Identification of Clinically Relevant Streptococcus and Enterococcus Species Based on Biochemical Methods and 16S rRNA, sodA, tuf, rpoB, and recA Gene Sequencing

 ,
,  ,
, 
Abstract
1. Introduction
2. Results
2.1. Serotyping and Identification of Gram-Positive Cocci with the Vitek® 2 System and MALDI-TOF MS
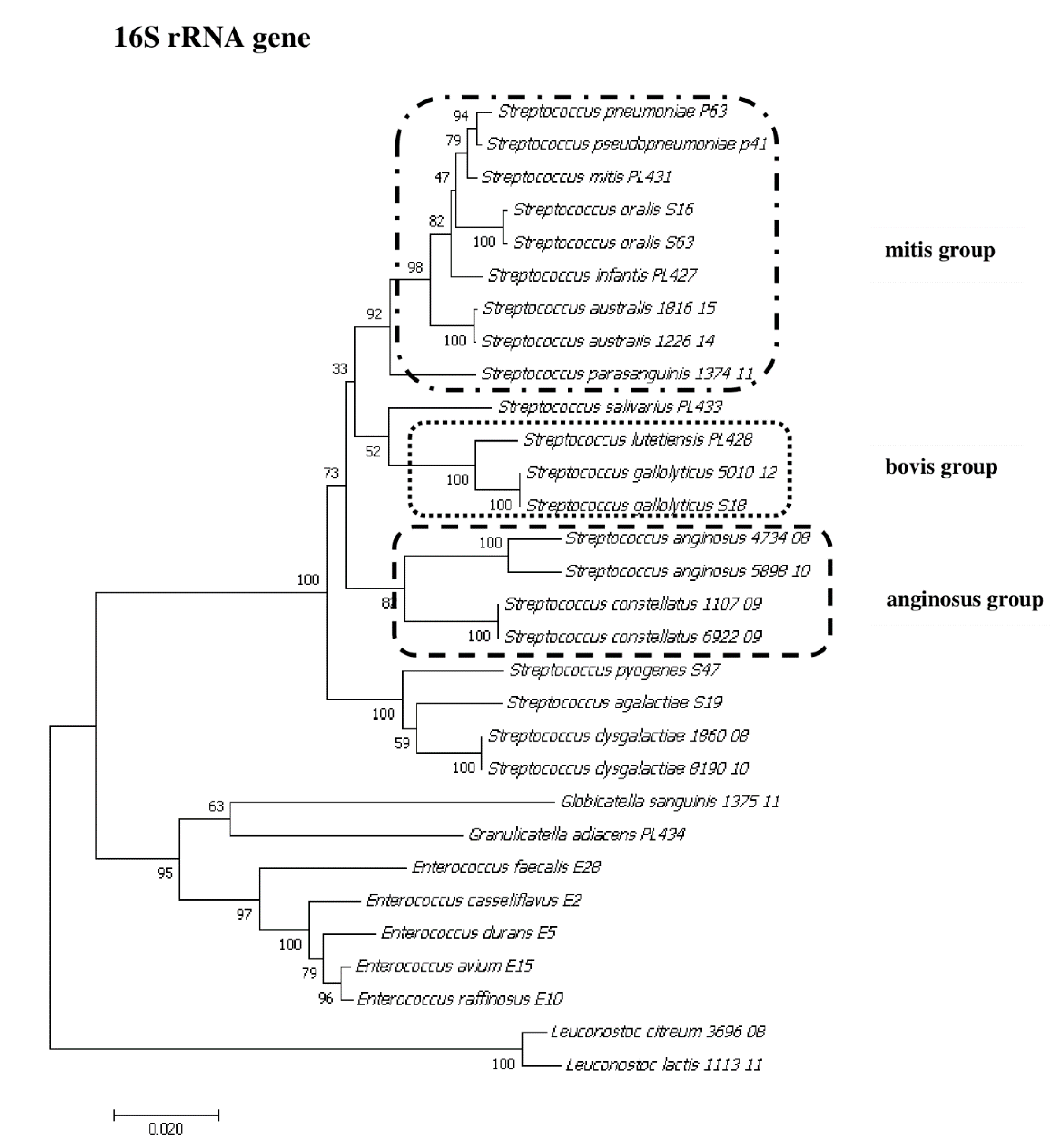
2.2. Sanger Sequencing of the 16S rRNA Gene
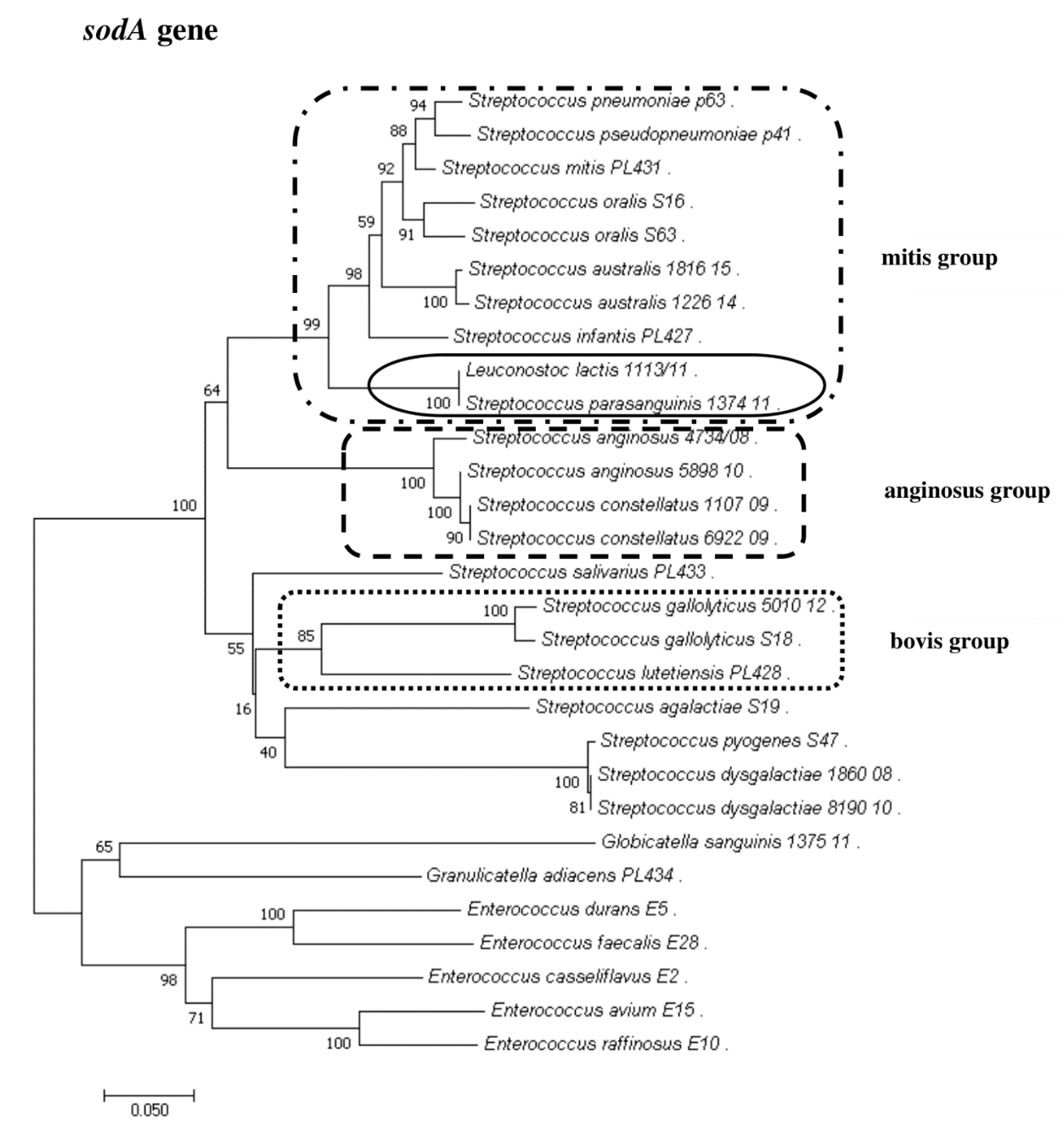
2.3. Sanger Sequencing of the sodA Gene
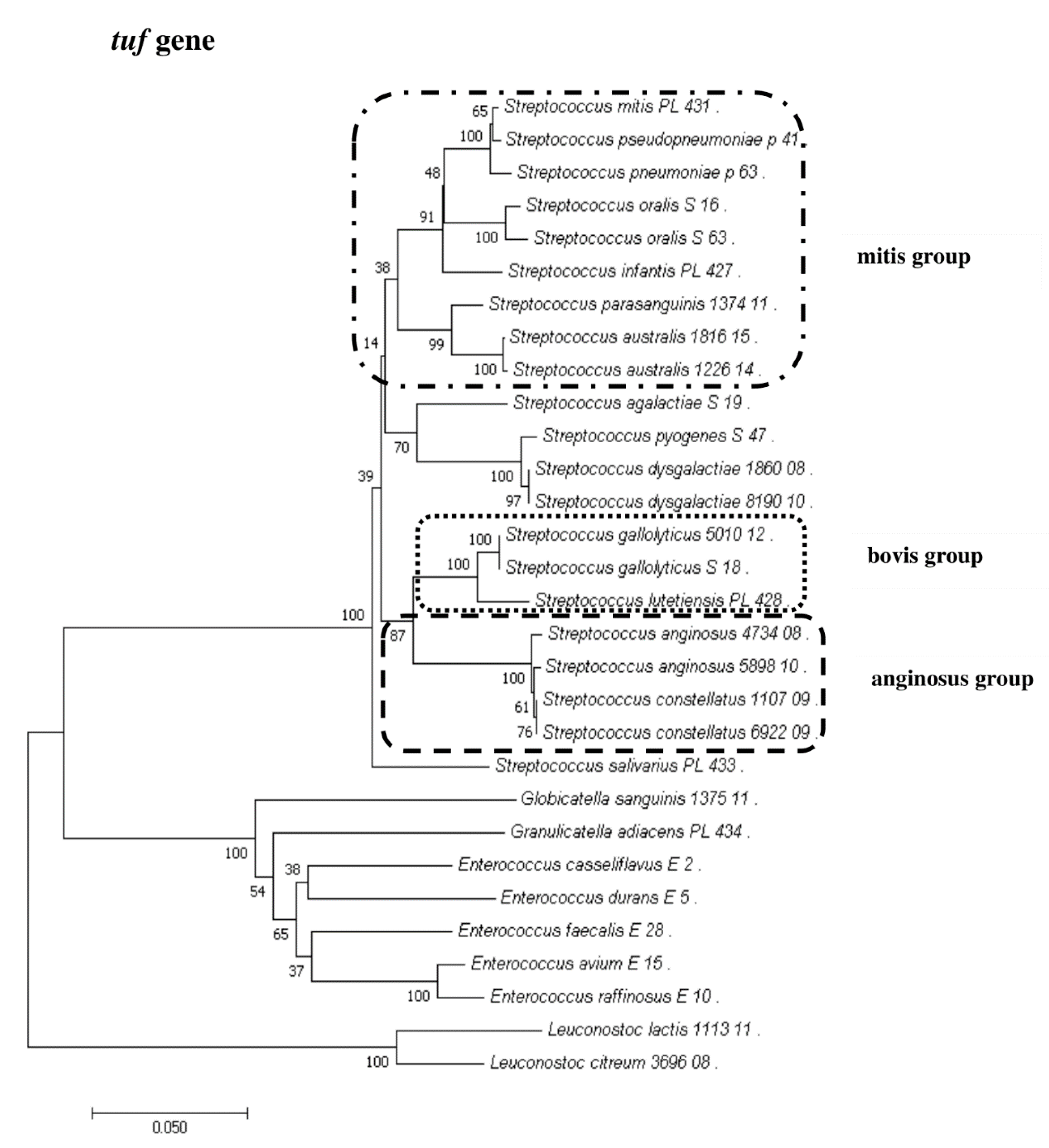
2.4. Sanger Sequencing of the tuf Gene
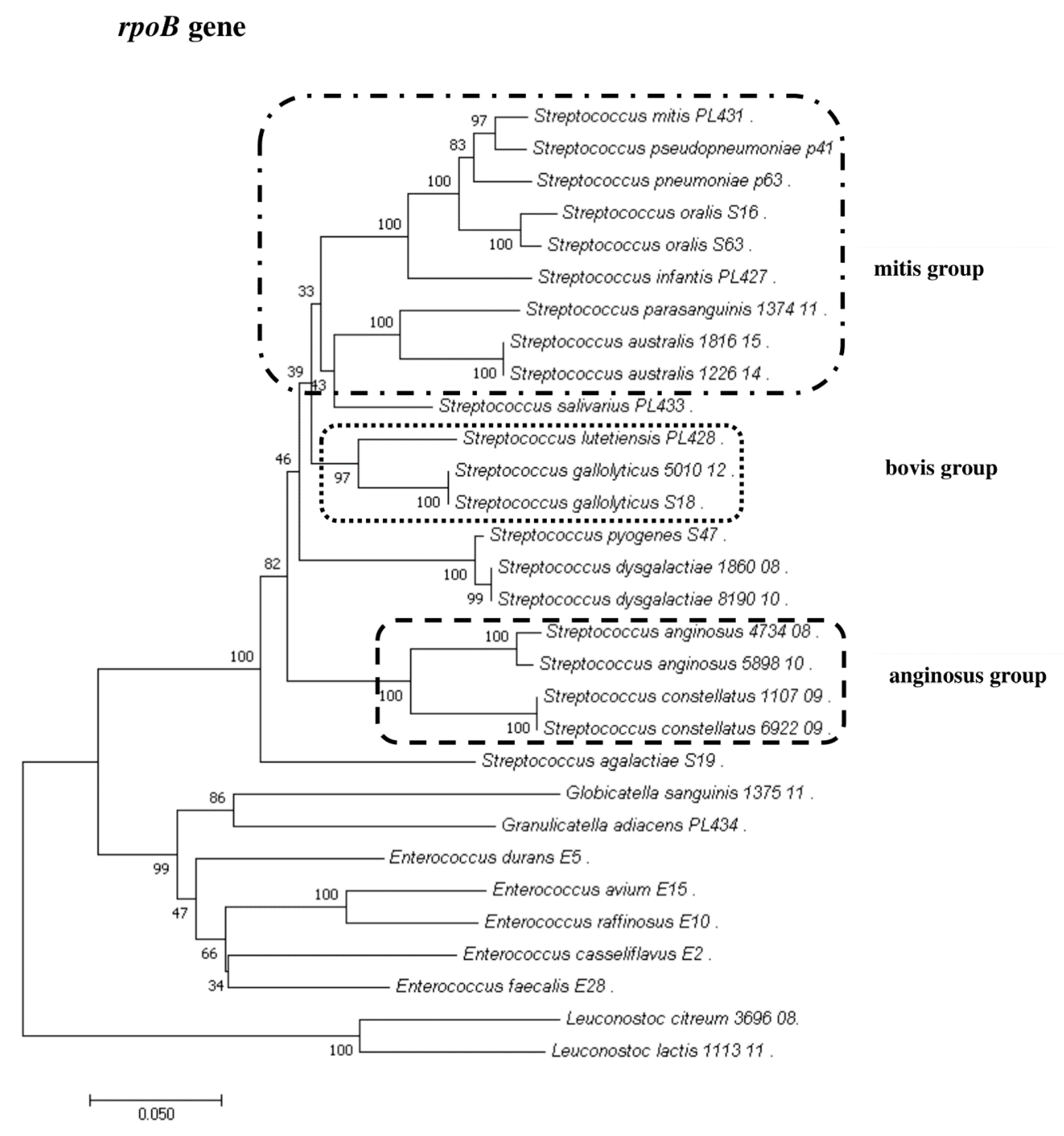
2.5. Sanger Sequencing of the rpoB Gene
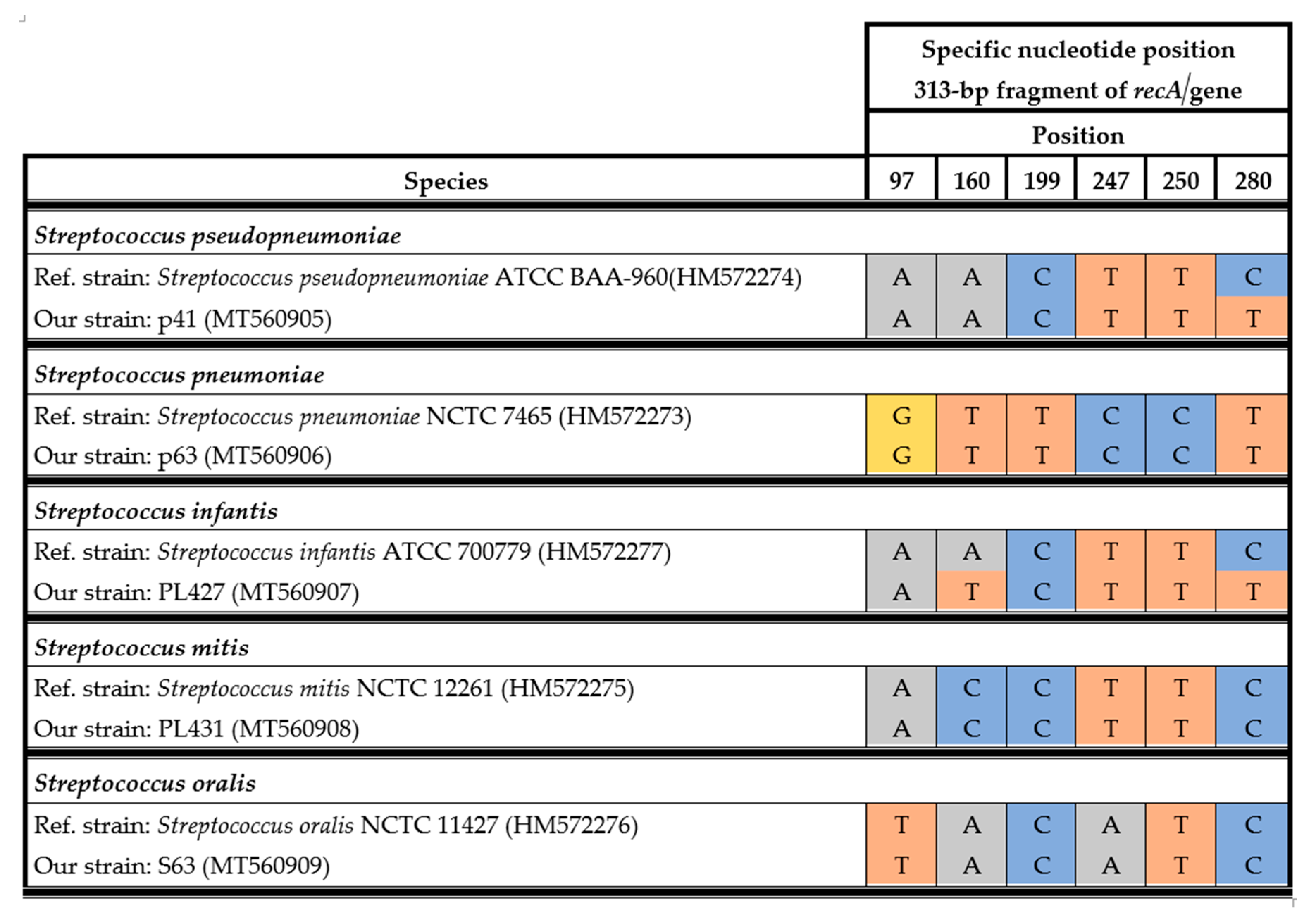
2.6. Analysis of the recA Gene for the Streptococcal mitis Group
2.7. Comparison of the Sequencing Methods
2.8. Phylogenetic Analysis of Streptococcus and Enterococcus
3. Discussion
4. Materials and Methods
4.1. Ethical Approval
4.2. Bacterial Isolates
4.3. Phenotypic Identification Tests
4.4. MALDI-TOF MS Identification
4.5. Genomic DNA Extraction
4.6. PCR Amplification of the 16S rRNA, sodA, rpoB tuf, and recA Genes
4.7. Sanger Sequencing Analysis of the 16S rRNA, sodA, rpoB, and tuf Genes
4.8. Sanger Sequencing Analysis of the recA Gene
4.9. Nucleotide Sequence Accession Numbers
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lal, D.; Verma, M.; Lal, R. Exploring internal features of 16S rRNA gene for identification of clinically relevant species of the genus Streptococcus. Ann. Clin. Microbiol. Antimicrob. 2011, 10, 28. [Google Scholar] [CrossRef] [PubMed]
- Kohler, W. The present state of species within the genera Streptococcus and Enterococcus. Int. J. Med. Microbiol. 2007, 297, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.Y.; Zhi, X.Y.; Li, H.W.; Klenk, H.P.; Li, W.J. Comparative genomics of the bacterial genus Streptococcus illuminates evolutionary implications of species groups. PLoS ONE 2014, 9, e101229. [Google Scholar] [CrossRef] [PubMed]
- Farley, M.M.; Strasbaugh, L.J. Group B streptococcal disease in nonpregnant adults. Clin. Infect. Dis. 2001, 33, 556–561. [Google Scholar] [CrossRef]
- Henriques-Normark, B.; Tuomanen, E.I. The pneumococcus: Epidemiology, microbiology, and pathogenesis. Cold Spring Harb. Perspect. Med. 2013, 3, a010215. [Google Scholar] [CrossRef] [PubMed]
- Pekuz, S.; Soysal, A.; Akkoc, G.; Atıcı, S.; Yakut, N.; Gelmez, G.A.; Kadayifci, E.K.; Güneser, D.; Demir, S.O.; Söyletir, G.; et al. Prevalence of nasopharyngeal carriage, serotype distribution, and antimicrobial resistance of Streptococcus pneumoniae among children with chronic diseases. Jpn. J. Infect. Dis. 2019, 72, 7–13. [Google Scholar] [CrossRef]
- Gerhardt, P.; Murray, R.G.E.; Willis, A.; Krieg, N.R. Methods for General and Molecular Bacteriology; American Society for Microbiology: Washington, DC, USA, 1994; pp. 619–647. [Google Scholar]
- Kawamura, Y.; Hou, X.G.; Sultana, F.; Miura, H.; Ezaki, T. Determination of 16S rRNA sequences of Streptococcus mitis and Streptococcus gordonii and phylogenetic relationships among members of the genus Streptococcus. Int. J. Syst. Bacteriol. 1995, 45, 406–408. [Google Scholar] [CrossRef]
- Facklam, R.; Elliott, J.A. Identification, classification, and clinical relevance of catalase-negative, gram-positive cocci, excluding the streptococci and enterococci. Clin. Microbiol. Rev. 1995, 8, 479–495. [Google Scholar] [CrossRef]
- Parks, T.; Barrett, L.; Jones, N. Invasive streptococcal disease: A review for clinicians. Br. Med. Bull. 2015, 115, 77–89. [Google Scholar] [CrossRef]
- Schleifer, K.H.; Kilpper-Balz, R. Transfer of Streptococcus faecalis and Streptococcus faecium to the genus Enterococcus nom. rev. as Enterococcus faecalis comb. nov. and Enterococcus faecium comb. nov. Int. J. Syst. Evol. 1984, 34, 31–34. [Google Scholar] [CrossRef]
- Devriese, L.; Baele, M.; Butaye, P. The Genus Enterococcus. In The Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2006; pp. 163–174. [Google Scholar] [CrossRef]
- Kosecka-Strojek, M.; Sabat, A.J.; Akkerboom, V.; Kooistra-Smid, A.M.D.M.; Miedzobrodzki, J.; Friedrich, A.W. Development of a reference data set for assigning Streptococcus and Enterococcus species based on next generation sequencing of the 16S-23S rRNA region. Antimicrob Resist. Infect. Control 2019, 8, 178. [Google Scholar] [CrossRef] [PubMed]
- Savini, V.; Franco, A.; Gherardi, G.; Marrollo, R.; Argentieri, A.V.; De Araujo, F.P.; Amoruso, R.; Battisti, A.; Fazii, P.; Carretto, E. Beta-hemolytic, multi-lancefield antigen-agglutinating Enterococcus durans from a pregnant woman, mimicking Streptococcus agalactiae. J. Clin. Microbiol. 2014, 52, 2181–2182. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ferede, Z.T.; Tullu, K.D.; Derese, S.G.; Yeshanew, A.G. Prevalence and antimicrobial susceptibility pattern of Enterococcus species isolated from different clinical samples at Black Lion Specialized Teaching Hospital, Addis Ababa, Ethiopia. BMC Res. Notes 2018, 11, 793. [Google Scholar] [CrossRef] [PubMed]
- Top, J.; Willems, R.; Bonten, M. Emergence of CC17 Enterococcus faecium: From commensal to hospital-adapted pathogen. FEMS Immunol. Med. Microbiol. 2008, 52, 297–308. [Google Scholar] [CrossRef]
- Fernández-Hidalgo, N.; Escolà-Vergé, L. Enterococcus faecalis Bacteremia: Consider an echocardiography, but consult an infectious diseases specialist. J. Am. Coll. Cardiol. 2019, 74, 202–204. [Google Scholar] [CrossRef]
- Ligozzi, M.; Bernini, C.; Bonora, M.G.; De Fatima, M.; Zuliani, J.; Fontana, R. Evaluation of the VITEK 2 system for identification and antimicrobial susceptibility testing of medically relevant gram-positive cocci. J. Clin. Microbiol. 2002, 40, 1681–1686. [Google Scholar] [CrossRef]
- Chen, J.H.; She, K.K.; Wong, O.Y.; Teng, J.L.; Yam, W.C.; Lau, S.K.; Woo, P.C.; Cheng, V.C.; Yuen, K.Y. Use of MALDI Biotyper plus ClinProTools mass spectra analysis for correct identification of Streptococcus pneumoniae and Streptococcus mitis/oralis. J. Clin. Pathol. 2015, 68, 652–656. [Google Scholar] [CrossRef]
- Singhal, N.; Kumar, M.; Kanaujia, P.K.; Virdi, J.S. MALDI-TOF mass spectrometry: An emerging technology for microbial identification and diagnosis. Front. Microbiol. 2015, 6, 791. [Google Scholar] [CrossRef]
- Hoshino, T.; Fujiwara, T.; Kilian, M. Use of phylogenetic and phenotypic analyses to identify nonhemolytic streptococci isolated from bacteremic patients. J. Clin. Microbiol. 2005, 43, 6073–6085. [Google Scholar] [CrossRef]
- Isaksson, J.; Rasmussen, M.; Nilson, B.; Stadler, L.S.; Kurland, S.; Olaison, L.; Ek, E.; Herrmann, B. Comparison of species identification of endocarditis associated viridans streptococci using rnpB genotyping and 2 MALDI-TOF systems. Diagn. Microbiol. Infect. Dis. 2015, 81, 240–245. [Google Scholar] [CrossRef]
- Clarridge, J.E. Impact of 16S rRNA gene sequence analysis for identification of bacteria on clinical microbiology and infectious diseases. Clin. Microbiol. Rev. 2004, 17, 840–862. [Google Scholar] [CrossRef] [PubMed]
- Poyart, C.; Quesne, G.; Coulon, S.; Berche, P.; Trieu-Cuot, P. Identification of streptococci to species level by sequencing the gene encoding the manganese-dependent superoxide dismutase. J. Clin. Microbiol. 1998, 36, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xing, J.; Li, B.; Wang, P.; Liu, J. Use of tuf as a target for sequence-based identification of Gram-positive cocci of the genus Enterococcus, Streptococcus, coagulase-negative Staphylococcus, and Lactococcus. Ann. Clin. Microbiol. Antimicrob. 2012, 11, 31. [Google Scholar] [CrossRef] [PubMed]
- Drancourt, M.; Roux, V.; Fournier, P.E.; Raoult, D. rpoB gene sequence-based identification of aerobic Gram-positive cocci of the genera Streptococcus, Enterococcus, Gemella, Abiotrophia, and Granulicatella. J. Clin. Microbiol. 2004, 42, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Jensen, A.; Scholz, C.F.P.; Kilian, M. Re-evaluation of the taxonomy of the Mitis group of the genus Streptococcus based on whole genome phylogenetic analyses, and proposed reclassification of Streptococcus dentisani as Streptococcus oralis subsp. dentisani comb. nov., Streptococcus tigurinus as Streptococcus oralis subsp. tigurinus comb. nov., and Streptococcus oligofermentans as a later synonym of Streptococcus cristatus. Int. J. Syst. Evol. Microbiol. 2016, 66, 4803–4820. [Google Scholar] [CrossRef]
- Sadowy, E.; Hryniewicz, W. Identification of Streptococcus pneumoniae and other Mitis streptococci: Importance of molecular methods. Eur. J. Clin. Microbiol. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Dekker, J.P.; Lau, A.F. An Update on the Streptococcus bovis Group: Classification, Identification, and Disease Associations. J. Clin. Microbiol. 2016, 54, 1694–1699. [Google Scholar] [CrossRef]
- Werno, A.M.; Christner, M.; Anderson, T.P.; Murdoch, D.R. Differentiation of Streptococcus pneumoniae from nonpneumococcal streptococci of the Streptococcus mitis group by Matrix-Assisted Laser Desorption Ionization–Time of Flight Mass Spectrometry. J. Clin. Microbiol. 2012, 50, 2863–2867. [Google Scholar] [CrossRef]
- Zbinden, A.; Köhler, N.; Bloemberg, G.V. recA-based PCR assay for accurate differentiation of Streptococcus pneumoniae from other viridans streptococci. J. Clin. Microbiol. 2011, 49, 523–527. [Google Scholar] [CrossRef]
- BIO-RAD. 2017. Manufacturer’s Instructions: Agglutination Test for the Grouping Streptococci Belonging to Groups A, B, C, D, F, G. PASTOREX™ STREP. 2017. Available online: https://www.biorad.com/webroot/web/pdf/inserts/CDG/en/61721_880999_EN.pdf (accessed on 6 November 2020).
- Gherardi, G.; Angeletti, S.; Panitti, M.; Pompilio, A.; Di Bonaventura, G.; Crea, F.; Avola, A.; Fico, L.; Palazzo, C.; Sapia, G.F.; et al. Comparative evaluation of the Vitek-2 Compact and Phoenix systems for rapid identification and antibiotic susceptibility testing directly from blood cultures of Gram-negative and Gram-positive isolates. Diagn. Microbiol. Infect. Dis. 2012, 72, 20–31. [Google Scholar] [CrossRef]
- de Cueto, M.; Ceballos, E.; Martinez-Martinez, L.; Perea, E.J.; Pascual, A. Use of positive blood cultures for direct identification and susceptibility testing with the Vitek 2 system. J. Clin. Microbiol. 2004, 42, 3734–3738. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lee, S.; Yang, B.; Lu, J. Rapid identification and susceptibility testing using the VITEK 2 system using culture fluids from positive BacT/ALERT blood cultures. J. Microbiol. Immunol. Infect. 2008, 41, 259–264. [Google Scholar] [PubMed]
- Romero-Gómez, M.P.; Gómez-Gil, R.; Paño-Pardo, J.R.; Mingorance, J. Identification and susceptibility testing of microorganism by direct inoculation from positive blood culture bottles by combining MALDI-TOF and Vitek-2 Compact is rapid and effective. J. Infect. 2012, 65, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Dubois, D.; Segonds, C.; Prere, M.F.; Marty, N.; Oswald, E. Identification of clinical Streptococcus pneumoniae isolates among other alpha and nonhemolytic streptococci by use of the Vitek MS matrix-assisted laser desorption ionization-time of flight mass spectrometry system. J. Clin. Microbiol. 2013, 51, 1861–1867. [Google Scholar] [CrossRef]
- Monteiro, J.; Inoue, F.M.; Lobo, A.P.; Sugawara, E.K.; Boaretti, F.M.; Tufik, S. Fast and reliable bacterial identification direct from positive blood culture using a new TFA sample preparation protocol and the Vitek® MS system. J. Microbiol. Methods 2015, 109, 157–159. [Google Scholar] [CrossRef]
- Febbraro, F.; Rodio, D.M.; Puggioni, G.; Antonelli, G.; Pietropaolo, V.; Trancassini, M. MALDI-TOF MS Versus VITEK®2: Comparison of systems for the identification of microorganisms responsible for bacteremia. Curr. Microbiol. 2016, 73, 843–850. [Google Scholar] [CrossRef][Green Version]
- Srinivasan, R.; Karaoz, U.; Volegova, M.; MacKichan, J.; Kato-Maeda, M.; Miller, S.; Nadarajan, R.; Brodie, E.L.; Lynch, S.V. Use of 16S rRNA Gene for identification of a broad range of clinically relevant bacterial pathogens. PLoS ONE 2015, 10, e0117617. [Google Scholar] [CrossRef]
- Janda, M.J.; Sharon, L.A. 16S rRNA gene sequencing for bacterial identification in the diagnostic laboratory: Pluses, Perils, and Pitfalls. J. Clin. Microbiol. 2007, 45, 2761–2764. [Google Scholar] [CrossRef]
- Summanen, P.H.; Rowlinson, M.; Wooton, J.; Finegold, S.M. Evaluation of genotypic and phenotypic methods for differentiation of the members of the Anginosus group streptococci. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 1123–1128. [Google Scholar] [CrossRef][Green Version]
- Stackebrandt, E.; Frederiksen, W.; Garrity, G.M.; Grimont, P.; Kämpfer, P.A.D.; Maiden, M.C.J.; Nesme, X.; Rosselló-Mora, R.; Swings, J.; Trüper, H.G.; et al. Report of the ad hoc committee for the re-evaluation of the species definition in bacteriology. Int. J. Syst. Evol. Microbiol. 2002, 52, 1043–1047. [Google Scholar] [CrossRef]
- Lerat, E.; Daubin, V.; Ochman, H.; Moran, N.A. Evolutionary origins of genomic repertoires in bacteria. PLoS Biol. 2005, 3, e130. [Google Scholar] [CrossRef] [PubMed]
- Leray, M.; Knowlton, N.; Ho, S.L.; Nguyen, B.N.; Machida, R.J. GenBank is a reliable resource for 21st century biodiversity research. Proc. Natl. Acad. Sci. USA 2019, 116, 22651–22656. [Google Scholar] [CrossRef] [PubMed]
- Zbinden, A.; Mueller, N.J.; Tarr, P.E.; Spröer, C.; Keller, P.M.; Bloemberg, G.V. Streptococcus tigurinus sp. nov., isolated from blood of patients with endocarditis, meningitis and spondylodiscitis. Int. J. Syst. Evol. Microbiol. 2012, 62, 2941–2945. [Google Scholar] [CrossRef] [PubMed]
- Poyart, C.; Quesne, G.; Trieu-Cuot, P. Taxonomic dissection of the Streptococcus bovis group by analysis of manganese-dependent superoxide dismutase gene (sodA) sequences: Reclassification of ‘Streptococcus infantarius subsp. coli’ as Streptococcus lutetiensis sp. nov. and of Streptococcus bovis biotype 11.2 as Streptococcus pasteurianus sp. nov. Int. J. Syst. Evol. Microbiol. 2002, 52, 1247–1255. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.R.; Fedorka-Cray, P.J.; Barrett, J.B. Use of a genus- and species-specific multiplex PCR for identification of enterococci. J. Clin. Microbiol. 2004, 42, 3558–3565. [Google Scholar] [CrossRef]
- Castillo-Rojas, G.; Mazari-Hiríart, M.; de León, S.P.; Amieva-Fernández, R.I.; Agis-Juárez, R.A.; Huebner, J.; López-Vidal, Y. Comparison of Enterococcus faecium and Enterococcus faecalis strains isolated from water and clinical samples: Antimicrobial susceptibility and genetic relationships. PLoS ONE 2013, 8, e59491. [Google Scholar] [CrossRef]
- Clarridge, J.E.; Osting, C.; Jalali, M.; Osborne, J.; Waddington, M. Genotypic and phenotypic characterization of “Streptococcus milleri” group isolates from a Veterans Administration hospital population. J. Clin. Microbiol. 1999, 37, 3681–3687. [Google Scholar] [CrossRef]
- Arinto-Garcia, R.; Pinho, M.D.; Carriço, J.A.; Melo-Cristino, J.; Ramirez, M. Comparing Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry and phenotypic and molecular methods for identification of species within the Streptococcus anginosus Group. J. Clin. Microbiol. 2015, 53, 3580–3588. [Google Scholar] [CrossRef]
- Limia, A.; Alarcón, T.; Jiménez, M.L.; López-Brea, M. Comparison of three methods for identification of Streptococcus milleri group isolates to species level. Eur. J. Clin. Microbiol. Infect. Dis. 2000, 19, 128–131. [Google Scholar] [CrossRef]
- Collins, M.D.; Aguirre, M.; Facklam, R.R.; Shallcross, J.; Williams, A.M. Globicatella sanguis gen.nov., sp.nov., a new gram-positive catalase-negative bacterium from human sources. J. Appl. Bacteriol. 1992, 73, 433–437. [Google Scholar] [CrossRef]
- Shewmaker, P.L.; Steigerwalt, A.G.; Shealey, L.; Weyant, R.; Facklam, R. DNA relatedness, phenotypic characteristics, and antimicrobial susceptibilities of Globicatella sanguinis strains. J. Clin. Microbiol. 2001, 39, 4052–4057. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.O.; Buckwalter, S.P.; Henry, M.W.; Wu, F.; Maloney, K.F.; Abraham, B.K.; Hartman, B.J.; Brause, B.D.; Whittier, S.; Walsh, T.J.; et al. Globicatella sanguinis Osteomyelitis and Bacteremia: Review of an emerging human pathogen with an expanding spectrum of disease. Open Forum Infect. Dis. 2017, 4, ofw277. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, Y.; Hou, X.G.; Sultana, F.; Liu, S.; Yamamoto, H.; Ezaki, T. Transfer of Streptococcus adjacens and Streptococcus defectivus to Abiotrophia gen. nov. as Abiotrophia adiacens comb. nov. and Abiotrophia defectiva comb. nov., respectively. Int. J. Syst. Bacteriol. 1995, 45, 798–803. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.D.; Lawson, P.A. The genus Abiotrophia (Kawamura et al.) is not monophyletic: Proposal of Granulicatella gen. nov., Granulicatella adiacens comb. nov., Granulicatella elegans comb. nov. and Granulicatella balaenopterae comb. nov. Int. J. Syst. Evol. Microbiol. 2000, 50, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Patel, T.; Molloy, A.; Smith, R.; Balakrishnan, I. Successful treatment of Leuconostoc bacteremia in a neutropenic patient with tigecycline. Infect. Dis. Rep. 2012, 4, e31. [Google Scholar] [CrossRef]
- Albanese, A.; Spanu, T.; Sali, M.; Novegno, F.; D’Inzeo, T.; Santangelo, R. Molecular identification of Leuconostoc mesenteroides as a cause of brain abscess in an immunocompromised patient. J. Clin. Microbiol. 2006, 44, 3044–3045. [Google Scholar] [CrossRef]
- Ke, D.; Picard, F.J.; Martineau, F.; Ménard, C.; Roy, P.H.; Ouellette, M.; Bergeron, M.G. Development of a PCR assay for rapid detection of enterococci. J. Clin. Microbiol. 1999, 37, 3497–3503. [Google Scholar] [CrossRef]
- Arbique, J.C.; Poyart, C.; Trieu-Cuot, G.; Quesne, M.; Carvalho, G.S.; Steigerwalt, A.G.; Morey, R.E.; Jackson, D.; Davidson, R.J.; Facklam, R.R. Accuracy of phenotypic and genotypic testing for identification of Streptococcus pneumoniae and description of Streptococcus pseudopneumoniae sp. nov. J. Clin. Microbiol. 2004, 42, 4686–4696. [Google Scholar] [CrossRef]
- Harf-Monteil, C.; Granello, C.; Le Brun, C.; Monteil, H.; Riegel, P. Incidence and pathogenic effect of Streptococcus pseudopneumoniae. J. Clin. Microbiol. 2006, 44, 2240–2241. [Google Scholar] [CrossRef]
- Nielsen, X.C.; Justesen, U.S.; Dargis, R.; Kemp, M.; Christensen, J.J. Identification of clinically relevant nonhemolytic Streptococci on the basis of sequence analysis of 16S-23S intergenic spacer region and partial gdh gene. J. Clin. Microbiol. 2009, 47, 932–939. [Google Scholar] [CrossRef]
- Garnier, F.; Gerbaud, G.; Courvalin, P.; Galimand, M. Identification of clinically relevant viridans group streptococci to the species level by PCR. J. Clin. Microbiol. 1997, 35, 2337–2341. [Google Scholar] [CrossRef] [PubMed]
- Harju, I.; Lange, C.; Kostrzewa, M.; Maier, T.; Rantakokko-Jalava, K.; Haanperä, M. Improved differentiation of Streptococcus pneumoniae and other S. mitis group streptococci by MALDI Biotyper using an improved MALDI Biotyper database content and a novel result interpretation algorithm. J. Clin. Microbiol. 2017, 55, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.E.; Li, K.; Filkins, L.M.; Zhu, B.; Kuchma, S.L.; Schwartzman, J.D.; O’Toole, G.A. Pseudomonas aeruginosa can inhibit growth of streptococcal species via siderophore production. J. Bacteriol. 2019, 201, e00014–e00019. [Google Scholar] [CrossRef] [PubMed]
- Besser, J.; Carleton, H.A.; Gerner-Smidt, P.; Lindsey, R.L.; Trees, E. Next-generation sequencing technologies and their application to the study and control of bacterial infections. Clin. Microbiol. Infect. 2018, 24, 335–341. [Google Scholar] [CrossRef]
- Woo, P.C.; Leung, A.S.; Leung, K.W.; Yuen, K.Y. Identification of slide coagulase positive, tube coagulase negative Staphylococcus aureus by 16S ribosomal RNA gene sequencing. Mol. Pathol. 2001, 54, 244–247. [Google Scholar] [CrossRef]
- Sabat, A.J.; Van Zanten, E.; Akkerboom, V.; Wisselink, G.; van Slochteren, K.; de Boer, R.F.; Hendrix, R.; Friedrich, A.W.; Rossen, J.W.A.; Kooistra-Smid, A.M.D. Targeted next-generation sequencing of the 16S-23S rRNA region for culture-independent bacterial identification-increased discrimination of closely related species. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate No. | Serotype | Species | Vitek® 2 System Results | MALDI-TOF MS | |
|---|---|---|---|---|---|
| Probability of Identification | Discrimination Level | Species (Score Values) | |||
| E15 | G | Enterococcus avium | 95% | Very good | not performed |
| E2 | D | Enterococcus casseliflavus | 98% | Excellent | not performed |
| E5 | AGL | Enterococcus durans | 94% | Very good | not performed |
| E28 | AGL | Enterococcus faecalis | 99% | Excellent | not performed |
| E10 | D | Enterococcus raffinosus | 94% | Very good | not performed |
| 1375/11 | - | Globicatella sanguinis | 96% | Excellent | Globicatella sanguinis (2.39) |
| PL434 | D | Kocuria rosea | 90% | Good | Granulicatella adiacens (2.22) |
| 3696/08 | D | Leuconostoc citreum | 97% | Excellent | Leuconostoc citreum (2.10) |
| 1113/11 | AGL | Leuconostoc lactis | 97% | Excellent | Leuconostoc lactis (2.20) |
| S19 | B | Streptococcus agalactiae | 97%, 99% | Excellent | not performed |
| 4734/08 | C | Streptococcus anginosus | 97% | Excellent | not performed |
| 5898/10 | AGL | Streptococcus anginosus | 95% | Very good | not performed |
| 1816/15 | - | Streptococcus mitis/oralis | 99% | Excellent | not performed |
| 1226/14 | - | Streptococcus mitis/oralis | 95% | Very good | not performed |
| 1107/09 | C | Streptococcus gordonii | 97% | Excellent | not performed |
| 6922/09 | C | Streptococcus anginosus/gordonii | 96% | Low | not performed |
| 1860/08 | G | Streptococcusagalactiae/dysgalactiae | 98% | Low | not performed |
| 8190/10 | G | Streptococcus dysgalactiae | 96% | Excellent | not performed |
| 5010/12 | AGL | Streptococcus gallolyticus | 99% | Excellent | not performed |
| S18 | AGL | Streptococcus gallolyticus | 97% | Excellent | not performed |
| PL427 | - | Streptococcus mitis/oralis | 99% | Excellent | not performed |
| PL428 | - | Streptococcus lutetiensis | 97% | Excellent | not performed |
| PL431 | D | Streptococcus mitis/oralis | 90% | Good | not performed |
| S16 | - | Streptococcus parasanguinis | 99% | Excellent | not performed |
| S63 | C | Streptococcus mitis/oralis | 98% | Excellent | not performed |
| 1374/11 | - | Streptococcus mitis/oralis | 99% | Excellent | not performed |
| p63 | C | Streptococcus pneumoniae | 99% | Excellent | not performed |
| p41 | D | × | × | × | Streptococcus pneumoniae (2.08) |
| S47 | A | Streptococcus pyogenes | 95% | Very good | not performed |
| PL433 | G | Streptococcus salivarius | 96% | Excellent | not performed |
| Identified Species | Isolate No. | 16S rRNA Gene | sodA Gene | tuf Gene | rpoB Gene |
|---|---|---|---|---|---|
| Enterococcus avium | E15 | Enterococcus avium | Enterococcus avium | Enterococcus avium | Enterococcus avium |
| Enterococcus casseliflavus | E2 | Enterococcus casseliflavus | Enterococcus casseliflavus | Enterococcus casseliflavus | Enterococcus casseliflavus |
| Enterococcus durans | E5 | Enterococcus durans | Enterococcus durans | Enterococcus durans | Enterococcus durans |
| Enterococcus faecalis | E28 | Enterococcus faecalis | Enterococcus faecium | Enterococcus faecalis | Enterococcus faecalis |
| Enterococcus raffinosus | E10 | Enterococcus raffinosus/gilvus | Enterococcus raffinosus | Enterococcus raffinosus | Enterococcus raffinosus |
| Globicatella sanguinis | 1375/11 | Globicatella sanguinis | Globicatella sanguinis | Globicatella sanguinis | Globicatella sanguinis |
| Granulicatella adiacens | PL434 | Granulicatella adiacens | Granulicatella adiacens | Granulicatella adiacens | Granulicatella adiacens |
| Leuconostoc citreum | 3696/08 | Leuconostoc citreum | no amplification product | Leuconostoc citreum | Leuconostoc citreum |
| Leuconostoc lactis | 1113/11 | Leuconostoc lactis /garlicum | × | Leuconostoc lactis /garlicum | Leuconostoc lactis |
| Streptococcus agalactiae | S19 | Streptococcus agalactiae | Streptococcus agalactiae | Streptococcus agalactiae | Streptococcus agalactiae |
| Streptococcus anginosus | 4734/08 | Streptococcus anginosus | Streptococcus anginosus | Streptococcus anginosus | Streptococcus anginosus |
| Streptococcus anginosus | 5898/10 | Streptococcus anginosus | Streptococcusanginosus/milleri | Streptococcus anginosus/milleri | Streptococcus anginosus/ intermedius |
| Streptococcus australis | 1816/15 | Streptococcus australis | Streptococcus australis | Streptococcus australis | Streptococcus australis |
| Streptococcus australis | 1226/14 | Streptococcus australis/sanguinis | Streptococcus australis | Streptococcus australis | Streptococcus australis |
| Streptococcus constellatus | 1107/09 | Streptococcus constellatus | Streptococcus anginosus/milleri | Streptococcus anginosus/milleri | Streptococcus constellatus |
| Streptococcus constellatus | 6922/09 | Streptococcus constellatus | Streptococcus anginosus/milleri | Streptococcus anginosus/milleri | Streptococcus constellatus |
| Streptococcus dysgalactiae | 1860/08 | Streptococcus dysgalactiae | Streptococcus dysgalactiae | Streptococcus dysgalactiae | Streptococcus dysgalactiae |
| Streptococcus dysgalactiae | 8190/10 | Streptococcus dysgalactiae | Streptococcus dysgalactiae | Streptococcus dysgalactiae | Streptococcus dysgalactiae |
| Streptococcus gallolyticus | 5010/12 | Streptococcus gallolyticus | Streptococcus gallolyticus | Streptococcus gallolyticus | Streptococcus gallolyticus |
| Streptococcus gallolyticus | S18 | Streptococcus gallolyticus | Streptococcus gallolyticus | Streptococcus gallolyticus | Streptococcus gallolyticus |
| Streptococcus infantis | PL427 | Streptococcus infantis | Streptococcus infantis | Streptococcus oralis | Streptococcusaustralis/infantis |
| Streptococcus lutetiensis | PL428 | Streptococcus lutetiensis | Streptococcus lutetiensis/ infantarius | Streptococcus lutetiensis/ infantarius | Streptococcus lutetiensis |
| Streptococcus mitis | PL431 | Streptococcus mitis | Streptococcus mitis | Streptococcus mitis | Streptococcus mitis |
| Streptococcus oralis | S16 | Streptococcus oralis | Streptococcus oralis | Streptococcus oralis/infantis | Streptococcus oralis |
| Streptococcus oralis | S63 | Streptococcus oralis | Streptococcus oralis | Streptococcus oralis/infantis | Streptococcus oralis |
| Streptococcus parasanguinis | 1374/11 | Streptococcus parasanguinis | Streptococcus parasanguinis/oralis | Streptococcus parasanguinis | Streptococcus parasanguinis |
| Streptococcus pneumoniae | p63 | Streptococcus pneumoniae | Streptococcus pneumoniae | Streptococcus pneumoniae | Streptococcus pneumoniae |
| Streptococcus pseudopneumoniae | p41 | Streptococcus pneumoniae/mitis | Streptococcus mitis/cristatus | Streptococcus pneumoniae/mitis | Streptococcus pseudopneumoniae/mitis |
| Streptococcus pyogenes | S47 | Streptococcus pyogenes | Streptococcus pyogenes/dysgalactiae | Streptococcus pyogenes | Streptococcus pyogenes |
| Streptococcus salivarius | PL433 | Streptococcus salivarius | Streptococcus salivarius | Streptococcus salivarius | Streptococcus salivarius |
| Streptococcus spp. (n = 21 Strains) Enterococcus spp. (n = 5 Strains) | Unambiguous Species Identification | No Identification at the Species Level | ||
|---|---|---|---|---|
| Sanger Sequencing | 16S rRNA gene | Enterococcus | 4 (80%) | 1 (20%) |
| Streptococcus | 19 (90%) | 2(10%) | ||
| sodA gene | Enterococcus | 4 (80%) | 1 (20%) | |
| Streptococcus | 12 (57%) | 9 (43%) | ||
| tuf gene | Enterococcus | 5 (100%) | 0 (0%) | |
| Streptococcus | 13 (62%) | 8 (38%) | ||
| rpoB gene | Enterococcus | 5 (100%) | 0 (0%) | |
| Streptococcus | 18 (86%) | 3 (14%) | ||
| Species | Isolate Number | |
|---|---|---|
| Enterococcus n = 5 | Enterococcus avium | E15 c |
| Enterococcus casseliflavus | E2 c | |
| Enterococcus durans | E5 c | |
| Enterococcus faecalis | E28 c | |
| Enterococcus raffinosus | E10 c | |
| Other species n = 4 | Globicatella sanguinis | 1375/11 a |
| Granulicatella adiacens | PL434 b | |
| Leuconostoc citreum | 3696/08 a | |
| Leuconostoc lactis | 1113/11 a | |
| Streptococcus n = 21 | Streptococcus agalactiae | S19 c |
| Streptococcus anginosus | 4734/08 a | |
| 5898/10 a | ||
| Streptococcus australis | 1816/15 a | |
| 1226/14a | ||
| Streptococcus constellatus | 1107/09 a | |
| 6922/09 a | ||
| Streptococcus dysgalactiae | 1860/08 a | |
| 8190/10 a | ||
| Streptococcus gallolyticus | 5010/12 a | |
| S18 c | ||
| Streptococcus infantis | PL427 b | |
| Streptococcus lutetiensis | PL428 b | |
| Streptococcus mitis | PL431 b | |
| Streptococcus oralis | S16 c | |
| S63 c | ||
| Streptococcus parasanguinis | 1374/11 a | |
| Streptococcus pneumoniae | p63 c | |
| Streptococcus pseudopneumoniae | p41 c | |
| Streptococcus pyogenes | S47 c | |
| Streptococcus salivarius | PL433 b |
| Target Gene | PCR Program | Cycles (Steps 2–4) | Amplification Primers (5′→3′) | Amount of Sequenced PCR Product | Reference |
|---|---|---|---|---|---|
| 16S rRNA (1284-bp) |
| 25 | LPW57 AGTTTGATCCTGGCTCAG | 250 ng | [68] |
| LPW58 AGGCCCGGGAACGTATTCAC | |||||
| sodA (430-bp) |
| 35 | d1 CCITAYICITAYGAYGCIYTIGARCC | 100 ng | [24] |
| d2 ARRTARTAIGCRTGYTCCCAIACRTC | |||||
| tuf (830-bp) |
| 30 | tuf-F CCAATGCCACAAACTCGT | 200 ng | [25] |
| tuf-R CCTGAACCAACAGTACGT | |||||
| rpoB (860-bp) |
| 35 | Strepto-F AARYTIGGMCCTGAAGAAAT | 200 ng | [26] |
| Strepto-R TGIARTTTRTCATCAACCATGTG | |||||
| recA (850-bp) |
| 30 | RStrGseq81 GAAAWWIATYGARAAAGAITTTGGTAA | 150 ng | [31] |
| RStrGseq937 TTYTCAGAWCCTTGICCAATYTTYTC |
| Target Gene | PCR Program | Cycles (Steps 2–4) | Amplification Primers (5′→3′) | Amount of Sequenced PCR Product | Reference |
|---|---|---|---|---|---|
| sodA (430-bp) |
| 30 | sodA-F TRCAYCATGAYAARCACCAT | 100 ng | [21] |
| sodA-R ARRTARTAMGCRTGYTCCCARACRTC | |||||
| tuf (830-bp) |
| 35 | U1 AAYATGATIACIGGIGCIGCICARATGGA | 200 ng | [60] |
| U2 AYRTTITCICCIGGCATIACCAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kosecka-Strojek, M.; Wolska, M.; Żabicka, D.; Sadowy, E.; Międzobrodzki, J. Identification of Clinically Relevant Streptococcus and Enterococcus Species Based on Biochemical Methods and 16S rRNA, sodA, tuf, rpoB, and recA Gene Sequencing. Pathogens 2020, 9, 939. https://doi.org/10.3390/pathogens9110939
Kosecka-Strojek M, Wolska M, Żabicka D, Sadowy E, Międzobrodzki J. Identification of Clinically Relevant Streptococcus and Enterococcus Species Based on Biochemical Methods and 16S rRNA, sodA, tuf, rpoB, and recA Gene Sequencing. Pathogens. 2020; 9(11):939. https://doi.org/10.3390/pathogens9110939
Chicago/Turabian StyleKosecka-Strojek, Maja, Mariola Wolska, Dorota Żabicka, Ewa Sadowy, and Jacek Międzobrodzki. 2020. "Identification of Clinically Relevant Streptococcus and Enterococcus Species Based on Biochemical Methods and 16S rRNA, sodA, tuf, rpoB, and recA Gene Sequencing" Pathogens 9, no. 11: 939. https://doi.org/10.3390/pathogens9110939
APA StyleKosecka-Strojek, M., Wolska, M., Żabicka, D., Sadowy, E., & Międzobrodzki, J. (2020). Identification of Clinically Relevant Streptococcus and Enterococcus Species Based on Biochemical Methods and 16S rRNA, sodA, tuf, rpoB, and recA Gene Sequencing. Pathogens, 9(11), 939. https://doi.org/10.3390/pathogens9110939