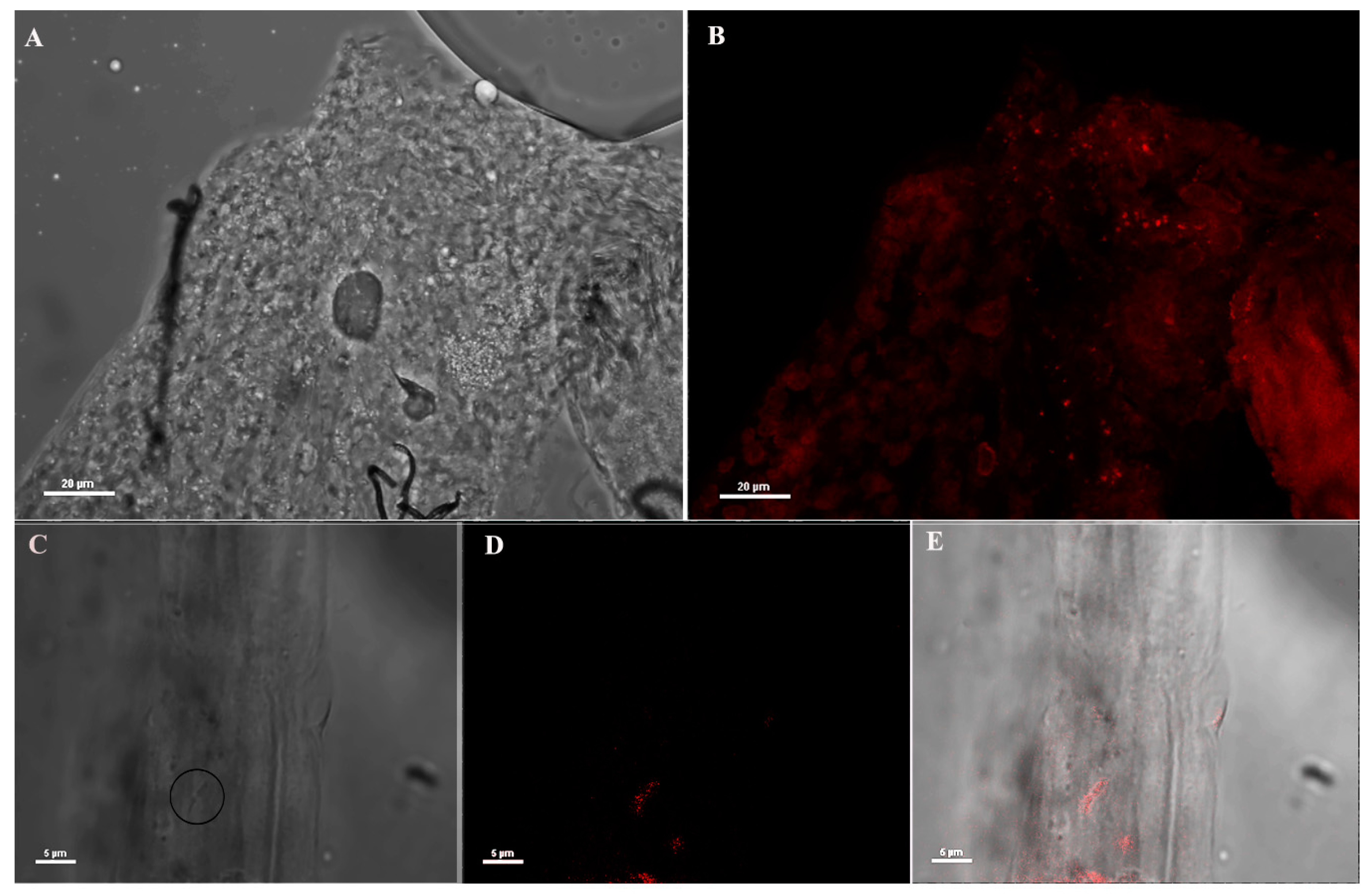
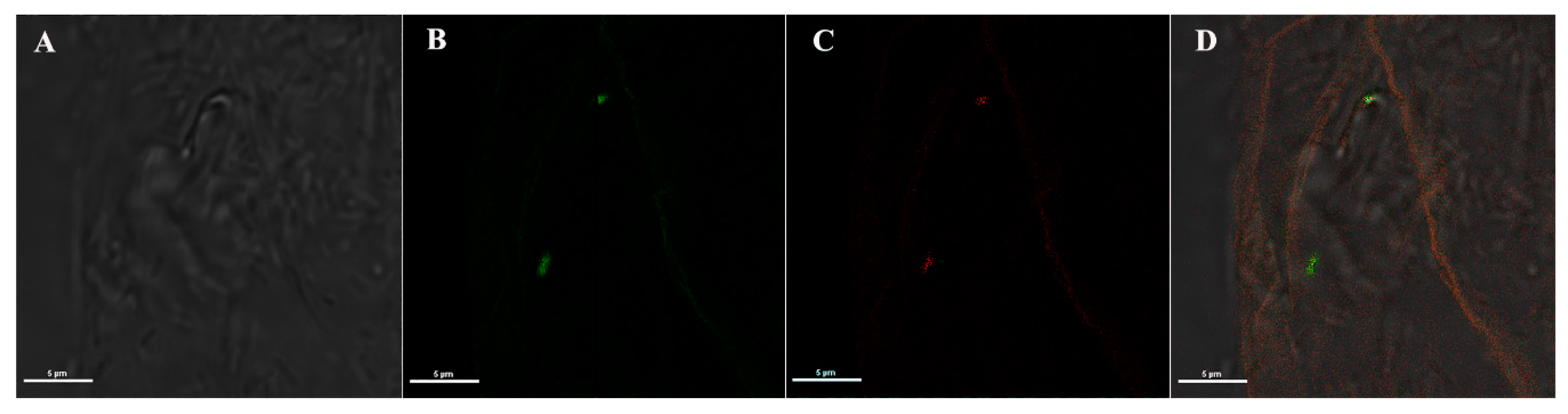
3.3. Immuno Fluorescence Assay (IFA) on An. stephensi Mosquitoes Microinjected with Anti-Asaia Antibody
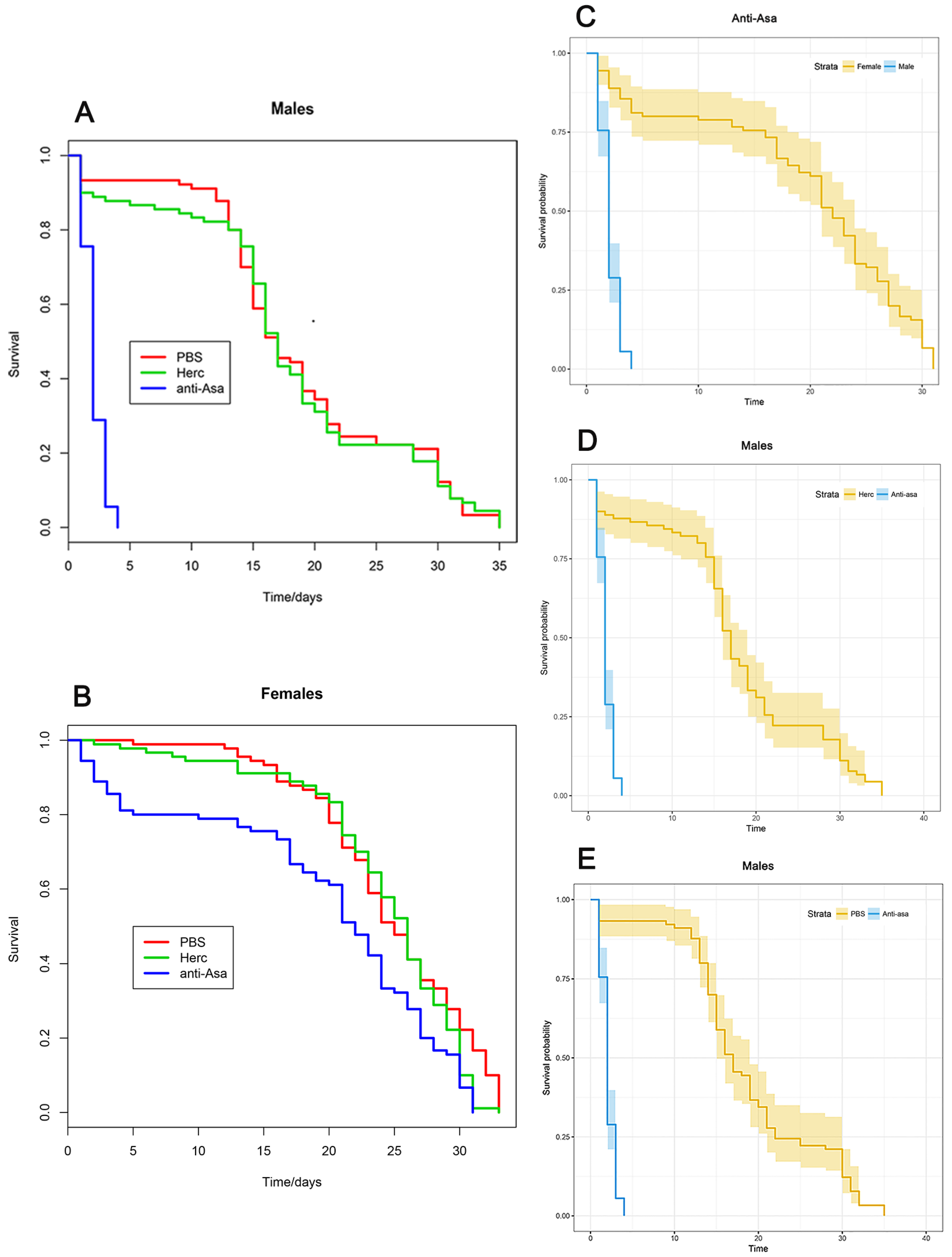
3.4. Mosquito Survival Test
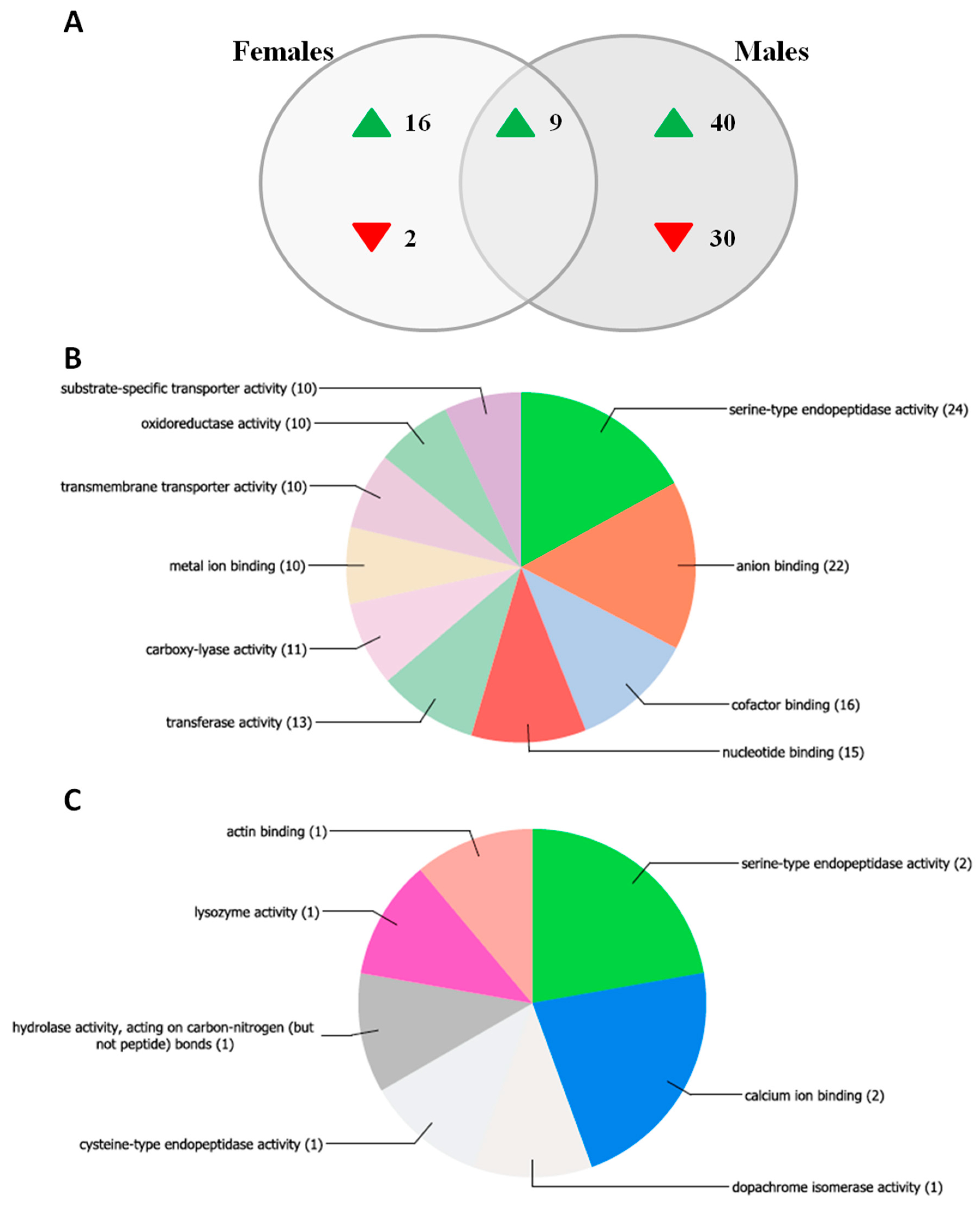
3.5. Mosquito Transcriptome Analysis by Microarray Assay
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Norris, E.J.; Coats, J.R. Current and Future Repellent Technologies: The Potential of Spatial Repellents and Their Place in Mosquito-Borne Disease Control. Int. J. Environ. Res. Public Health 2017, 14, 124. [Google Scholar] [CrossRef]
- Riehle, M.A.; Jacobs-Lorena, M. Using bacteria to express and display anti-parasite molecules in mosquitoes: Current and future strategies. Insect Biochem. Mol. Biol. 2005, 35, 699–707. [Google Scholar] [CrossRef]
- Ricci, I.; Valzano, M.; Ulissi, U.; Epis, S.; Cappelli, A.; Favia, G. Symbiotic control of mosquito borne disease. Pathog. Glob. Health 2012, 106, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Ricci, I.; Damiani, C.; Capone, A.; DeFreece, C.; Rossi, P.; Favia, G. Mosquito/microbiota interactions: From complex relationships to biotechnological perspectives. Curr. Opin. Microbiol. 2012, 15, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Mancini, M.V.; Spaccapelo, R.; Damiani, C.; Accoti, A.; Tallarita, M.; Petraglia, E.; Rossi, P.; Cappelli, A.; Capone, A.; Peruzzi, G.; et al. Paratransgenesis to control malaria vectors: A semi-field pilot study. Parasit. Vectors 2016, 9, 140. [Google Scholar] [CrossRef] [PubMed]
- Bongio, N.J.; Lampe, D.J. Inhibition of Plasmodium berghei Development in Mosquitoes by Effector Proteins Secreted from Asaia sp. Bacteria Using a Novel Native Secretion Signal. PLoS ONE 2015, 10, e0143541. [Google Scholar] [CrossRef]
- Cappelli, A.; Damiani, C.; Mancini, M.V.; Valzano, M.; Rossi, P.; Serrao, A.; Ricci, I.; Favia, G. Asaia Activates Immune Genes in Mosquito Eliciting an Anti-Plasmodium Response: Implications in Malaria Control. Front. Genet. 2019, 10, 836. [Google Scholar] [CrossRef]
- Gonella, E.; Mandrioli, M.; Tedeschi, R.; Crotti, E.; Pontini, M.; Alma, A. Activation of Immune Genes in Leafhoppers by Phytoplasmas and Symbiotic Bacteria. Front. Physiol. 2019, 10, 795. [Google Scholar] [CrossRef]
- Chouaia, B.; Rossi, P.; Epis, S.; Mosca, M.; Ricci, I.; Damiani, C.; Ulissi, U.; Crotti, E.; Daffonchio, D.; Bandi, C.; et al. Delayed larval development in Anopheles mosquitoes deprived of Asaia bacterial symbionts. BMC Microbiol. 2012, 12, S2. [Google Scholar] [CrossRef]
- Mitraka, E.; Stathopoulos, S.; Siden-Kiamos, I.; Christophides, G.K.; Louis, C. Asaia accelerates larval development of Anopheles gambiae. Pathog. Glob. Health 2013, 107, 305–311. [Google Scholar] [CrossRef]
- Forthal, D.N. Functions of Antibodies. Antib. Infect. Dis. 2015, 23–48. [Google Scholar] [CrossRef]
- Dinparast Djadid, N.; Jazayeri, H.; Raz, A.; Favia, G.; Ricci, I.; Zakeri, S. Identification of the midgut microbiota of An. stephensi and An. maculipennis for their application as a paratransgenic tool against malaria. PLoS ONE 2011, 6, e28484. [Google Scholar] [CrossRef] [PubMed]
- Kanost, M.R.; Jiang, H. Clip-domain serine proteases as immune factors in insect hemolymph. Curr. Opin. Insect Sci. 2015, 11, 47–55. [Google Scholar] [CrossRef]
- Yassine, H.; Kamareddine, L.; Chamat, S.; Christophides, G.K.; Osta, M.A. A Serine Protease Homolog Negatively Regulates TEP1 Consumption in Systemic Infections of the Malaria Vector Anopheles gambiae. J. Innate Immun. 2014, 6, 806–818. [Google Scholar] [CrossRef]
- Povelones, M.; Bhagavatula, L.; Yassine, H.; Tan, L.A.; Upton, L.M.; Osta, M.A.; Christophides, G.K. The CLIP-Domain Serine Protease Homolog SPCLIP1 Regulates Complement Recruitment to Microbial Surfaces in the Malaria Mosquito Anopheles gambiae. PLoS Pathog. 2013, 9, e1003623. [Google Scholar] [CrossRef]
- Gendrin, M.; Turlure, F.; Rodgers, F.H.; Cohuet, A.; Morlais, I.; Christophides, G.K. The Peptidoglycan Recognition Proteins PGRPLA and PGRPLB Regulate Anopheles Immunity to Bacteria and Affect Infection by Plasmodium. J. Innate Immun. 2017, 9, 333–342. [Google Scholar] [CrossRef]
- Sierralta, J.; Mendoza, C. PDZ-containing proteins: Alternative splicing as a source of functional diversity. Brain Res. Rev. 2004, 47, 105–115. [Google Scholar] [CrossRef]
- Vogel, H.; Shukla, S.P.; Engl, T.; Weiss, B.; Fischer, R.; Steiger, S.; Heckel, D.G.; Kaltenpoth, M.; Vilcinskas, A. The digestive and defensive basis of carcass utilization by the burying beetle and its microbiota. Nat. Commun. 2017, 8, 15186. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, J.S.; MacDonald, S.J.; Jander, G.; Nakabachi, A.; Thomas, G.H.; Douglas, A.E. Genomic evidence for complementary purine metabolism in the pea aphid, Acyrthosiphon pisum, and its symbiotic bacterium Buchnera aphidicola. Insect Mol. Biol. 2010, 19, 241–248. [Google Scholar] [CrossRef]
- Kim, J.K.; Kwon, J.Y.; Kim, S.K.; Han, S.H.; Won, Y.J.; Lee, J.H.; Kim, C.H.; Fukatsu, T.; Lee, B. Purine biosynthesis, biofilm formation, and persistence of an insect-microbe gut symbiosis. Appl. Environ. Microbiol. 2014, 80, 4374–4382. [Google Scholar] [CrossRef]
- Kim, J.K.; Jang, H.A.; Won, Y.J.; Kikuchi, Y.; Han, S.H.; Kim, C.H.; Nikoh, N.; Fukatsu, T.; Lee, B.L. Purine biosynthesis-deficient Burkholderia mutants are incapable of symbiotic accommodation in the stinkbug. ISME J. 2014, 8, 552–563. [Google Scholar] [CrossRef] [PubMed]
- Pondeville, E.; David, J.P.; Guittard, E.; Maria, A.; Jacques, J.C.; Ranson, H.; Bourgouin, C.; Dauphin-Villemant, C. Microarray and RNAi analysis of P450s in Anopheles gambiae male and female steroidogenic tissues: CYP307A1 is required for ecdysteroid synthesis. PLoS ONE 2013, 8, e79861. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Wang, Y.; Li, F.; Liu, J.; Sun, Y.; Wu, J. Cloning and characterization of a mannose binding C-type lectin gene from salivary gland of Aedes albopictus. Parasit. Vectors 2014, 7, 337. [Google Scholar] [CrossRef] [PubMed]
- Cohuet, A.; Osta, M.A.; Morlais, I.; Awono-Ambene, P.H.; Michel, K.; Simard, F.; Christophides, G.K.; Fontenille, D.; Kafatos, F.C. Anopheles and Plasmodium: From laboratory models to natural systems in the field. EMBO Rep. 2006, 7, 1285–1289. [Google Scholar] [CrossRef] [PubMed]
- Danielli, A.; Kafatos, F.C.; Loukeris, T.G. Cloning and characterization of four Anopheles gambiae serpin isoforms, differentially induced in the midgut by Plasmodium berghei invasion. J. Biol. Chem. 2003, 278, 4184–4193. [Google Scholar] [CrossRef]
- Köhler, G.; Milstein, C. Continuous cultures of fused cells secreting antibody of predefined specificity. Nature 1975, 256, 495–497. [Google Scholar] [CrossRef]
- Kaplan, E.L.; Meier, P. Nonparametric estimation from incomplete observations. J. Am. Statist. Assn. 1958, 53, 457–481. [Google Scholar] [CrossRef]
- Pike, A.; Vadlamani, A.; Sandiford, S.L.; Gacita, A.; Dimopoulos, G. Characterization of the Rel2-regulated transcriptome and proteome of Anopheles stephensi identifies new anti-Plasmodium factors. Insect Biochem. Mol. Biol. 2014, 52, 82–93. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Short, S.M.; Mongodin, E.F.; MacLeod, H.J.; Talyuli, O.A.C.; Dimopoulos, G. Amino acid metabolic signaling influences Aedes aegypti midgut microbiome variability. PLoS Negl. Trop. Dis. 2017, 11, e0005677. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Silver, J.; Oshlack, A.; Holmes, M.; Diyagama, D.; Holloway, A.; Smyth, G.K. A comparison of background correction methods for two-colour microarrays. Bioinformatics 2007, 23, 2700–2707. [Google Scholar] [CrossRef] [PubMed]
- Smyth, G.K.; Speed, T. Normalization of cDNA microarray data. Methods 2003, 31, 265–273. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mancini, M.V.; Damiani, C.; Short, S.M.; Cappelli, A.; Ulissi, U.; Capone, A.; Serrao, A.; Rossi, P.; Amici, A.; Kalogris, C.; et al. Inhibition of Asaia in Adult Mosquitoes Causes Male-Specific Mortality and Diverse Transcriptome Changes. Pathogens 2020, 9, 380. https://doi.org/10.3390/pathogens9050380
Mancini MV, Damiani C, Short SM, Cappelli A, Ulissi U, Capone A, Serrao A, Rossi P, Amici A, Kalogris C, et al. Inhibition of Asaia in Adult Mosquitoes Causes Male-Specific Mortality and Diverse Transcriptome Changes. Pathogens. 2020; 9(5):380. https://doi.org/10.3390/pathogens9050380
Chicago/Turabian StyleMancini, Maria Vittoria, Claudia Damiani, Sarah M. Short, Alessia Cappelli, Ulisse Ulissi, Aida Capone, Aurelio Serrao, Paolo Rossi, Augusto Amici, Cristina Kalogris, and et al. 2020. "Inhibition of Asaia in Adult Mosquitoes Causes Male-Specific Mortality and Diverse Transcriptome Changes" Pathogens 9, no. 5: 380. https://doi.org/10.3390/pathogens9050380
APA StyleMancini, M. V., Damiani, C., Short, S. M., Cappelli, A., Ulissi, U., Capone, A., Serrao, A., Rossi, P., Amici, A., Kalogris, C., Dimopoulos, G., Ricci, I., & Favia, G. (2020). Inhibition of Asaia in Adult Mosquitoes Causes Male-Specific Mortality and Diverse Transcriptome Changes. Pathogens, 9(5), 380. https://doi.org/10.3390/pathogens9050380