The Role of ST2 Receptor in the Regulation of Brucella abortus Oral Infection

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. The Absence of the ST2 Receptor Confers Partial Resistance to Oral Infection
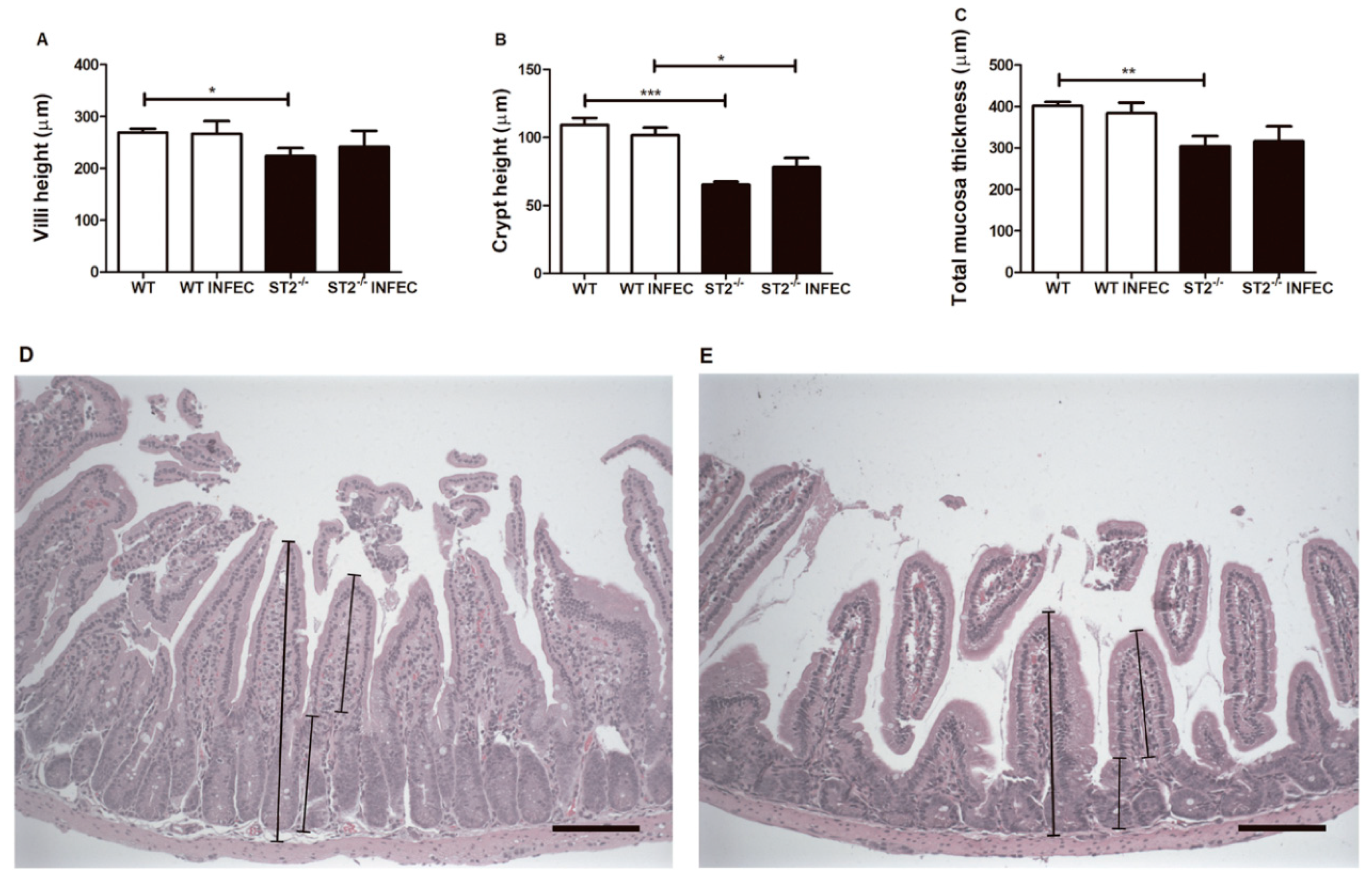
2.2. Absence of ST2 Resulted in Change of Intestinal Architecture
2.3. ST2 Receptor is Important in the Maintenance of the Intestinal Epithelial Barrier
2.4. Lack of ST2 Receptor Modulates the Recruitment of Neutrophils and Eosinophils and Increases the Production of IFN-γ and TNF-α in Small Intestine after Brucella abortus Infection
2.5. ST2 Receptor Does Not Play a Role in Systemic Infection Caused by Brucella abortus
2.6. The Absence of the ST2 Receptor Does Not alter the Production of Nitric Oxide by Macrophages
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Bacteria
4.3. Bacterial Counting in B. abortus Infected Mice
4.4. Intestinal Permeability Assay
4.5. Measurement of Myeloperoxidase (MPO) and Eosinophilic Peroxidase Activity (EPO)Activity
4.6. Measurement of Cytokine Concentrations
4.7. Real-Time PCR (RT–PCR)
4.8. Generation of BMDMs
4.9. Nitrite Measurement by Griess Reagent
4.10. Gut Pathology
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Moreno, E.; Moriyon, I. Brucella melitensis: A nasty bug with hidden credentials for virulence. Proc. Natl. Acad. Sci. USA 2002, 99, 1–3. [Google Scholar] [CrossRef] [PubMed]
- De Figueiredo, P.; Ficht, T.A.; Rice-Ficht, A.; Rossetti, C.A.; Adams, L.G. Pathogenesis and immunobiology of brucellosis: Review of Brucella-host interactions. Am. J. Pathol. 2015, 185, 1505–1517. [Google Scholar] [CrossRef] [PubMed]
- Moreno, E. Retrospective and prospective perspectives on zoonotic brucellosis. Front. Microbiol. 2014, 5, 213. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.R.; Heo, S.T.; Lee, K.H.; Kim, Y.R.; Yoo, S.J. Foodborne outbreak of human brucellosis caused by ingested raw materials of fetal calf on Jeju Island. Am. J. Trop. Med. Hyg. 2015, 92, 267–269. [Google Scholar] [CrossRef]
- Seleem, M.N.; Boyle, S.M.; Sriranganathan, N. Brucellosis: A re-emerging zoonosis. Vet. Microbiol. 2010, 140, 392–398. [Google Scholar] [CrossRef]
- Silva, F.L.; Paixao, T.A.; Borges, A.M. Brucelose Bovina. Cad. Tec. Vet. Zoot. 2005, 47, 1–12. [Google Scholar]
- Secretaria de Estado da Saúde do Paraná (Ed.) Protocolo de Manejo Clínico e Vigilância em Saúde para Brucelose Humana No Estado do Paraná; Secretaria de Estado da Saúde do Paraná: Curitiba, Brazil, 2015; p. 70.
- World Health Organization (WHO). Brucellosis in Humans and Animals; World Health Organization: Geneva, Switzerland, 2006; pp. 1–102. [Google Scholar]
- Solera, J. Update on brucellosis: Therapeutic challenges. Int. J. Antimicrob. Agents 2010, 36 (Suppl. 1), S18–S20. [Google Scholar] [CrossRef]
- Young, E.J. Brucellosis: A model zoonosis in developing countries. Apmis. Suppl. 1988, 3, 17–20. [Google Scholar]
- Dornand, J.; Gross, A.; Lafont, V.; Liautard, J.; Oliaro, J.; Liautard, J.P. The innate immune response against Brucella in humans. Vet. Microbiol. 2002, 90, 383–394. [Google Scholar] [CrossRef]
- Campos, M.A.; Rosinha, G.M.; Almeida, I.C.; Salgueiro, X.S.; Jarvis, B.W.; Splitter, G.A.; Qureshi, N.; Bruna-Romero, O.; Gazzinelli, R.T.; Oliveira, S.C. Role of Toll-like receptor 4 in induction of cell-mediated immunity and resistance to Brucella abortus infection in mice. Infect. Immun. 2004, 72, 176–186. [Google Scholar] [CrossRef]
- Huang, L.Y.; Ishii, K.J.; Akira, S.; Aliberti, J.; Golding, B. Th1-like cytokine induction by heat-killed Brucella abortus is dependent on triggering of TLR9. J. Immunol. 2005, 175, 3964–3970. [Google Scholar] [CrossRef] [PubMed]
- Weiss, D.S.; Takeda, K.; Akira, S.; Zychlinsky, A.; Moreno, E. MyD88, but not toll-like receptors 4 and 2, is required for efficient clearance of Brucella abortus. Infect. Immun. 2005, 73, 5137–5143. [Google Scholar] [CrossRef] [PubMed]
- Macedo, G.C.; Magnani, D.M.; Carvalho, N.B.; Bruna-Romero, O.; Gazzinelli, R.T.; Oliveira, S.C. Central role of MyD88-dependent dendritic cell maturation and proinflammatory cytokine production to control Brucella abortus infection. J. Immunol. 2008, 180, 1080–1087. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, S.C.; De Oliveira, F.S.; Macedo, G.C.; De Almeida, L.A.; Carvalho, N.B. The role of innate immune receptors in the control of Brucella abortus infection: Toll-like receptors and beyond. Microbes Infect. 2008, 10, 1005–1009. [Google Scholar] [CrossRef]
- Gomes, M.T.; Campos, P.C.; Pereira Gde, S.; Bartholomeu, D.C.; Splitter, G.; Oliveira, S.C. TLR9 is required for MAPK/NF-kappaB activation but does not cooperate with TLR2 or TLR6 to induce host resistance to Brucella abortus. J. Leukoc. Biol. 2016, 99, 771–780. [Google Scholar] [CrossRef]
- Gomes, M.T.; Campos, P.C.; Oliveira, F.S.; Corsetti, P.P.; Bortoluci, K.R.; Cunha, L.D.; Zamboni, D.S.; Oliveira, S.C. Critical role of ASC inflammasomes and bacterial type IV secretion system in caspase-1 activation and host innate resistance to Brucella abortus infection. J. Immunol. 2013, 190, 3629–3638. [Google Scholar] [CrossRef]
- Marim, F.M.; Franco, M.M.C.; Gomes, M.T.R.; Miraglia, M.C.; Giambartolomei, G.H.; Oliveira, S.C. The role of NLRP3 and AIM2 in inflammasome activation during Brucella abortus infection. Semin. Immunopathol. 2017, 39, 215–223. [Google Scholar] [CrossRef]
- Costa Franco, M.M.; Marim, F.; Guimaraes, E.S.; Assis, N.R.G.; Cerqueira, D.M.; Alves-Silva, J.; Harms, J.; Splitter, G.; Smith, J.; Kanneganti, T.D.; et al. Brucella abortus Triggers a cGAS-Independent STING Pathway To Induce Host Protection That Involves Guanylate-Binding Proteins and Inflammasome Activation. J. Immunol. 2018, 200, 607–622. [Google Scholar] [CrossRef]
- Rakoff-Nahoum, S.; Paglino, J.; Eslami-Varzaneh, F.; Edberg, S.; Medzhitov, R. Recognition of commensal microflora by toll-like receptors is required for intestinal homeostasis. Cell 2004, 118, 229–241. [Google Scholar] [CrossRef]
- Artis, D. Epithelial-cell recognition of commensal bacteria and maintenance of immune homeostasis in the gut. Nat. Rev. Immunol. 2008, 8, 411–420. [Google Scholar] [CrossRef]
- Barber, G.N. STING: Infection, inflammation and cancer. Nat. Rev. Immunol. 2015, 15, 760–770. [Google Scholar] [CrossRef]
- Hodzic, Z.; Schill, E.M.; Bolock, A.M.; Good, M. IL-33 and the intestine: The good, the bad, and the inflammatory. Cytokine 2017, 100, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, K.; Takagi, T.; Tsukamoto, T.; Tetsuka, T.; Tominaga, S. Presence of a novel primary response gene ST2L, encoding a product highly similar to the interleukin 1 receptor type 1. FEBS Lett. 1993, 318, 83–87. [Google Scholar] [CrossRef]
- Schmitz, J.; Owyang, A.; Oldham, E.; Song, Y.; Murphy, E.; McClanahan, T.K.; Zurawski, G.; Moshrefi, M.; Qin, J.; Li, X.; et al. IL-33, an interleukin-1-like cytokine that signals via the IL-1 receptor-related protein ST2 and induces T helper type 2-associated cytokines. Immunity 2005, 23, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Bergers, G.; Reikerstorfer, A.; Braselmann, S.; Graninger, P.; Busslinger, M. Alternative promoter usage of the Fos-responsive gene Fit-1 generates mRNA isoforms coding for either secreted or membrane-bound proteins related to the IL-1 receptor. EMBO J. 1994, 13, 1176–1188. [Google Scholar] [CrossRef] [PubMed]
- Iwahana, H.; Yanagisawa, K.; Ito-Kosaka, A.; Kuroiwa, K.; Tago, K.; Komatsu, N.; Katashima, R.; Itakura, M.; Tominaga, S. Different promoter usage and multiple transcription initiation sites of the interleukin-1 receptor-related human ST2 gene in UT-7 and TM12 cells. Eur. J. Biochem. 1999, 264, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Lohning, M.; Stroehmann, A.; Coyle, A.J.; Grogan, J.L.; Lin, S.; Gutierrez-Ramos, J.C.; Levinson, D.; Radbruch, A.; Kamradt, T. T1/ST2 is preferentially expressed on murine Th2 cells, independent of interleukin 4, interleukin 5, and interleukin 10, and important for Th2 effector function. Proc. Natl. Acad. Sci. USA 1998, 95, 6930–6935. [Google Scholar] [CrossRef] [PubMed]
- Neill, D.R.; Wong, S.H.; Bellosi, A.; Flynn, R.J.; Daly, M.; Langford, T.K.A.; Bucks, C.; Kane, C.M.; Fallon, P.G.; Pannell, R.; et al. Nuocytes represent a new innate effector leukocyte that mediates type-2 immunity. Nature 2010, 464, U1367–U1369. [Google Scholar] [CrossRef]
- Kurowska-Stolarska, M.; Stolarski, B.; Kewin, P.; Murphy, G.; Corrigan, C.J.; Ying, S.; Pitman, N.; Mirchandani, A.; Rana, B.; Van Rooijen, N.; et al. IL-33 Amplifies the Polarization of Alternatively Activated Macrophages That Contribute to Airway Inflammation. J. Immunol. 2009, 183, 6469–6477. [Google Scholar] [CrossRef]
- Cherry, W.B.; Yoon, J.; Barternes, K.R.; Iijima, K.; Kita, H. A novel IL-1 family cytokine, IL-33, potently activates human eosinophils. J. Allergy Clin. Immunol. 2008, 121, 1484–1490. [Google Scholar] [CrossRef]
- Suzukawa, M.; Iikura, M.; Koketsu, R.; Nagase, H.; Tamura, C.; Komiya, A.; Nakae, S.; Matsushima, K.; Ohta, K.; Yamamoto, K.; et al. An IL-1 Cytokine Member, IL-33, Induces Human Basophil Activation via Its ST2 Receptor. J. Immunol. 2008, 181, 5981–5989. [Google Scholar] [CrossRef] [PubMed]
- Smithgall, M.D.; Comeau, M.R.; Yoon, B.R.P.; Kaufman, D.; Armitage, R.; Smith, D.E. IL-33 amplifies both T(h)1-and T(h)2-type responses through its activity on human basophils, allergen-reactive T(h)2 cells, iNKT and NK Cells. Int. Immunol. 2008, 20, 1019–1030. [Google Scholar] [CrossRef] [PubMed]
- Martin, N.T.; Martin, M.U. Interleukin 33 is a guardian of barriers and a local alarmin. Nat. Immunol. 2016, 17, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Le, H.; Kim, W.; Kim, J.; Cho, H.R.; Kwon, B. Interleukin-33: A mediator of inflammation targeting hematopoietic stem and progenitor cells and their progenies. Front. Immunol. 2013, 4, 104. [Google Scholar] [CrossRef] [PubMed]
- Mahapatro, M.; Foersch, S.; Hefele, M.; He, G.W.; Giner-Ventura, E.; McHedlidze, T.; Kindermann, M.; Vetrano, S.; Danese, S.; Gunther, C.; et al. Programming of Intestinal Epithelial Differentiation by IL-33 Derived from Pericryptal Fibroblasts in Response to Systemic Infection. Cell Rep. 2016, 15, 1743–1756. [Google Scholar] [CrossRef] [PubMed]
- Liew, F.Y.; Girard, J.P.; Turnquist, H.R. Interleukin-33 in health and disease. Nat. Rev. Immunol. 2016, 16, 676–689. [Google Scholar] [CrossRef] [PubMed]
- Hodzic, E.; Granov, N. Gigantic Thrombus of the Left Atrium in Mitral Stenosis. Med. Arch. 2017, 71, 449–452. [Google Scholar] [CrossRef]
- Pham, O.H.; O’Donnell, H.; Al-Shamkhani, A.; Kerrinnes, T.; Tsolis, R.M.; McSorley, S.J. T cell expression of IL-18R and DR3 is essential for non-cognate stimulation of Th1 cells and optimal clearance of intracellular bacteria. PLoS Pathog. 2017, 13, e1006566. [Google Scholar] [CrossRef]
- Brandao, A.P.; Oliveira, F.S.; Carvalho, N.B.; Vieira, L.Q.; Azevedo, V.; Macedo, G.C.; Oliveira, S.C. Host susceptibility to Brucella abortus infection is more pronounced in IFN-gamma knockout than IL-12/beta2-microglobulin double-deficient mice. Clin. Dev. Immunol. 2012, 2012, 589494. [Google Scholar] [CrossRef]
- Pasquevich, K.A.; Carabajal, M.V.; Guaimas, F.F.; Bruno, L.; Roset, M.S.; Coria, L.M.; Rey Serrantes, D.A.; Comerci, D.J.; Cassataro, J. Omp19 Enables Brucella abortus to Evade the Antimicrobial Activity from Host’s Proteolytic Defense System. Front. Immunol. 2019, 10, 1436. [Google Scholar] [CrossRef]
- Sangari, F.J.; Seoane, A.; Rodriguez, M.C.; Aguero, J.; Garcia Lobo, J.M. Characterization of the urease operon of Brucella abortus and assessment of its role in virulence of the bacterium. Infect. Immun. 2007, 75, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Delpino, M.V.; Marchesini, M.I.; Estein, S.M.; Comerci, D.J.; Cassataro, J.; Fossati, C.A.; Baldi, P.C. A bile salt hydrolase of Brucella abortus contributes to the establishment of a successful infection through the oral route in mice. Infect. Immun. 2007, 75, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Brenchley, J.M.; Douek, D.C. Microbial translocation across the GI tract. Annu. Rev. Immunol. 2012, 30, 149–173. [Google Scholar] [CrossRef] [PubMed]
- Lumsden, A.B.; Henderson, J.M.; Kutner, M.H. Endotoxin levels measured by a chromogenic assay in portal, hepatic and peripheral venous blood in patients with cirrhosis. Hepatology 1988, 8, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Bullard, J.; Kalra, M.; Assefa, S.; Kaul, A.K.; Vonfeldt, K.; Strom, S.C.; Conrad, R.S.; Sharp, H.L.; Kaul, R. Status of bacterial colonization, Toll-like receptor expression and nuclear factor-kappa B activation in normal and diseased human livers. Clin. Immunol. 2011, 138, 41–49. [Google Scholar] [CrossRef]
- Neutra, M.R.; Mantis, N.J.; Kraehenbuhl, J.P. Collaboration of epithelial cells with organized mucosal lymphoid tissues. Nat. Immunol. 2001, 2, 1004–1009. [Google Scholar] [CrossRef]
- Nakato, G.; Hase, K.; Suzuki, M.; Kimura, M.; Ato, M.; Hanazato, M.; Tobiume, M.; Horiuchi, M.; Atarashi, R.; Nishida, N.; et al. Cutting Edge: Brucella abortus exploits a cellular prion protein on intestinal M cells as an invasive receptor. J. Immunol. 2012, 189, 1540–1544. [Google Scholar] [CrossRef]
- Paixao, T.A.; Roux, C.M.; Den Hartigh, A.B.; Sankaran-Walters, S.; Dandekar, S.; Santos, R.L.; Tsolis, R.M. Establishment of systemic Brucella melitensis infection through the digestive tract requires urease, the type IV secretion system, and lipopolysaccharide O antigen. Infect. Immun. 2009, 77, 4197–4208. [Google Scholar] [CrossRef]
- Zhao, X.; Yang, J.; Ju, Z.; Wu, J.; Wang, L.; Lin, H.; Sun, S. Clostridium butyricum Ameliorates Salmonella Enteritis Induced Inflammation by Enhancing and Improving Immunity of the Intestinal Epithelial Barrier at the Intestinal Mucosal Level. Front. Microbiol. 2020, 11, 299. [Google Scholar] [CrossRef]
- Ferrero, M.C.; Fossati, C.A.; Rumbo, M.; Baldi, P.C. Brucella invasion of human intestinal epithelial cells elicits a weak proinflammatory response but a significant CCL20 secretion. FEMS Immunol. Med. Microbiol. 2012, 66, 45–57. [Google Scholar] [CrossRef]
- Ackermann, M.R.; Cheville, N.F.; Deyoe, B.L. Bovine ileal dome lymphoepithelial cells: Endocytosis and transport of Brucella abortus strain 19. Vet. Pathol. 1988, 25, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Rossetti, C.A.; Drake, K.L.; Siddavatam, P.; Lawhon, S.D.; Nunes, J.E.; Gull, T.; Khare, S.; Everts, R.E.; Lewin, H.A.; Adams, L.G. Systems biology analysis of Brucella infected Peyer’s patch reveals rapid invasion with modest transient perturbations of the host transcriptome. PLoS ONE 2013, 8, e81719. [Google Scholar] [CrossRef]
- Churin, Y.; Al-Ghoul, L.; Kepp, O.; Meyer, T.F.; Birchmeier, W.; Naumann, M. Helicobacter pylori CagA protein targets the c-Met receptor and enhances the motogenic response. J. Cell Biol. 2003, 161, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Sheng, H. Amphiregulin promotes intestinal epithelial regeneration: Roles of intestinal subepithelial myofibroblasts. Endocrinology 2010, 151, 3728–3737. [Google Scholar] [CrossRef] [PubMed]
- Shoyab, M.; McDonald, V.L.; Bradley, J.G.; Todaro, G.J. Amphiregulin: A bifunctional growth-modulating glycoprotein produced by the phorbol 12-myristate 13-acetate-treated human breast adenocarcinoma cell line MCF-7. Proc. Natl. Acad. Sci. USA 1988, 85, 6528–6532. [Google Scholar] [CrossRef] [PubMed]
- Ranganathan, S.; Doucet, M.; Grassel, C.L.; Delaine-Elias, B.; Zachos, N.C.; Barry, E.M. Evaluating Shigella flexneri Pathogenesis in the Human Enteroid Model. Infect. Immun. 2019, 87. [Google Scholar] [CrossRef]
- Rathinam, V.A.K.; Chan, F.K. Inflammasome, Inflammation, and Tissue Homeostasis. Trends Mol. Med. 2018, 24, 304–318. [Google Scholar] [CrossRef]
- Elinav, E.; Thaiss, C.A.; Flavell, R.A. Analysis of microbiota alterations in inflammasome-deficient mice. Methods Mol. Biol. 2013, 1040, 185–194. [Google Scholar] [CrossRef]
- Camilleri, M.; Madsen, K.; Spiller, R.; Van Meerveld, B.G.; Verne, G.N. Intestinal barrier function in health and gastrointestinal disease (vol 24, pg 503, 2012). Neurogastroenterol. Motil. 2012, 24, 976. [Google Scholar] [CrossRef]
- Gutierrez-Jimenez, C.; Mora-Cartin, R.; Altamirano-Silva, P.; Chacon-Diaz, C.; Chaves-Olarte, E.; Moreno, E.; Barquero-Calvo, E. Neutrophils as Trojan Horse Vehicles for Brucella abortus Macrophage Infection. Front. Immunol. 2019, 10, 1012. [Google Scholar] [CrossRef]
- Staurengo-Ferrari, L.; Trevelin, S.C.; Fattori, V.; Nascimento, D.C.; De Lima, K.A.; Pelayo, J.S.; Figueiredo, F.; Casagrande, R.; Fukada, S.Y.; Teixeira, M.M.; et al. Interleukin-33 Receptor (ST2) Deficiency Improves the Outcome of Staphylococcus aureus-Induced Septic Arthritis. Front. Immunol. 2018, 9, 962. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Kolachala, V.; Dalmasso, G.; Nguyen, H.; Laroui, H.; Sitaraman, S.V.; Merlin, D. Temporal and spatial analysis of clinical and molecular parameters in dextran sodium sulfate induced colitis. PLoS ONE 2009, 4, e6073. [Google Scholar] [CrossRef] [PubMed]
- Vieira, A.T.; Fagundes, C.T.; Alessandri, A.L.; Castor, M.G.; Guabiraba, R.; Borges, V.O.; Silveira, K.D.; Vieira, E.L.; Goncalves, J.L.; Silva, T.A.; et al. Treatment with a novel chemokine-binding protein or eosinophil lineage-ablation protects mice from experimental colitis. Am. J. Pathol. 2009, 175, 2382–2391. [Google Scholar] [CrossRef] [PubMed]
- Weischenfeldt, J.; Porse, B. Bone Marrow-Derived Macrophages (BMM): Isolation and Applications. Cold Spring Harb. Protoc. 2008, 2008, pdb-prot5080. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, R.; Campos, P.C.; Rungue, M.; Rocha, V.; Santos, D.; Mendes, V.; Marinho, F.V.; Martins, F.; Ricci, M.F.; dos Reis, D.C.; et al. The Role of ST2 Receptor in the Regulation of Brucella abortus Oral Infection. Pathogens 2020, 9, 328. https://doi.org/10.3390/pathogens9050328
Santos R, Campos PC, Rungue M, Rocha V, Santos D, Mendes V, Marinho FV, Martins F, Ricci MF, dos Reis DC, et al. The Role of ST2 Receptor in the Regulation of Brucella abortus Oral Infection. Pathogens. 2020; 9(5):328. https://doi.org/10.3390/pathogens9050328
Chicago/Turabian StyleSantos, Raiany, Priscila C. Campos, Marcella Rungue, Victor Rocha, David Santos, Viviani Mendes, Fabio V. Marinho, Flaviano Martins, Mayra F. Ricci, Diego C. dos Reis, and et al. 2020. "The Role of ST2 Receptor in the Regulation of Brucella abortus Oral Infection" Pathogens 9, no. 5: 328. https://doi.org/10.3390/pathogens9050328
APA StyleSantos, R., Campos, P. C., Rungue, M., Rocha, V., Santos, D., Mendes, V., Marinho, F. V., Martins, F., Ricci, M. F., dos Reis, D. C., D. Cassali, G., Alves-Filho, J. C., Vieira, A. T., & Oliveira, S. C. (2020). The Role of ST2 Receptor in the Regulation of Brucella abortus Oral Infection. Pathogens, 9(5), 328. https://doi.org/10.3390/pathogens9050328