Efficient Transovarial Transmission of Babesia Spp. in Rhipicephalus microplus Ticks Fed on Water Buffalo (Bubalus bubalis)

 ,
, 
 ,
, 

Abstract
1. Introduction
2. Results and Discussion
3. Materials and Methods
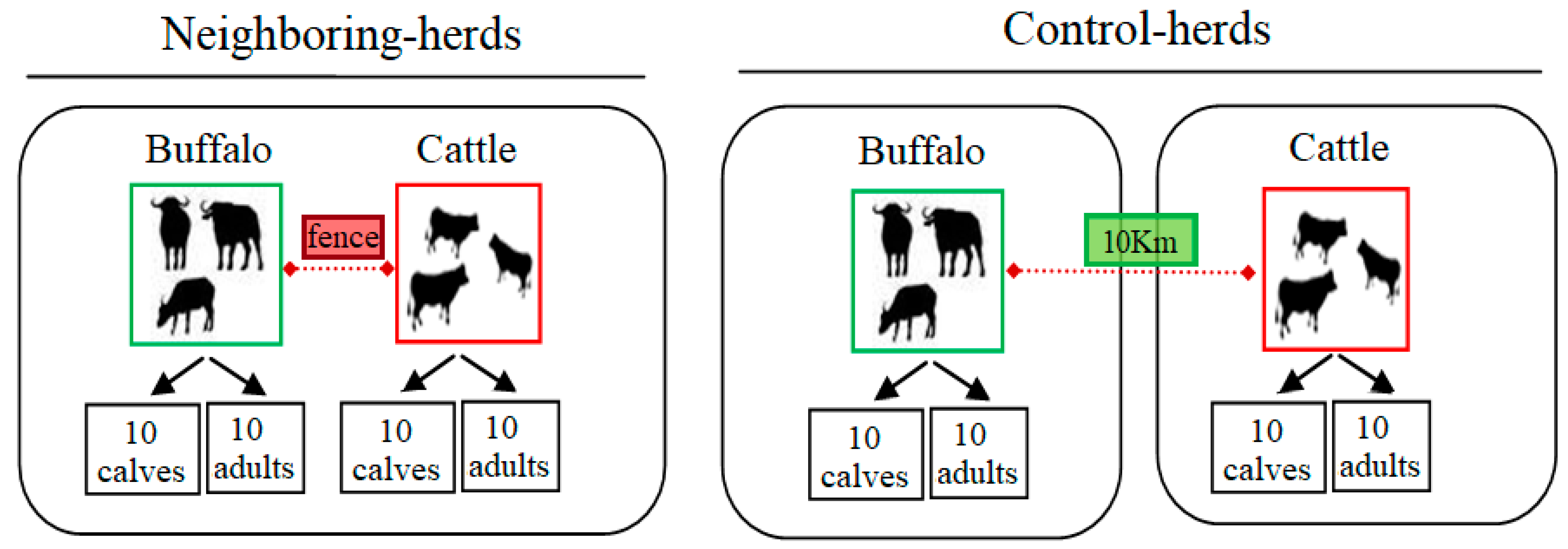
3.1. Study Site and Samples’ Selection
3.2. Tick Infestation
3.3. Analysis of the Reproductive Parameters of Adult Female Ticks
3.4. DNA Extraction and Quantification of Babesia Infection Levels
3.5. Statistical Analysis
3.6. Ethics Statement
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability Statement
References
- Suarez, C.E.; Noh, S. Emerging perspectives in the research of bovine babesiosis and anaplasmosis. Vet. Parasitol. 2011, 180, 109–125. [Google Scholar] [PubMed]
- OIE. Babesiosis bovina. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2012; World Organization for Animal Health: Paris, France, 2012; pp. 1–16. [Google Scholar]
- Nari, A. Strategies for the control of one-host ticks and relationship with tick-borne diseases in South America. Vet. Parasitol. 1995, 57, 153–165. [Google Scholar] [CrossRef]
- Alonso, M.; Arellano-Sota, C.; Cereser, V.H.; Cordoves, C.O.; Kessler, A.A.; Solari, M.A.; Vega, C.A. Epidemiology of bovine anaplasmosis and babesiosis in Latin America and the Caribbean. Int. J. Epidemiol. 1992, 11, 713–733. [Google Scholar]
- Bock, R.; Jackson, L.; Vos, A.; Jorgensen, W. Babesiosis of cattle. Parasitology 2004, 129, S247–S269. [Google Scholar] [CrossRef]
- Chauvin, A.C.; Oreau, E.M.; Onnet, S.B.; Lantard, O.P.; Alandrin, L.M.; Moreau, E.; Bonnet, S.; Plantard, O.; Malandrin, L. Babesia and its hosts: Adaptation to long-lasting interactions as a way to achieve efficient transmission. Vet. Res. 2009, 40, 37. [Google Scholar] [CrossRef]
- Cantu, A.; Ortega-S, J.A.; Mosqueda, J.; Garcia-Vazquez, Z.; Henke, S.E.; George, J.E. Immunologic and molecular identification of Babesia bovis and Babesia bigemina in free-ranging white-tailed deer in northern Mexico. J. Wildl. Dis. 2007, 43, 504–507. [Google Scholar] [CrossRef]
- Iseki, H.; Alhassan, A.; Ohta, N.; Thekisoe, O.M.M.; Yokoyama, N.; Inoue, N.; Nambota, A.; Yasuda, J.; Igarashi, I. Development of a multiplex loop-mediated isothermal amplification (mLAMP) method for the simultaneous detection of bovine Babesia parasites. J. Microbiol. Methods 2007, 71, 281–287. [Google Scholar] [CrossRef]
- Obregón, D.; Oliveira, M.C.S.; Tizioto, P.; Funnes, E.M.; Martínez, S.; Roque, E.; Fonseca, A.H.; Corona, B. Diagnóstico de Babesia bovis en búfalos de la región occidental de Cuba a través de un ensayo de nPCR. Rev. Salud Anim. 2012, 34, 101–108. [Google Scholar]
- Barros-Battesti, D.; Reyes, M.H.; Onofrio, V.C.; Beati, L.; Famada, K.; Guglielmone, A.A. The ixodid ticks (Acari: Ixodidae) of Cuba. Syst. Appl. Acarol. 2009, 14, 101–128. [Google Scholar]
- Gaínza, Y.A.; Martínez-Marrero, S.; Corona-González, B. Ticks of veterinary interest in Cuba, and its importance in the changing climatic conditions. Rev. Electrónica Vet. 2014, 15, 1–22. [Google Scholar]
- Estrada-Peña, A.; Bouattour, A.; Camicas, J.-L.; Guglielmone, A.; Horak, I.; Jongejan, F.; Latif, A.; Pegram, R.; Walker, A. The known distribution and ecological preferences of the tick subgenus Boophilus (Acari: Ixodidae) in Africa and Latin America. Exp. Appl. Acarol. 2006, 38, 219–235. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Peña, A.; Naranjo, V.; Acevedo-Whitehouse, K.; Mangold, A.J.; Kocan, K.M.; De la Fuente, J. Phylogeographic analysis reveals association of tick-borne pathogen, Anaplasma marginale, MSP1a sequences with ecological traits affecting tick vector performance. BMC Biol. 2009, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Obregón, D.; Rodríguez, J.D.; Roque, E.; Alemán, Y. Rhipicephalus (Boophilus) microplus (acari: Ixodidae) en búfalos (Bubalus bubalis), en Cuba. Rev. Salud Anim. 2010, 32, 132–134. [Google Scholar]
- Benitez, D.; Cetrá, B.; Florin-Christensen, M. Rhipicephalus (Boophilus) microplus ticks can complete their life cycle on the water buffalo (Bubalus bubalis). J. Buffalo Sci. 2012, 1, 193–197. [Google Scholar]
- Mitat, A.O.B. Búfalos de agua en Cuba: Origen y evolución. Rev. ACPA 2009, 3, 45–48. [Google Scholar]
- Ferreri, L.; Benitez, D.; Dominguez, M.; Rodriguez, A.; Asenzo, G.; Mesplet, M.; Florin-Christensen, M.; Schnittger, L. Water Buffalos as carriers of Babesia bovis in Argentina. Ann. N. Y. Acad. Sci. 2008, 1149, 149–151. [Google Scholar] [CrossRef]
- Néo, T.A.; Giglioti, R.; Obregón, D.; Bilhassi, T.B.; Oliveira, H.N.; Machado, R.Z.; Aníbal, F.F.; Brito, L.G.; Malagó, W.; Bressani, F.A.; et al. Detection of Babesia bovis and B. bigemina in Water buffaloes (Bubalus bubalis) in endemic areas of São Paulo State, Brazil. Open J. Vet. Med. 2016, 6, 75–84. [Google Scholar] [CrossRef]
- Terkawi, M.A.; Huyen, N.X.; Shinuo, C.; Inpankaew, T.; Maklon, K.; Aboulaila, M.; Ueno, A.; Goo, Y.K.; Yokoyama, N.; Jittapalapong, S.; et al. Molecular and serological prevalence of Babesia bovis and Babesia bigemina in water buffaloes in the northeast region of Thailand. Vet. Parasitol. 2011, 178, 201–207. [Google Scholar] [CrossRef]
- Romero-Salas, D.; Mira, A.; Mosqueda, J.; García-Vázquez, Z.; Hidalgo-Ruiz, M.; Vela, N.A.O.; de León, A.A.P.; Florin-Christensen, M.; Schnittger, L. Molecular and serological detection of Babesia bovis- and Babesia bigemina-infection in bovines and water buffaloes raised jointly in an endemic field. Vet. Parasitol. 2016, 217, 101–107. [Google Scholar] [CrossRef]
- Silva, J.B.; André, M.R.; da Fonseca, A.H.; de Albuquerque Lopes, C.T.; da Silva Lima, D.H.; de Andrade, S.J.T.; Oliveira, C.M.C.; Barbosa, J.D. Molecular and serological prevalence of Babesia bovis and Babesia bigemina in water buffaloes in the north region of Brazil. Vet. Parasitol. 2013, 197, 678–681. [Google Scholar] [CrossRef]
- Obregón, D.; Cabezas-cruz, A.; Armas, Y.; Silva, J.B.; Fonseca, A.H.; André, M.R.; Alfonso, P.; Oliveira, M.C.S.; Machado, R.Z.; Corona-González, B. High co-infection rates of Babesia bovis, Babesia bigemina, and Anaplasma marginale in water buffalo in Western Cuba. Parasitol. Res. 2019, 118, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Nithikathkul, C.; Polseela, P.; Changsap, B.; Leemingsawat, S. Ixodid ticks on domestic animals in Samut Prakan Province, Thailand. Southeast Asian J. Trop. Med. Public Health 2002, 33, 41–44. [Google Scholar] [PubMed]
- Gomes, R.A.; Machado, R.Z.; Starke-buzetti, W.A.; Bonesso, M.A. Immune-humoral response of water buffalo (Bubalus bubalis) against Anaplasma marginale. Rev. Bras. Parasitol. Vet. 2008, 17, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Gomes, R.A. Resposta Imune-Humoral de Búfalos (Bubalus Bubalis) Infectados Naturalmente por Babesia Bovis, B. Bigemina e Anaplasma Marginale; UNESP-Campus de Jaboticabal: Jaboticabal, Brazil, 2007. [Google Scholar]
- Tabor, A.E.; Ali, A.; Rehman, G.; Garcia, G.R.; Zangirolamo, A.F.; Malardo, T.; Jonsson, N.N. Cattle Tick Rhipicephalus microplus-host interface: A review of resistant and susceptible host responses. Front. Cell. Infect. Microbiol. 2017, 7, 1–18. [Google Scholar] [CrossRef]
- Porto Neto, L.R.; Jonsson, N.N.; D’Occhio, M.J.; Barendse, W. Molecular genetic approaches for identifying the basis of variation in resistance to tick infestation in cattle. Vet. Parasitol. 2011, 180, 165–172. [Google Scholar] [CrossRef]
- Robbertse, L.; Richards, S.A.; Maritz-Olivier, C. Bovine immune factors underlying tick resistance: Integration and future directions. Front. Cell. Infect. Microbiol. 2017, 7, 1–16. [Google Scholar] [CrossRef]
- Constantinoiu, C.C.; Jackson, L.A.; Jorgensen, W.K.; Lew-Tabor, A.E.; Piper, E.K.; Mayer, D.G.; Venus, B.; Jonsson, N.N. Local immune response against larvae of Rhipicephalus (Boophilus) microplus in Bos taurus indicus and Bos taurus taurus cattle. Int. J. Parasitol. 2010, 40, 865–875. [Google Scholar] [CrossRef]
- Piper, E.K.; Jackson, L.A.; Bielefeldt-Ohmann, H.; Gondro, C.; Lew-Tabor, A.E.; Jonsson, N.N. Tick-susceptible Bos taurus cattle display an increased cellular response at the site of larval Rhipicephalus (Boophilus) microplus attachment, compared with tick-resistant Bos indicus cattle. Int. J. Parasitol. 2010, 40, 431–441. [Google Scholar] [CrossRef]
- Quintão-Silva, M.G.; Melo, M.N.; Ribeiro, M.F.B. Comparison of duplex PCR and microscopic techniques for the identification of Babesia bigemina and Babesia bovis in engorged female ticks of Boophilus microplus. Zoonoses Public Health 2007, 54, 147–151. [Google Scholar] [CrossRef]
- Mahoney, D.F.; Mirre, G.B. Bovine babesiasis: Estimation of infection rates in the tick vector Boophilus microplus (Canestrini). Ann. Trop. Med. Parasitol. 1971, 65, 309–317. [Google Scholar] [CrossRef]
- Jonsson, N.N.; Bock, R.E.; Jorgensen, W.K.; Morton, J.M.; Stear, M.J. Is endemic stability of tick-borne disease in cattle a useful concept? Trends Parasitol. 2012, 28, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Gohil, S.; Kats, L.M.; Sturm, A.; Cooke, B.M. Recent insights into alteration of red blood cells by Babesia bovis: Moovin’ forward. Trends Parasitol. 2010, 26, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Giglioti, R.; de Oliveira, H.N.; Okino, C.H.; de Sena Oliveira, M.C. qPCR estimates of Babesia bovis and Babesia bigemina infection levels in beef cattle and Rhipicephalus microplus larvae. Exp. Appl. Acarol. 2018, 75, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.Y.X.; Langevelde, F.; Estrada-Peña, A.; Suzán, G.; de Boer, F. The diversity-disease relationship: Evidence and criticisms of the dilution effect. Parasitology 2016, 1, 1–12. [Google Scholar] [CrossRef] [PubMed]
- INSMET. Instituto de Meteorología de la República de CUBA. Centro del Clima. El Clima de Cuba. Características Generales. Available online: http://www.met.inf.cu/asp/genesis.asp?TB0=PLANTILLAS&TB1=CLIMAC&TB2=/clima/ClimaCuba.htm (accessed on 1 January 2016).
- LNP (National Parasitology Laboratory). Serie Histórica de la Anaplasmosis y Babesiosis en Cuba; Sistema de información estadística del IMV. Annual balance report: 2014; Ministry of Agriculture of the Republic of Cuba: Habana, Cuba, 2014. [Google Scholar]
- Obregón, D.; Corona, B.G.; de la Fuente, J.; Cabezas-Cruz, A.; Gonçalves, L.R.; Matos, C.A.; Armas, Y.; Hinojosa, Y.; Alfonso, P.; Oliveira, M.C.S.; et al. Molecular evidence of the reservoir competence of water buffalo (Bubalus bubalis) for Anaplasma marginale in Cuba. Vet. Parasitol. Reg. Stud. Reports 2018, 13, 180–187. [Google Scholar] [CrossRef] [PubMed]
- De la Vega, R.; Camejo, A.; Dias, G. Dynamics of Boophilus microplus (Acari: Ixodidae) female feeding on bovine. Rev. Salud Anim. 2003, 25, 192–195. [Google Scholar]
- De la Vega, R.; Moreno, A.; Diaz, G. Sampling method of the cattle tick (Boophilus microplus) in dairy cows. Rev. Salud Anim. 1984, 6, 397–406. [Google Scholar]
- De la Vega, R.; Díaz, G.; Camejo, A.; García, I. Application of tick sampling on acaricidal bath frequency in Rhipicephalus microplus (acari: Ixodidae). Rev. Salud Anim. 2009, 31, 105–107. [Google Scholar]
- Veríssimo, C.J.; da Silva, R.G.; Oliveira, A.D.; Ribeiro, W.R.; Rocha, U.F. Boophilus micropius tick instar counts on a crossbred herd. B. Ind. Anim 1997, 54, 19–24. [Google Scholar]
- Oliveira, M.C.S.; Oliveira-Sequeira, T.C.G.; Araujo, J.P.; Amarante, A.F.T.; Oliveira, H.N. Babesia spp. infection in Boophilus microplus engorged females and eggs in Sao Paulo State, Brazil. Vet. Parasitol. 2005, 130, 61–67. [Google Scholar] [CrossRef]
- Giglioti, R.; Forim, M.R.; Oliveira, H.N.; Chagas, A.C.S.; Ferrezini, J.; Brito, L.G.; Falcoski, T.O.R.S.; Albuquerque, L.G.; Oliveira, M.C.S. In vitro acaricidal activity of neem (Azadirachta indica) seed extracts with known azadirachtin concentrations against Rhipicephalus microplus. Vet. Parasitol. 2011, 181, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Buling, A.; Criado-Fornelio, A.; Asenzo, G.; Benitez, D.; Barba-Carretero, J.C.; Florin-Christensen, M. A quantitative PCR assay for the detection and quantification of Babesia bovis and B. bigemina. Vet. Parasitol. 2007, 147, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Obregón, D.; Rabelo, M.D.; Giglioti, R.; Bilhassi, T.B.; Néo, T.A.; Corona, B.; Alfonso, P.; Machado, R.Z.; Oliveira, M.C.S. Standardization of a SYBR green based real-time PCR system for detection and molecular quantification of Babesia bovis and B. bigemina in water buffaloes (Bubalus bubalis). J. Buffalo Sci. 2016, 5, 44–52. [Google Scholar] [CrossRef]


{kind=link}
| Neighboring-Herds | Control-Herds | ||||
|---|---|---|---|---|---|
| Indicators * | Age Group | Cattle | Buffalo | Cattle | Buffalo |
| (TIP) Tick infestation prevalence | Adults | 80% | 0% | 60% | 0% |
| Calves | 50% | 100% | 70% | 60% | |
| (TIR) Tick infestation rate ( ± SD) | Adults | 7 ± 3ab | 0 | 5 ± 2b | 0 |
| Calves | 7 ± 4ab | 12 ± 10a | 5 ± 3b | 5 ± 4b | |
| Nymphs/larvae ratio ( ± SE) | Adults | - | - | - | - |
| Calves | 36 ± 0.009%c | 30 ± 0.009%b | 35 ± 0.009%c | 24 ± 0.008%a | |
| Neighboring-Herds | Control-Herds | ||||
|---|---|---|---|---|---|
| Indicators | Age Group | Cattle | Buffalo | Cattle | Buffalo |
| (CE) Conversion efficiency ( ± SE) | Adults | 43 ± 0.04% | - | 42 ± 0.07% | - |
| Calves | 38 ± 0.05% | 49 ± 0.03% | 41 ± 0.05% | 50 ± 0.07% | |
| (IR) Incubation rate ( ± SE) | Adults | 98 ± 0.02% | - | 99 ± 0.04% | - |
| Calves | 98 ± 0.02% | 95 ± 0.01% | 96 ± 0.03% | 99 ± 0.04% | |
| (REI) Reprod. efficiency index ( ± SD) | Adults | 8.4 ± 2.0 × 105 | - | 8.4 ± 5.2 × 105 | - |
| Calves | 7.4 ± 3.0 × 105 | 9.5 ± 2.2 × 105 | 7.9 ± 1.6 × 105 | 9.8 ± 2.0 × 105 | |
| Neighboring-Herds | Control-Herds | ||||
|---|---|---|---|---|---|
| Hemoparasite * | Age Group | Cattle | Buffalo | Cattle | Buffalo |
| B. bovis (LogCN/µL) | Adults | 8.5 ± 1.3b | - | 8.6 ± 1.4b | - |
| Calves | 7.8 ± 0.7b | 7.3 ± 0.6a | 7.7 ± 1.0b | 7.2 ± 0.5a | |
| B. bigemina (LogCN/µL) | Adults | 5.3 ± 1.0ab | - | 4.8 ± 1.4a | - |
| Calves | 7.2 ± 1.3b | 5.6 ± 0.7a | 6.0 ± 2.0ab | 5.4 ± 0.4a | |
| Neighboring-Herds | Control-Herds | ||||
|---|---|---|---|---|---|
| Hemoparasite * | Age Group | Cattle | Buffalo | Cattle | Buffalo |
| B. bovis (LogCN/µL) | Adults | 8.1 ± 0.3c | 6.6 ± 1.4ab | 8.0 ± 0.0bc | 7.5 ± 1.0bc |
| Calves | 8.3 ± 0.3c | 6.7 ± 0.9a | 8.1 ± 0.3c | 7.5 ± 0.1abc | |
| B. bigemina (LogCN/µL) | Adults | 6.1 ± 0.8b | 5.0 ± 1.0ab | 6.3 ± 1.1abc | 4.8 ± 1.8ab |
| Calves | 9.0 ± 0.7d | 6.5 ± 0.7ab | 8.4 ± 0.7cd | 5.0 ± 0.1a | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Obregón, D.; Corona-González, B.; Díaz-Sánchez, A.A.; Armas, Y.; Roque, E.; de Sena Oliveira, M.C.; Cabezas-Cruz, A. Efficient Transovarial Transmission of Babesia Spp. in Rhipicephalus microplus Ticks Fed on Water Buffalo (Bubalus bubalis). Pathogens 2020, 9, 280. https://doi.org/10.3390/pathogens9040280
Obregón D, Corona-González B, Díaz-Sánchez AA, Armas Y, Roque E, de Sena Oliveira MC, Cabezas-Cruz A. Efficient Transovarial Transmission of Babesia Spp. in Rhipicephalus microplus Ticks Fed on Water Buffalo (Bubalus bubalis). Pathogens. 2020; 9(4):280. https://doi.org/10.3390/pathogens9040280
Chicago/Turabian StyleObregón, Dasiel, Belkis Corona-González, Adrian Alberto Díaz-Sánchez, Yasmani Armas, Eugenio Roque, Márcia Cristina de Sena Oliveira, and Alejandro Cabezas-Cruz. 2020. "Efficient Transovarial Transmission of Babesia Spp. in Rhipicephalus microplus Ticks Fed on Water Buffalo (Bubalus bubalis)" Pathogens 9, no. 4: 280. https://doi.org/10.3390/pathogens9040280
APA StyleObregón, D., Corona-González, B., Díaz-Sánchez, A. A., Armas, Y., Roque, E., de Sena Oliveira, M. C., & Cabezas-Cruz, A. (2020). Efficient Transovarial Transmission of Babesia Spp. in Rhipicephalus microplus Ticks Fed on Water Buffalo (Bubalus bubalis). Pathogens, 9(4), 280. https://doi.org/10.3390/pathogens9040280