A Retrospective Assessment of Temperature Trends in Northern Europe Reveals a Deep Impact on the Life Cycle of Ixodes ricinus (Acari: Ixodidae)


Abstract
1. Introduction
2. Results
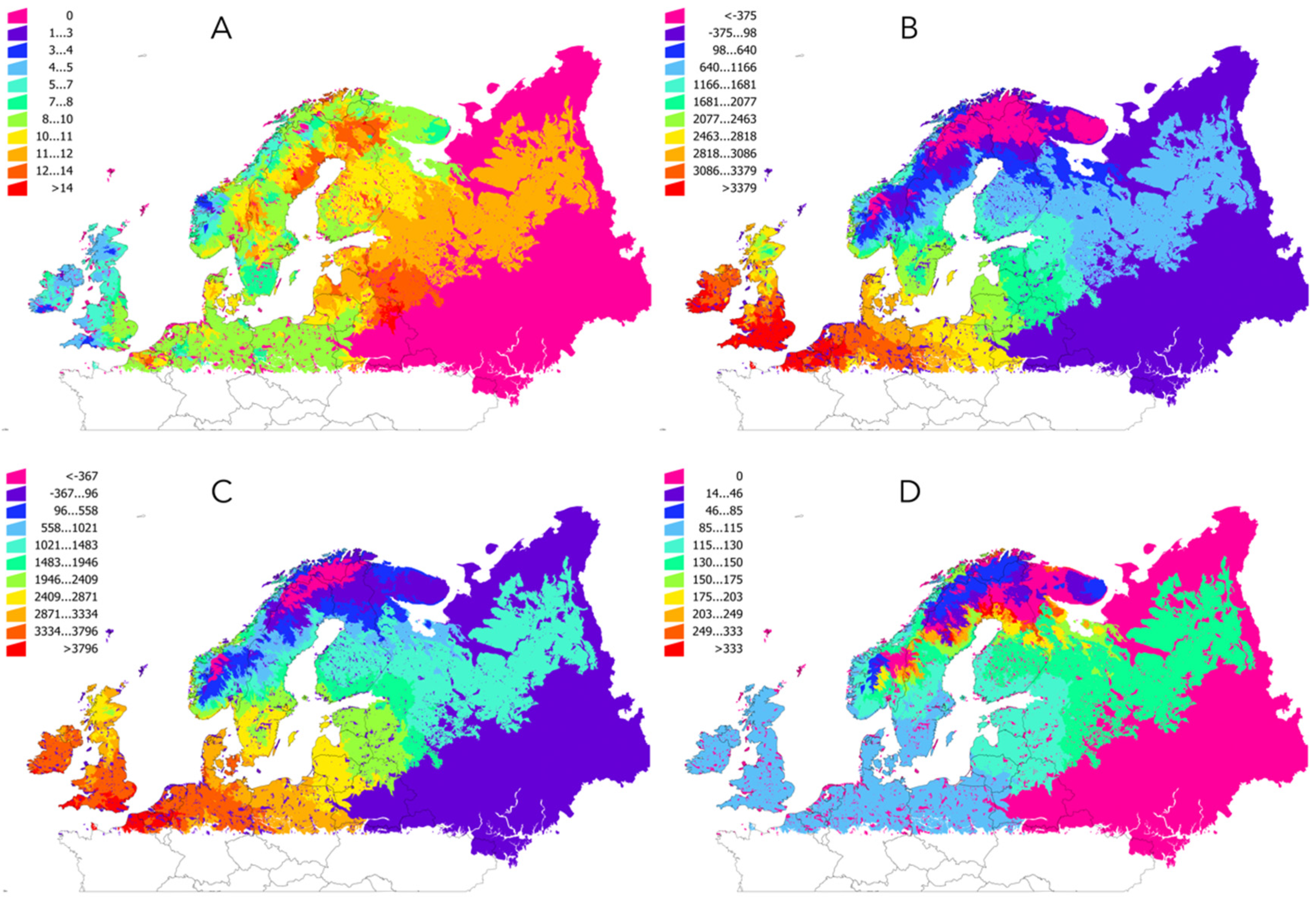
2.1. Temperature Increased in the Target Region in the Period 1950–2018
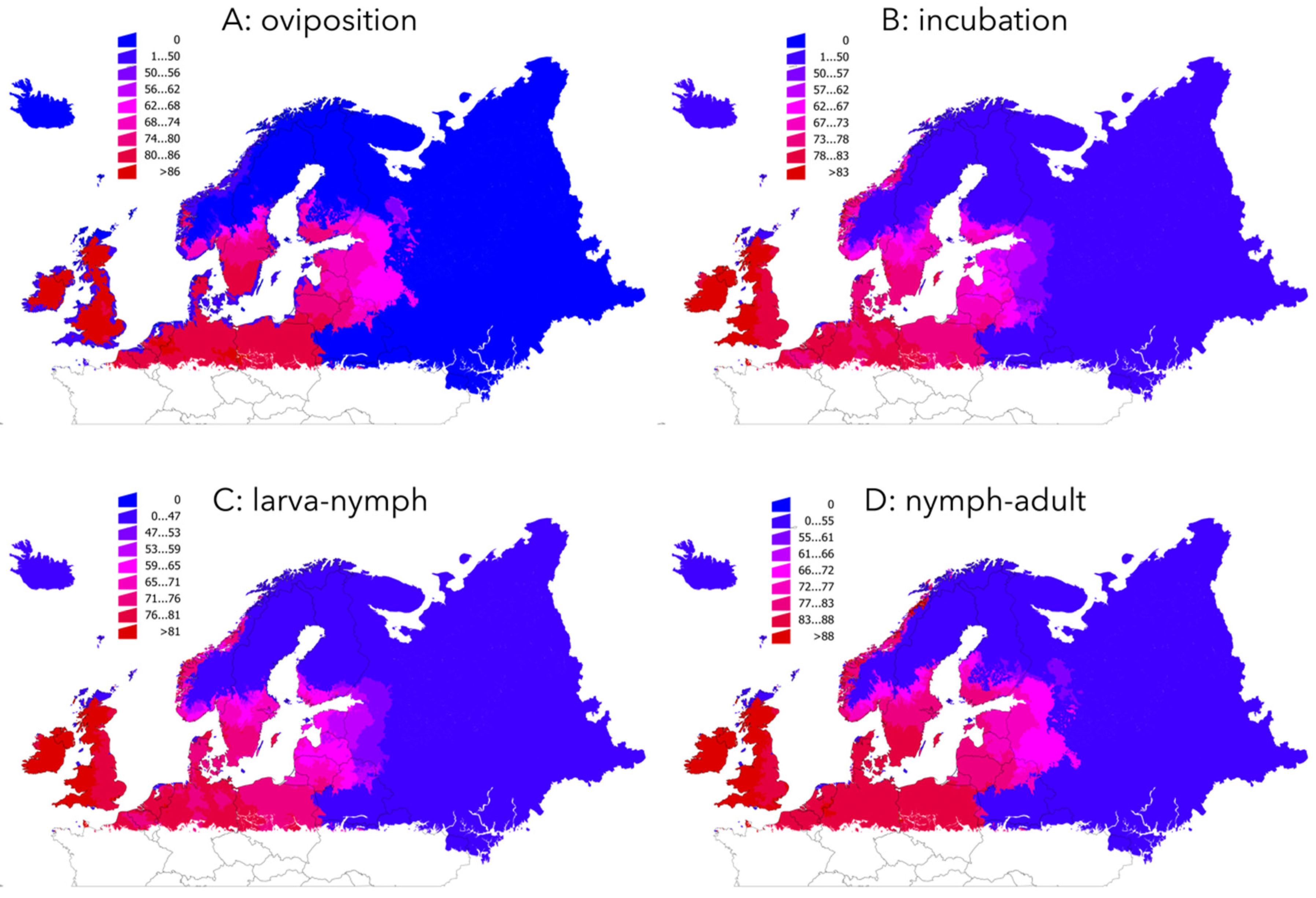
2.2. The Development Rates of I. ricinus Life Cycle are Faster Mainly in Northern Countries
3. Discussion
4. Material and Methods
4.1. Purpose
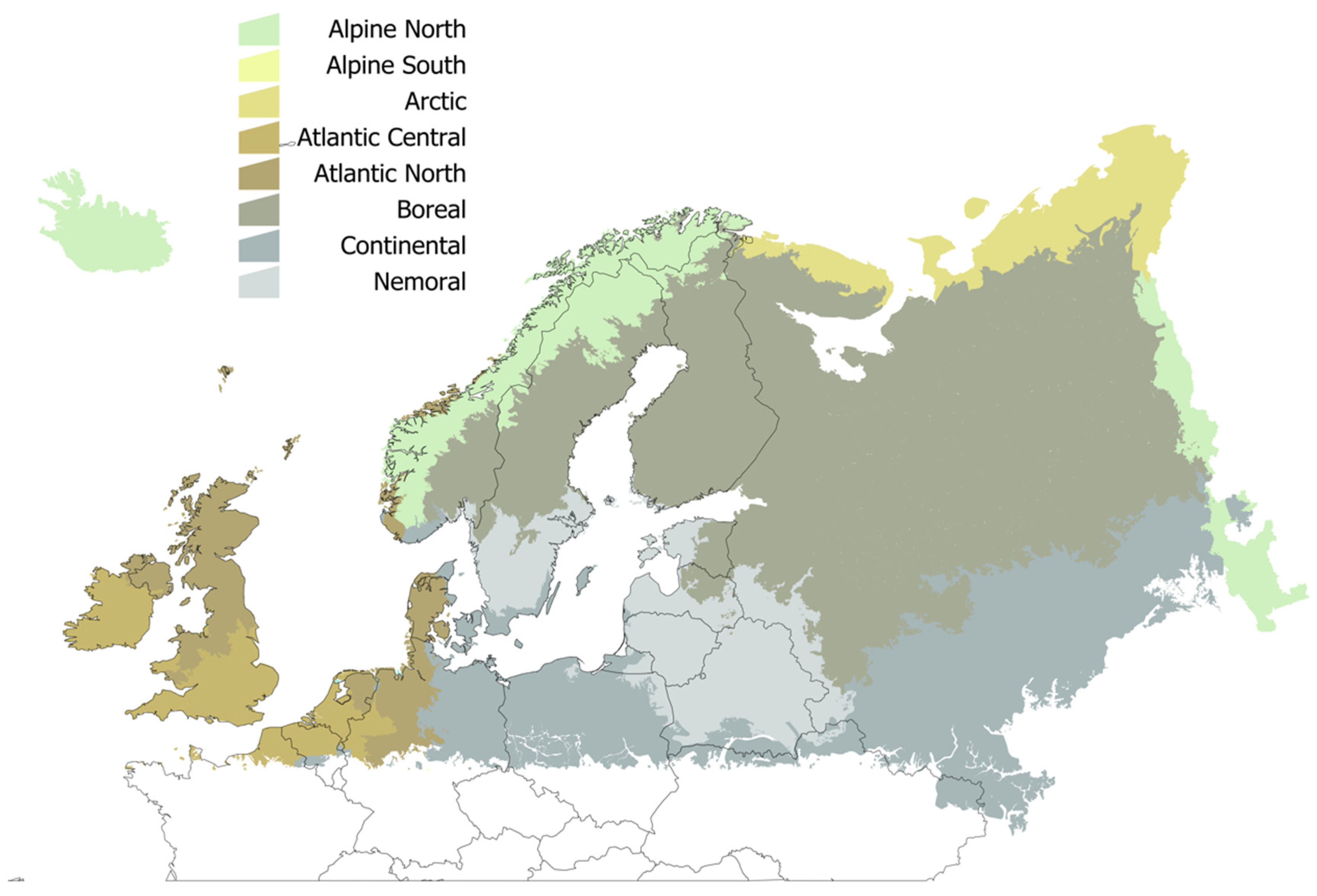
4.2. Data on Climate and Ecological Regions in Europe
4.3. Calculation of the Development of Ixodes ricinus
4.4. Other Calculations
5. Conclusions and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Ripple:, W.J.; Wolf, C.; Newsome, T.M.; Galetti, M.; Alamgir, M.; Crist, E.; 15,364 Scientist Signatories from 184 Countries. World scientists’ warning to humanity: A second notice. BioScience 2017, 67, 1026–1028. [Google Scholar]
- Araújo, M.B.; Pearson, R.G.; Thuiller, W.; Erhard, M. Validation of species-climate impact models under climate change. Glob. Chang. Biol. 2005, 11, 1504–1513. [Google Scholar] [CrossRef]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37. [Google Scholar] [CrossRef] [PubMed]
- Pearson, R.G.; Dawson, T.P. Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful? Glob. Ecol. Biogeogr. 2003, 12, 361–371. [Google Scholar] [CrossRef]
- Hales, S.; De Wet, N.; Maindonald, J.; Woodward, A. Potential effect of population and climate changes on global distribution of dengue fever: An empirical model. Lancet 2002, 360, 830–834. [Google Scholar] [CrossRef]
- Reiter, P. Climate change and mosquito-borne disease. Environ. Health Perspect. 2001, 109 (Suppl. 1), 141. [Google Scholar]
- Lambrechts, L.; Paaijmans, K.P.; Fansiri, T.; Carrington, L.B.; Kramer, L.D.; Thomas, M.B.; Scott, T.W. Impact of daily temperature fluctuations on dengue virus transmission by Aedes aegypti. Proc. Natl. Acad. Sci. USA 2011, 108, 7460–7465. [Google Scholar] [CrossRef]
- Rochlin, I.; Ninivaggi, D.V.; Hutchinson, M.L.; Farajollahi, A. Climate change and range expansion of the Asian tiger mosquito (Aedes albopictus) in Northeastern USA: Implications for public health practitioners. PLoS ONE 2013, 8, e60874. [Google Scholar] [CrossRef]
- Hanincová, K.; Kurtenbach, K.; Diuk-Wasser, M.; Brei, B.; Fish, D. Epidemic spread of Lyme borreliosis, northeastern United States. Emerg. Inf. Dis. 2006, 12, 604. [Google Scholar] [CrossRef]
- Kimberlin, D.W.; Brady, M.T.; Jackson, M.A.; Long, S.S. Red Book 2015 Report of the Committee on Infectious Diseases; American Academy of Pediatrics: Washington, DC, USA, 2015. [Google Scholar]
- Leblebicioglu, H.; Ozaras, R.; Irmak, H.; Sencan, I. Crimean-Congo hemorrhagic fever in Turkey: Current status and future challenges. Antivir. Res. 2016, 126, 21–34. [Google Scholar] [CrossRef]
- Al-Abri, S.S.; Al Abaidani, I.; Fazlalipour, M.; Mostafavi, E.; Leblebicioglu, H.; Pshenichnaya, N.; Nguyen, T.M.N. Current status of Crimean-Congo haemorrhagic fever in the World Health Organization Eastern Mediterranean Region: Issues, challenges, and future directions. Int. J. Infect. Dis. 2017, 58, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Vial, L.; Stachurski, F.; Leblond, A.; Huber, K.; Vourc’h, G.; René-Martellet, M.; Desjardins, I.; Balança, G.; Grosbois, V.; Pradier, S.; et al. Strong evidence for the presence of the tick Hyalomma marginatum Koch, 1844 in southern continental France. Ticks Tick-Borne Dis. 2016, 7, 1162–1167. [Google Scholar] [CrossRef] [PubMed]
- Dautel, H.; Kämmer, D.; Kahl, O. How an extreme weather spell in winter can influence vector tick abundance and tick-borne disease incidence. In Ecology and Prevention of Lyme Borreliosis; Wageningen Academic Publishers: Wageningen, The Netherlands, 2016; p. 362. [Google Scholar]
- Furness, R.W.; Furness, E.N. Ixodes ricinus parasitism of birds increases at higher winter temperatures. J. Vector Ecol. 2017, 43, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Randolph, S.E. Evidence that climate change has caused ‘emergence’ of tick-borne diseases in Europe? Int. J. Med. Microbiol. Suppl. 2004, 293, 5–15. [Google Scholar] [CrossRef]
- Sumilo, D.; Asokliene, L.; Bormane, A.; Vasilenko, V.; Golovljova, I.; Randolph, S.E. Climate change cannot explain the upsurge of tick-borne encephalitis in the Baltics. PLoS ONE 2007, 2, e500. [Google Scholar] [CrossRef] [PubMed]
- Leighton, P.A.; Koffi, J.K.; Pelcat, Y.; Lindsay, L.R.; Ogden, N.H. Predicting the speed of tick invasion: An empirical model of range expansion for the Lyme disease vector Ixodes scapularis in Canada. J. Appl. Ecol. 2012, 49, 457–464. [Google Scholar] [CrossRef]
- Ogden, N.H.; Maarouf, A.; Barker, I.K.; Bigras-Poulin, M.; Lindsay, L.R.; Morshed, M.G.; Charron, D.F. Climate change and the potential for range expansion of the Lyme disease vector Ixodes scapularis in Canada. Int. J. Parasitol. 2006, 36, 63–70. [Google Scholar] [CrossRef]
- Ogden, N.H.; St-Onge, L.; Barker, I.K.; Brazeau, S.; Bigras-Poulin, M.; Charron, D.F.; Michel, P. Risk maps for range expansion of the Lyme disease vector, Ixodes scapularis, in Canada now and with climate change. Int. J. Health Geogr. 2008, 7, 24. [Google Scholar] [CrossRef]
- Danielová, V.; Rudenko, N.; Daniel, M.; Holubová, J.; Materna, J.; Golovchenko, M.; Schwarzová, L. Extension of Ixodes ricinus ticks and agents of tick-borne diseases to mountain areas in the Czech Republic. Int. J. Med. Microbiol. 2006, 296, 48–53. [Google Scholar] [CrossRef]
- Jaenson, T.G.; Lindgren, E. The range of Ixodes ricinus and the risk of contracting Lyme borreliosis will increase northwards when the vegetation period becomes longer. Ticks Tick-Borne Dis. 2011, 2, 44–49. [Google Scholar] [CrossRef]
- Jaenson, T.G.; Eisen, L.; Comstedt, P.; Mejlon, H.A.; Lindgren, E.; Bergström, S.; Olsen, B. Risk indicators for the tick Ixodes ricinus and Borrelia burgdorferi sensu lato in Sweden. Med. Vet. Entomol. 2009, 23, 226–237. [Google Scholar] [CrossRef] [PubMed]
- Materna, J.; Daniel, M.; Danielová, V. Altitudinal distribution limit of the tick Ixodes ricinus shifted considerably towards higher altitudes in central Europe: Results of three years monitoring in the Krkonose Mts. (Czech Republic). Cent. Eur. J. Public Health 2005, 13, 24–28. [Google Scholar] [PubMed]
- Jore, S.; Viljugrein, H.; Hofshagen, M.; Brun-Hansen, H.; Kristoffersen, A.B.; Nygård, K.; Ytrehus, B. Multi-source analysis reveals latitudinal and altitudinal shifts in range of Ixodes ricinus at its northern distribution limit. Parasites Vectors 2011, 4, 84. [Google Scholar] [CrossRef] [PubMed]
- Tagliapietra, V.; Rosà, R.; Arnoldi, D.; Cagnacci, F.; Capelli, G.; Montarsi, F.; Rizzoli, A. Saturation deficit and deer density affect questing activity and local abundance of Ixodes ricinus (Acari, Ixodidae) in Italy. Vet. Parasitol. 2011, 183, 114–124. [Google Scholar] [CrossRef]
- Jaenson, T.G.; Jaenson, D.G.; Eisen, L.; Petersson, E.; Lindgren, E. Changes in the geographical distribution and abundance of the tick Ixodes ricinus during the past 30 years in Sweden. Parasites Vectors 2012, 5, 8. [Google Scholar] [CrossRef]
- Wu, X.; Duvvuri, V.R.; Lou, Y.; Ogden, N.H.; Pelcat, Y.; Wu, J. Developing a temperature-driven map of the basic reproductive number of the emerging tick vector of Lyme disease Ixodes scapularis in Canada. J. Theor. Biol. 2013, 319, 50–61. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Alexander, N.; Wint, G.W. Perspectives on modelling the distribution of ticks for large areas: So far so good? Parasites Vectors 2016, 9, 179. [Google Scholar] [CrossRef]
- Ogden, N.H.; Radojevic, M.; Wu, X.; Duvvuri, V.R.; Leighton, P.A.; Wu, J. Estimated effects of projected climate change on the basic reproductive number of the Lyme disease vector Ixodes scapularis. Environ. Health Perspect. 2014, 122, 631. [Google Scholar] [CrossRef]
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-Peña, A.; George, J.C.; Kazimirova, M. Driving forces for changes in geographical distribution of Ixodes ricinus ticks in Europe. Parasites Vectors 2013, 6, 1. [Google Scholar] [CrossRef]
- Laaksonen, M.; Sajanti, E.; Sormunen, J.J.; Penttinen, R.; Hänninen, J.; Ruohomäki, K.; Klemola, T. Crowdsourcing-based nationwide tick collection reveals the distribution of Ixodes ricinus and I. persulcatus and associated pathogens in Finland. Emerg. Microbes Infect. 2017, 6, e31. [Google Scholar] [CrossRef]
- Tälleklint, L.; Jaenson, T.G. Relationship between Ixodes ricinus density and prevalence of infection with Borrelia-like spirochetes and density of infected ticks. J. Med. Entomol. 2014, 33, 805–811. [Google Scholar] [CrossRef] [PubMed]
- Jaenson, T.G.; Petersson, E.H.; Jaenson, D.G.; Kindberg, J.; Pettersson, J.H.O.; Hjertqvist, M.; Bengtsson, H. The importance of wildlife in the ecology and epidemiology of the TBE virus in Sweden: Incidence of human TBE correlates with abundance of deer and hares. Parasites Vectors 2018, 11, 477. [Google Scholar] [CrossRef] [PubMed]
- Sormunen, J.J.; Klemola, T.; Vesterinen, E.J.; Vuorinen, I.; Hytönen, J.; Hänninen, J.; Penttinen, R. Assessing the abundance, seasonal questing activity, and Borrelia and tick-borne encephalitis virus (TBEV) prevalence of Ixodes ricinus ticks in a Lyme borreliosis endemic area in Southwest Finland. Ticks Tick-Borne Dis. 2016, 7, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Van den Besselaar, E.J.M.; Haylock, M.R.; Van der Schrier, G.; Klein Tank, A.M.G. A European daily high-resolution observational gridded data set of sea level pressure. J. Geophys. Res. Atmos. 2011, 116, D11110. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 1 September 2018).
- Mücher, C.A.; Klijn, J.A.; Wascher, D.M.; Schaminée, J.H. A new European Landscape Classification (LANMAP): A transparent, flexible and user-oriented methodology to distinguish landscapes. Ecol. Indicator 2010, 10, 87–103. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Estrada-Sánchez, D. Deconstructing Ixodes ricinus: A partial matrix model allowing mapping of tick development, mortality and activity rates. Med. Vet. Entomol. 2014, 28, 35–49. [Google Scholar] [CrossRef]
- Forkel, M.; Wutzler, T. Greenbrown—Land Surface Phenology and Trend Analysis. A Package for the R Software. Version 2.2, 2015-04-15. 2015. Available online: http://greenbrown.r-forge.r-project.org/ (accessed on 1 October 2016).
- Killick, R.; Eckley, I.A. changepoint: An R Package for Changepoint Analysis. J. Stat. Softw. 2014, 58, 1–19. Available online: http://www.jstatsoft.org/v58/i03/ (accessed on 1 October 2016). [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ecological Region | Trend 1950–2016 | AAT: 1950–1989 | AAT: 1990–2018 | % Change Between 1950–1989 and 1990–2018 |
|---|---|---|---|---|
| Alpine North | 8.38 | 0.96 | 267.00 | 86.90 |
| Alpine South | 8.91 | −2.19 | 65.32 | 26.29 |
| Atlantic Central | 7.63 | 3492.24 | 3745.04 | 7.25 |
| Atlantic North | 7.17 | 2871.40 | 3107.71 | 8.37 |
| Boreal | 10.66 | 742.51 | 1089.42 | 157.09 |
| Continental | 9.15 | 2890.30 | 3190.26 | 10.49 |
| Nemoral | 10.56 | 2135.75 | 2480.61 | 16.50 |
| Ecological Region | OV: Trend | INC: Trend | LN: Trend | NA: Trend |
|---|---|---|---|---|
| Alpine North | −0.69 | −0.83 | −0.89 | −0.50 |
| Alpine South | −0.42 | −0.69 | −0.78 | −0.36 |
| Atlantic Central | −0.31 | −0.42 | −0.47 | −0.23 |
| Atlantic North | −0.33 | −0.46 | −0.52 | −0.25 |
| Boreal | −1.35 | −1.78 | −1.94 | −1.15 |
| Continental | −0.42 | −0.63 | −0.72 | −0.34 |
| Nemoral | −0.69 | −1.09 | −1.25 | −0.56 |
| Ecoregion | OV: 1950–1989 | INC: 1950–1989 | LN: 1950–1989 | NA: 1950–1989 | OV: 1990–2018 | INC: 1990–2018 | LN: 1990–2018 | NA: 1990–2018 |
|---|---|---|---|---|---|---|---|---|
| Alpine North | 96.14 | 90.06 | 90.59 | 86.48 | 70.6 (23.49%) | 60.27 (31.47%) | 58.95 (33.43%) | 67.41 (21.05%) |
| Alpine South | 111.36 | 120.12 | 123.39 | 109.44 | 94 (15.58%) | 93.88 (21.84%) | 94.07 (23.76%) | 94.19 (13.93%) |
| Atlantic Central | 101.75 | 91.06 | 91.59 | 89.38 | 88.17 (23.49%) | 74.32 (18.03%) | 73.22 (19.65%) | 78.87 (11.59%) |
| Atlantic North | 104.21 | 91.74 | 92.79 | 88.42 | 89.82 (13.79%) | 73.8 (19.61%) | 73.01 (21.35%) | 77.34 (12.58%) |
| Boreal | 137.41 | 146.55 | 146.20 | 129.28 | 89.63 (34.01%) | 84.58 (42.55%) | 79.26 (46.07%) | 88.02 (31.06%) |
| Continental | 107.76 | 105.78 | 107.94 | 98.78 | 90.43 (16.06%) | 81.79 (22.76%) | 81.29 (24.76%) | 84.53 (14.48%) |
| Nemoral | 116.08 | 124.86 | 130.47 | 108.02 | 89.89 (22.91%) | 85.52 (32.18%) | 85.99 (34.89%) | 86.31 (20.33%) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Estrada-Peña, A.; Fernández-Ruiz, N. A Retrospective Assessment of Temperature Trends in Northern Europe Reveals a Deep Impact on the Life Cycle of Ixodes ricinus (Acari: Ixodidae). Pathogens 2020, 9, 345. https://doi.org/10.3390/pathogens9050345
Estrada-Peña A, Fernández-Ruiz N. A Retrospective Assessment of Temperature Trends in Northern Europe Reveals a Deep Impact on the Life Cycle of Ixodes ricinus (Acari: Ixodidae). Pathogens. 2020; 9(5):345. https://doi.org/10.3390/pathogens9050345
Chicago/Turabian StyleEstrada-Peña, Agustin, and Natalia Fernández-Ruiz. 2020. "A Retrospective Assessment of Temperature Trends in Northern Europe Reveals a Deep Impact on the Life Cycle of Ixodes ricinus (Acari: Ixodidae)" Pathogens 9, no. 5: 345. https://doi.org/10.3390/pathogens9050345
APA StyleEstrada-Peña, A., & Fernández-Ruiz, N. (2020). A Retrospective Assessment of Temperature Trends in Northern Europe Reveals a Deep Impact on the Life Cycle of Ixodes ricinus (Acari: Ixodidae). Pathogens, 9(5), 345. https://doi.org/10.3390/pathogens9050345