Resistance and Virulence Features of Bacteroides spp. Isolated from Abdominal Infections in Romanian Patients

 ,
, 
 ,
, 
 ,
, 

Abstract
1. Introduction
2. Results
2.1. Etiology of Intra-Abdominal Infections
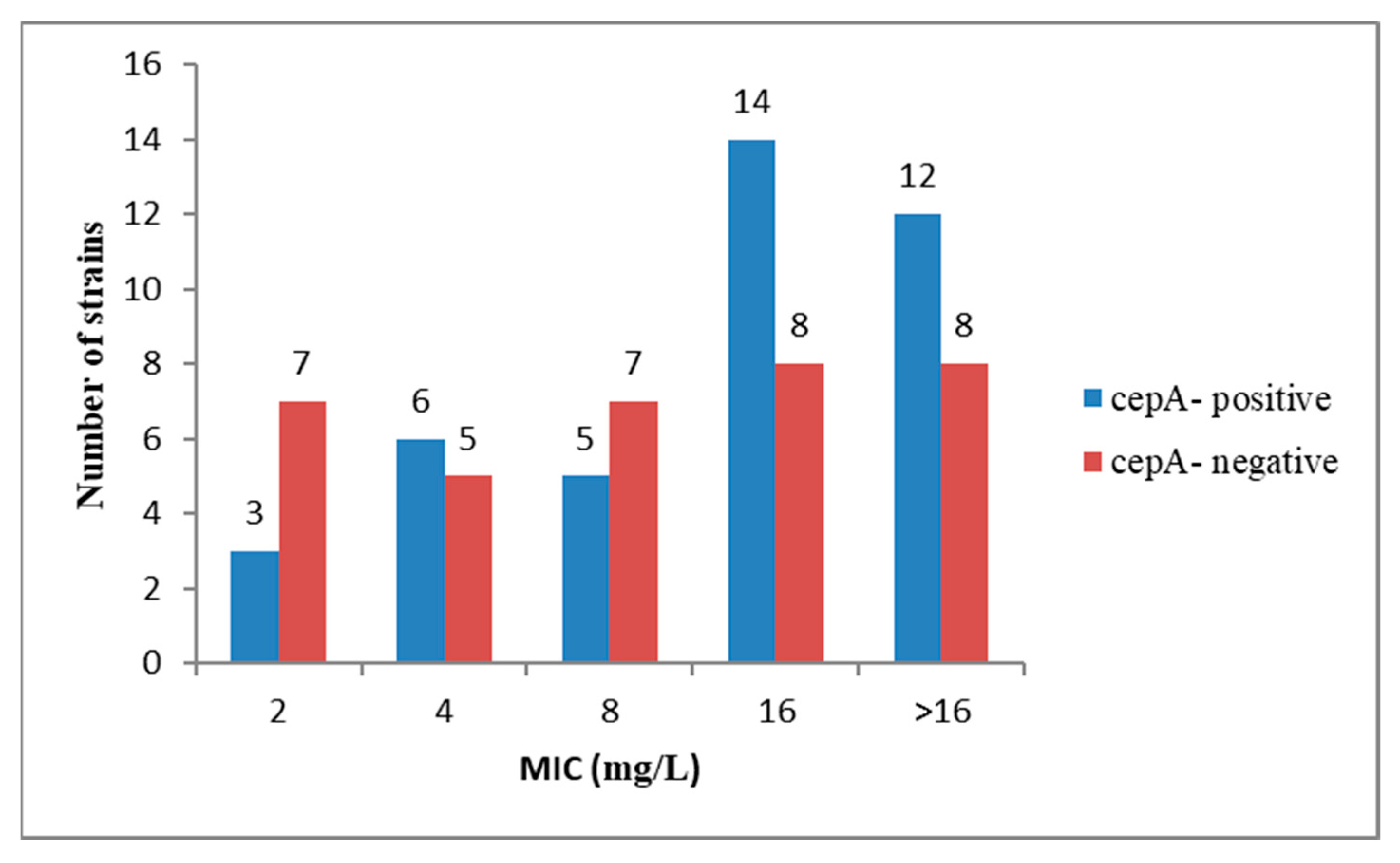
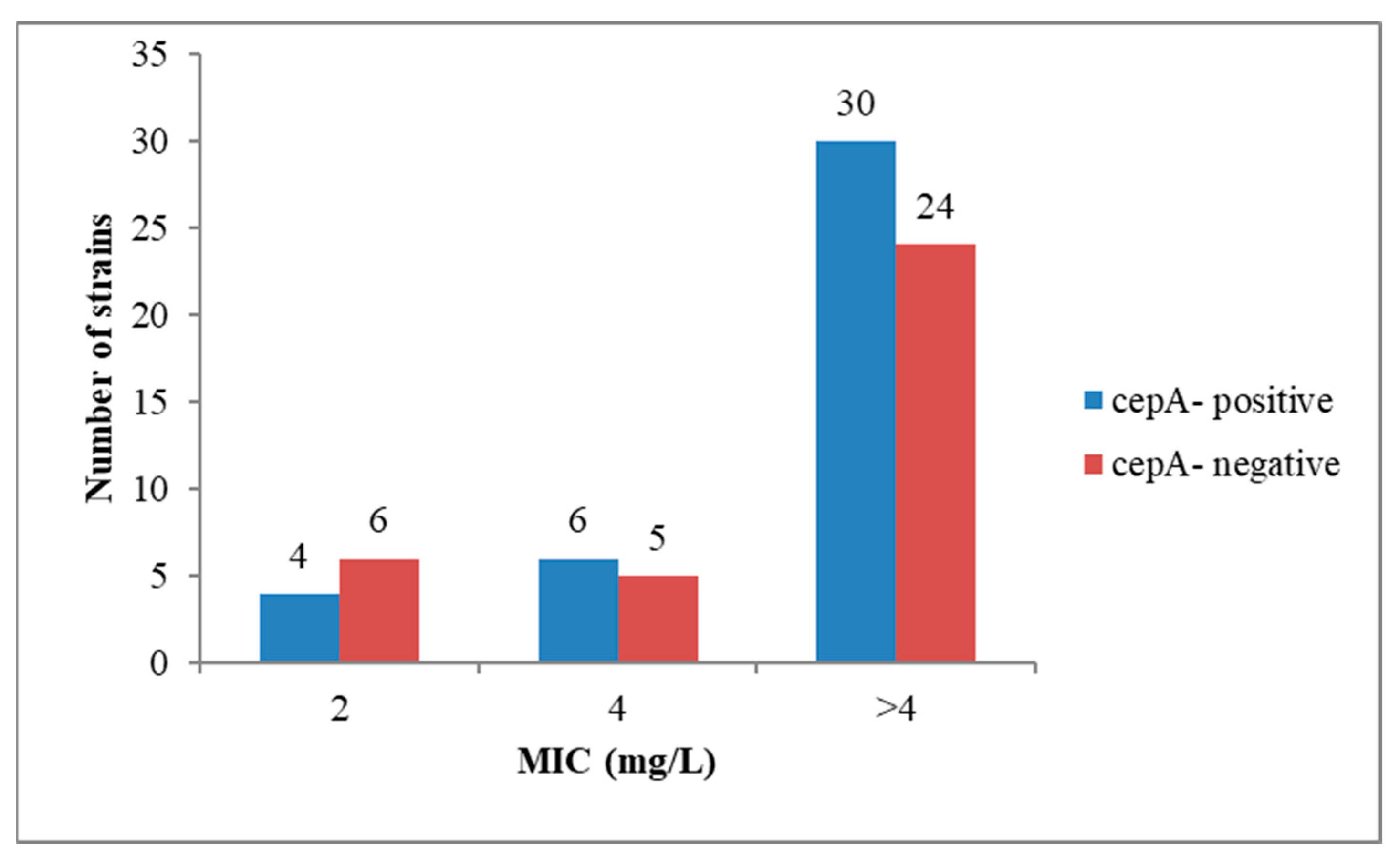
2.2. Antibiotic Resistance Profiles
2.3. Distribution of Resistance Genes among Bacteroides Isolates
2.4. Prevalence of Virulence Markers (VM) among Bacteroides spp.
3. Discussion
4. Materials and Methods
4.1. Anaerobic Bacterial Strains Isolation and Identification
4.2. Antibiotic Susceptibility Profiles
4.3. Detection of the Antibiotic Resistance and Virulence Markers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Brook, I. Indigenous Microbial Flora of Humans, Surgica Infections Diseases, 3rd ed.; Appleton and Lange: Norwalk, CT, USA, 1995; pp. 37–46. [Google Scholar]
- Jorgensen, J.H.; Carrol, K.C.; Funke, G.; Pfaller, M.A. Manual of Clinical Microbiology; ASM Press: Washington, DC, USA, 2015; Volume 1, Chapter 54; pp. 967–985. [Google Scholar]
- Noor, A.; Khetarpal, S. Anaerobic Infections. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. Available online: https://www.ncbi.nlm.nih.gov/books/NBK482349/ (accessed on 20 August 2020).
- Aldridge, K.E.; Ashcraft, D.; Cambre, K.; Pierson, C.L.; Jenkins, S.G.; Rosenblatt, J.E. Multicenter Survey of the Changing In Vitro Antimicrobial Susceptibilities of Clinical Isolates of Bacteroides fragilis Group, Prevotella, Fusobacterium, Porphyromonas, and Peptostreptococcus Species. Antimicrob. Agents Chemother. 2001, 45, 1238–1243. [Google Scholar] [CrossRef]
- Nagy, E.; Urbán, E.; Nord, C.E.; ESCMID Study Group on Antimicrobial Resistance in Anaerobic Bacteria. Antimicrobial susceptibility of Bacteroides fragilis group isolates in Europe: 20 years of experience. Clin. Microbiol. Infect. 2011, 17, 371–379. [Google Scholar] [CrossRef]
- Karlowsky, J.A.; Walkty, A.J.; Adam, H.J.; Baxter, M.R.; Hoban, D.J.; Zhanel, G.G. Prevalence of Antimicrobial Resistance among Clinical Isolates of Bacteroides fragilis Group in Canada in 2010–2011: CANWARD Surveillance Study. Antimicrob. Agents Chemother. 2012, 56, 1247–1252. [Google Scholar] [CrossRef]
- Eitel, Z.; Sóki, J.; Urbán, E.; Nagy, E. The prevalence of antibiotic resistance genes in Bacteroides fragilis group strains isolated in different European countries. Anaerobe 2013, 21, 43–49. [Google Scholar] [CrossRef]
- Píriz, S.; Vadillo, S.; Quesada, A.; Criado, J.; Cerrato, R.; Ayala, J.A. Relationship between penicillin-binding protein patterns and β-lactamases in clinical isolates of Bacteroides fragilis with different susceptibility to β-lactam antibiotics. J. Med. Microbiol. 2004, 53, 213–221. [Google Scholar] [CrossRef]
- Ghotaslou, R.; Yekani, M.; Memar, M.Y. The role of efflux pumps in Bacteroides fragilis resistance to antibiotics. Microbiol. Res. 2018, 210, 1–5. [Google Scholar] [CrossRef]
- Kierzkowska, M.; Majewska, A.; Szymanek-Majchrzak, K.; Sawicka-Grzelak, A.; Mlynarczyk, A.; Młynarczyk, G. In vitro effect of clindamycin against Bacteroides and Parabacteroides isolates in Poland. J. Glob. Antimicrob. Resist. 2018, 13, 49–52. [Google Scholar] [CrossRef]
- Johnsen, B.O.; Handal, N.; Meisal, R.; Bjørnholt, J.V.; Gaustad, P.; Leegaard, T.M. erm gene distribution among Norwegian Bacteroides isolates and evaluation of phenotypic tests to detect inducible clindamycin resistance in Bacteroides species. Anaerobe 2017, 47, 226–232. [Google Scholar] [CrossRef]
- Meggersee, R.; Abratt, V.R. The occurrence of antibiotic resistance genes in drug resistant Bacteroides fragilis isolates from Groote Schuur Hospital, South Africa. Anaerobe 2015, 32, 1–6. [Google Scholar] [CrossRef]
- Székely, E.; Eitel, Z.; Molnár, S.; Szász, I. Éva; Bilca, D.; Sóki, J. Analysis of Romanian Bacteroides isolates for antibiotic resistance levels and the corresponding antibiotic resistance genes. Anaerobe 2015, 31, 11–14. [Google Scholar] [CrossRef]
- Cox, G.M.; Harrison, T.S.; McDade, H.C.; Taborda, C.P.; Heinrich, G.; Casadevall, A.; Perfect, J.R. Superoxide Dismutase Influences the Virulence of Cryptococcus neoformans by Affecting Growth within Macrophages. Infect. Immun. 2003, 71, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Sarvari, K.P.; Soki, J.; Kristof, K.; Juhasz, E.; Miszti, C.; Latkoczy, K.; Melegh, S.Z.; Urban, E. A multicenter survey of the antibiotic susceptibility of clinical Bacteroides species from Hungary. Infect. Dis. 2018, 50, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Jeverica, S.; Kolenc, U.; Mueller-Premru, M.; Papst, L. Evaluation of the routine antimicrobial susceptibility testing results of clinically significant anaerobic bacteria in a Slovenian tertiary-care hospital in 2015. Anaerobe 2017, 47, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Avila-Campos, M.J.; Liu, C.; Song, Y.; Rowlinson, M.-C.; Finegold, S.M. Determination of bft Gene Subtypes in Bacteroides fragilis Clinical Isolates. J. Clin. Microbiol. 2007, 45, 1336–1338. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sarvari, K.P.; Sóki, J.; Iván, M.; Miszti, C.; Latkóczy, K.; Melegh, S.; Urbán, E. Detection of enterotoxin and protease genes among Hungarian clinical Bacteroides fragilis isolates. Anaerobe 2017, 48, 98–102. [Google Scholar] [CrossRef]
- Łuczak, M.; Obuch-Woszczatyński, P.; Pituch, H.; Leszczyński, P.; Martirosian, G.; Patrick, S.; Poxton, I.; Wintermans, R.G.; Dubreuil, L.; Meisel-Mikołajczyk, F. Search for enterotoxin gene in Bacteroides fragilis strains isolated from clinical specimens in Poland, Great Britain, The Netherlands and France. Med. Sci. Monit. 2001, 7, 222–225. [Google Scholar]
- Kierzkowska, M.; Majewska, A.; Szymanek-Majchrzak, K.; Sawicka-Grzelak, A.; Mlynarczyk, A.; Mlynarczyk, G. The presence of antibiotic resistance genes and bft genes as well as antibiotic susceptibility testing of Bacteroides fragilis strains isolated from inpatients of the Infant Jesus Teaching Hospital, Warsaw during 2007–2012. Anaerobe 2019, 56, 109–115. [Google Scholar] [CrossRef]
- Toprak, N.U.; Yagci, A.; Gulluoglu, B.M.; Akin, M.L.; Demirkalem, P.; Celenk, T.; Soyletir, G. A possible role of Bacteroides fragilis enterotoxin in the aetiology of colorectal cancer. Clin. Microbiol. Infect. 2006, 12, 782–786. [Google Scholar] [CrossRef]
- Parker, A.C.; Smith, C.J. Genetic and biochemical analysis of a novel Ambler class A beta-lactamase responsible for cefoxitin resistance in Bacteroides species. Antimicrob. Agents Chemother. 1993, 37, 1028–1036. [Google Scholar] [CrossRef]
- Oancea, S.; Stoia, M. Antibiotic resistance of bacterial pathogens: The magnitude of the problem from two perspectives—Romanian and worldwide. Rom. Biotechnol. Lett. 2010, 15, 5519e29. [Google Scholar]
- Almahdawy, O.T.; Pricop, R.; Sadik, O.; Najee, H.; Pircalabioru, G.G.; Marutescu, L.; Barbu, I.C.; Banu, O.; Cristea, V.; Grigore, R.; et al. Description of vancomycin resistance genes in Enterococcus sp. clinical strains isolated from Bucharest, Romania. Rom. Biotechnol. Lett. 2019, 24, 395–399. [Google Scholar] [CrossRef]
- Iancu, A.V.; Chifiriuc, M.C.; Tutunaru, D.; Arbune, M.; Gurău, G.; Coman, G.; Lazăr, V. Etiological investigation and antibiotic resistance profiles encountered in systemic bacterial infections. Biointerface Res. Appl. Chem. 2013, 3, 4. [Google Scholar]
- Ducu, R.; Gheorghe, I.; Chifiriuc, M.C.; Mihăescu, G.; Sârbu, I. Prevalence of vancomycin resistance phenotypes among enterococcus species isolated from clinical samples in a romanian hospital. Biointerface Res. Appl. Chem. 2019, 9, 4699–4704. [Google Scholar]
- Brukner, I.; Oughton, M.T. A Fundamental Change in Antibiotic Susceptibility Testing Would Better Prevent Therapeutic Failure: From Individual to Population-Based Analysis. Front. Microbiol. 2020, 11, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Maxson, T.; Blancett, C.D.; Graham, A.S.; Stefan, C.P.; Minogue, T.D. Rapid antibiotic susceptibility testing from blood culture bottles with species agnostic real-time polymerase chain reaction. PLoS ONE 2018, 13, e0209042. [Google Scholar] [CrossRef]
- Rolain, J.M.; Mallet, M.N.; Fournier, P.E.; Raoult, D. Real-time PCR for universal antibiotic susceptibility testing. J. Antimicrob. Chemother. 2004, 54, 538–541. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Group | Number | % |
|---|---|---|
| Gram-negative bacilli | 173 | 46 |
| Escherichia coli | 140 | 37 |
| Klebsiella pneumoniae | 11 | 3 |
| Hafnia alvei | 13 | 3 |
| Proteus mirabilis | 6 | 2 |
| Morganella morganii | 3 | 1 |
| Gram-positive cocci | 50 | 14 |
| Enterococcus faecium | 24 | 7 |
| Enterococcus faecalis | 9 | 2 |
| Enterococcus durans | 5 | 1 |
| Streptococcus anginosus | 6 | 2 |
| Streptococcus constellatus | 6 | 2 |
| Group | Number | % |
|---|---|---|
| Gram-negative rods | 94 | 25 |
| B. fragilis group | 75 | 20 |
| Parabacteroides | 7 | 2 |
| Fusobacterium | 4 | 1 |
| Megamonas | 5 | 1 |
| Prevotella | 2 | 0.5 |
| Dialister | 1 | 0.2 |
| Gram-positive rods | 44 | 12 |
| Clostridium | 31 | 8.2 |
| Eggerthella | 2 | 0.5 |
| Cutibacterium | 11 | 2.9 |
| Gram-positive cocci | 12 | 3 |
| Finegoldia magna | 7 | 1.8 |
| Parvimonas | 5 | 1.3 |
| Gram-negative cocci | 1 | 0.2 |
| Veionella | 1 | 0.2 |
| Species | Gene | ||||||
|---|---|---|---|---|---|---|---|
| (no.)% | |||||||
| cepA | erm F | tet Q | bft-1 | sod | katB | ftn | |
| B. fragilis (26) | 17 (65.3) | 4 (15.3) | 26 (100) | 15 (57.6) | 9 (34.6) | 7 (26.9) | 7 (30.4) |
| Peritoneal liquid | 10 (32) | 1 (3.2) | 12 (38.7) | 6 (19.3) | 6 (19.3) | 2 (6.4) | 2 (6.4) |
| Intra-abdominal abscess | 1 (33.3) | 1 (33.3) | 3 (100) | 2 (66.6) | 0 | 2 (66.6) | 2 (66.6) |
| Appendicular tissue | 2 (11.1) | 0 | 3 (16.6) | 2 (11.1) | 1 (5.5) | 1 (5.5) | 0 |
| Gallbladder secretion | 3 (37.5) | 0 | 3 (37.5) | 2 (25) | 1 (12.5) | 0 | 0 |
| Ascites fluid | 1 (11.1) | 2 (22.2) | 4 (44.4) | 3 (33.3) | 1 (11.1) | 2 (22.2) | 2 (22.2) |
| Perforated ulcer-biopsy tissue | 0 | 0 | 1 (25) | 0 | 0 | 0 | 1 (25) |
| B. vulgatus (16) | 10 (62.5) | 4 (25) | 15 (93.7) | 0 (0) | 2 (12.5) | 0 (0) | 0 (0) |
| Peritoneal liquid | 3 (9.6) | 1 (3.2) | 5 (16.1) | 0 | 2 (6.4) | 0 | 0 |
| Intra-abdominal abscess | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Appendicular tissue | 3 (16.6) | 0 | 3 (16.6) | 0 | 0 | 0 | 0 |
| Gallbladder secretion | 2 (25) | 3 (37.5) | 4 (50) | 0 | 0 | 0 | 0 |
| Ascites fluid | 1 (11.1) | 0 | 1 (11.1) | 0 | 0 | 0 | 0 |
| Perforated ulcer-biopsy tissue | 1 (25) | 0 | 2 (25) | 0 | 0 | 0 | 0 |
| B. uniformis (12) | 6 (50) | 1 (8.3) | 12 (100) | 0 (0) | 2 (16.6) | 0 (0) | 3 (25) |
| Peritoneal liquid | 0 | 0 | 3 (9.6) | 0 | 0 | 0 | 1 (3.2) |
| Intra-abdominal abscess | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Appendicular tissue | 2 (11.1) | 1 (5.5) | 5 (27.7) | 0 | 0 | 0 | 2 (11.1) |
| Gallbladder secretion | 1 (12.5) | 0 | 1 (12.5) | 0 | 0 | 0 | 0 |
| Ascites fluid | 1 (11.1) | 0 | 2 (22.2) | 0 | 2 (22.2) | 0 | 0 |
| Perforated ulcer-biopsy tissue | 2 (25) | 0 | 1 (25) | 0 | 0 | 0 | 0 |
| B. ovatus (7) | 1 (14.2) | 1 (14.2) | 7 (100) | 0 (0) | 1 (14.2) | 0 (0) | 2 (28.5) |
| Peritoneal liquid | 1 (3.2) | 1 (3.2) | 4 (13) | 0 | 0 | 0 | 2 (6.4) |
| Intra-abdominal abscess | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Appendicular tissue | 0 | 0 | 3 (16.6) | 0 | 1 (5.5) | 0 | 0 |
| Gallbladder secretion | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Ascites fluid | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Perforated ulcer-biopsy tissue | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B. thetaiotaomicron (8) | 4 (50) | 3 (37.5) | 8 (100) | 0 (0) | 5 (62.8) | 1 (12.5) | 1 (12.5) |
| Peritoneal liquid | 2 (6.4) | 2 (6.4) | 3 (9.6) | 0 | 2 (6.4) | 0 | 0 |
| Intra-abdominal abscess | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Appendicular tissue | 2 (11.1) | 1 (5.5) | 4 (22.2) | 0 | 2 (11.1) | 1 (5.5) | 1 (5.5) |
| Gallbladder secretion | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Ascites fluid | 0 | 0 | 1 (11.1) | 0 | 1 (11.1) | 0 | 0 |
| Perforated ulcer-biopsy tissue | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B. caccae (1) | 1 (100) | 0 (0) | 1 (100) | 0 (0) | 1 (100) | 0 (0) | 0 (0) |
| Peritoneal liquid | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Intra-abdominal abscess | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Appendicular tissue | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Gallbladder secretion | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Ascites fluid | 1 (11.1) | 0 | 1 (11.1) | 0 | 1 (11.1) | 0 | 0 |
| Perforated ulcer-biopsy tissue | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B. stercoris (5) | 1 (20) | 1 (20) | 5 (100) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| Peritoneal liquid | 1 (3.2) | 1 (3.2) | 4 (13) | 0 | 0 | 0 | 0 |
| Intra-abdominal abscess | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Appendicular tissue | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Gallbladder secretion | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Ascites fluid | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Perforated ulcer-biopsy tissue | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Total (no.)% | 40 (53.3) | 14 (18.6) | 74 (98.6) | 15 (57.6) | 20 (26.6) | 8 (10.6) | 13 (17.3) |
| Gene | Primer | Size | Ref. |
|---|---|---|---|
| bft-1 | 5-GAGCCGAAGACGGTGTATGTGATTTGT-3 5-TGCTCAGCGCCCAGTATATGACCTAGT-3 | 500 bp | [21] |
| katB | 5-GTAGCAGGAGAACGCGGAGCTGCT-3 5-GTTCATCCGCAGGCATCAGTCGGA-3 | 170 bp | GenBank |
| sod | 5-ACAATGCGCTGGAACCTGTA-3 5-TTTCGAAGGTTTCGGAGCGA-3 | 230 bp | GenBank |
| ftn | 5-ACG TTTCAGCGGTTTTGCAC-3 5-CGTTCGTGCTCAAAGACGTG-3 | 183 bp | GenBank |
| cepA | 5-CGCAATGCCAAAGGACAACA-3 5-ACGATACGTGAGATGTCCGC-3 | 779 bp | GenBank |
| tet Q | 5-CTGTTTGCCAGTGGAGCAAC-3 5-AGCAACTTTGTCTGCGCTTG-3 | 460 bp | GenBank |
| cfxA | 5 -GCTCAAACAGATAGTTTTAT-3 5 -GAGCTCACAATGATGTTGCC-3 | 802 bp | [22] |
| erm F | 5-AGGTGCAGGGAAAGGTCATT-3 5-ACCTCTGCCATTAACAGCAAT-3 | 446 bp | GenBank |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pricop, G.R.; Gheorghe, I.; Pircalabioru, G.G.; Cristea, V.; Popa, M.; Marutescu, L.; Chifiriuc, M.C.; Mihaescu, G.; Bezirtzoglou, E. Resistance and Virulence Features of Bacteroides spp. Isolated from Abdominal Infections in Romanian Patients. Pathogens 2020, 9, 940. https://doi.org/10.3390/pathogens9110940
Pricop GR, Gheorghe I, Pircalabioru GG, Cristea V, Popa M, Marutescu L, Chifiriuc MC, Mihaescu G, Bezirtzoglou E. Resistance and Virulence Features of Bacteroides spp. Isolated from Abdominal Infections in Romanian Patients. Pathogens. 2020; 9(11):940. https://doi.org/10.3390/pathogens9110940
Chicago/Turabian StylePricop, Gabriela Roxana, Irina Gheorghe, Gratiela Gradisteanu Pircalabioru, Violeta Cristea, Marcela Popa, Luminita Marutescu, Mariana Carmen Chifiriuc, Grigore Mihaescu, and Eugenia Bezirtzoglou. 2020. "Resistance and Virulence Features of Bacteroides spp. Isolated from Abdominal Infections in Romanian Patients" Pathogens 9, no. 11: 940. https://doi.org/10.3390/pathogens9110940
APA StylePricop, G. R., Gheorghe, I., Pircalabioru, G. G., Cristea, V., Popa, M., Marutescu, L., Chifiriuc, M. C., Mihaescu, G., & Bezirtzoglou, E. (2020). Resistance and Virulence Features of Bacteroides spp. Isolated from Abdominal Infections in Romanian Patients. Pathogens, 9(11), 940. https://doi.org/10.3390/pathogens9110940