Bacteriophage Cocktail for the Prevention of Multiple-Antibiotic-Resistant and Mono-Phage-Resistant Vibrio coralliilyticus Infection in Pacific Oyster (Crassostrea gigas) Larvae


 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Induction of Phage-Resistant Variant
2.2. Antimicrobial Susceptibility of the Phage-Resistant Variant
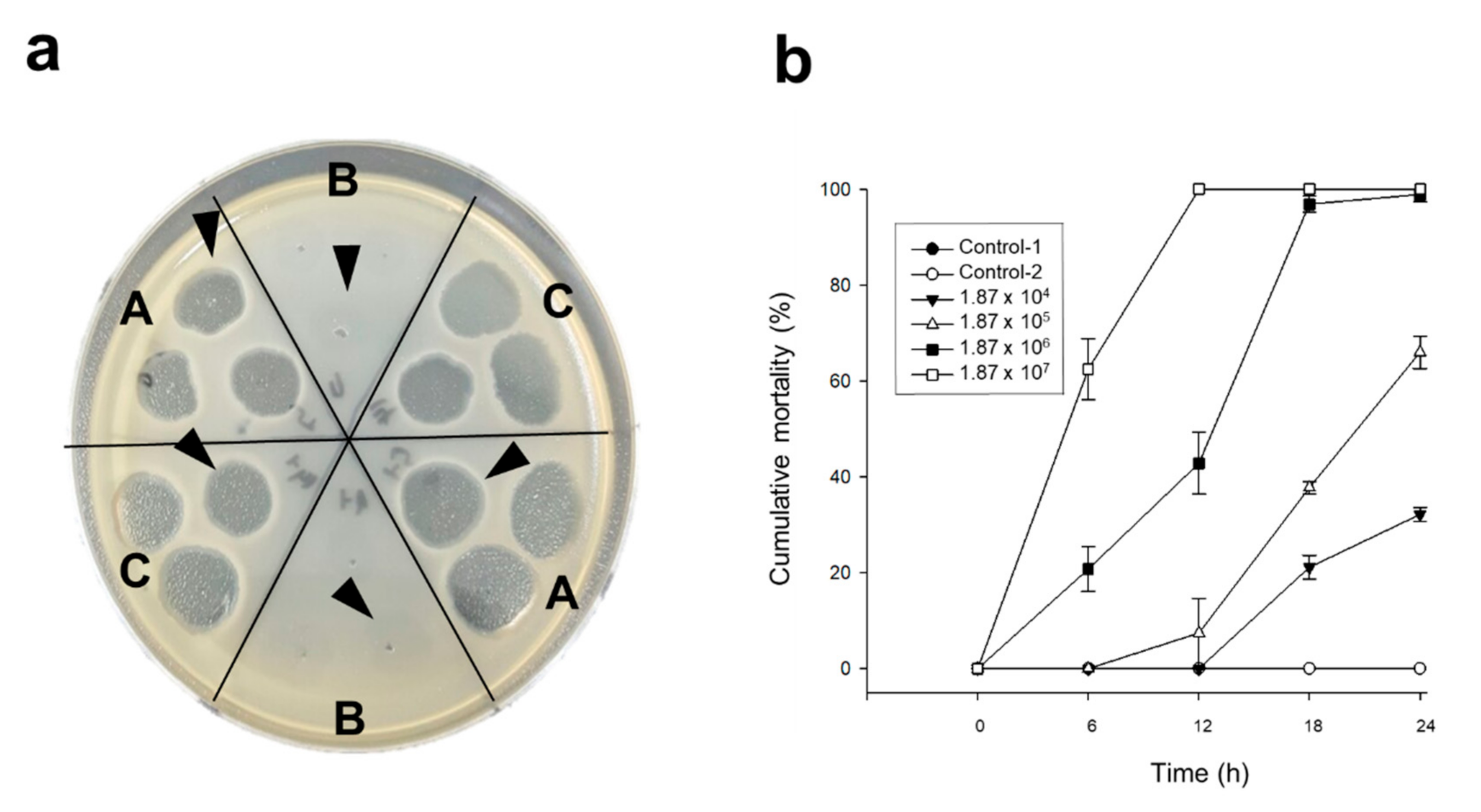
2.3. The Bactericidal Effects of Each Phage and pVco-C against VcoR-7
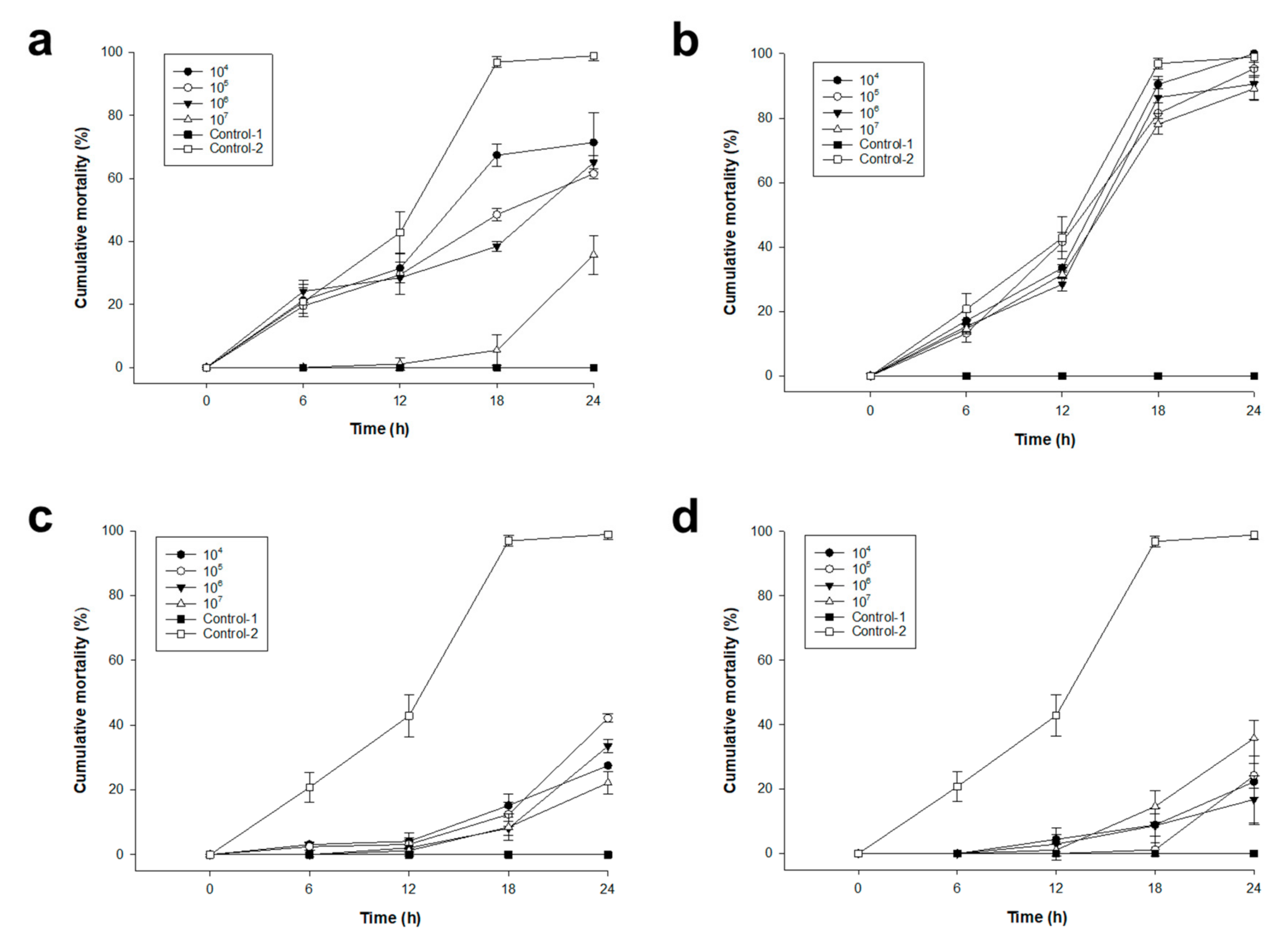
2.4. Pacific Oyster Larvae Infection Prevention Using Various Concentrations of pVco-5, pVco-7, pVco-14, and pVco-C
3. Discussion
4. Materials and Methods
4.1. Bacterial Strain and Growth Conditions
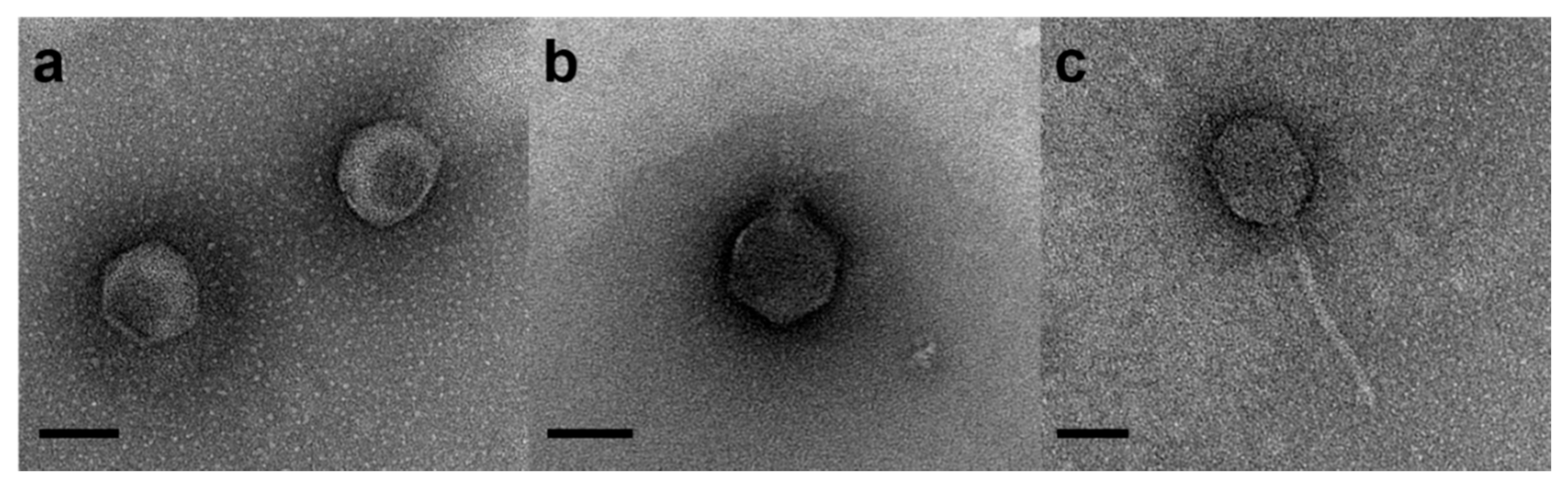
4.2. Phage Cocktail Preparation
4.3. Induction of Phage-Resistant Variants of V. coralliilyticus
4.4. Antimicrobial Susceptibility of the Phage-Resistant Variant
4.5. Pathogenicity of the Phage-Resistant Variant
4.6. Bacterial Cell Lysis Test of the Phage-Resistant Variant
4.7. Prophylactic Efficacy of the Phages and Phage Cocktail
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. FAO FishStat, Global Aquaculture Production for Species (Tonnes): Pacific Oyster. 2019. Available online: http://www.fao.org/fishery/topic/16140/en (accessed on 31 October 2019).
- Tubiash, H.S.; Chanley, P.E.; Leifson, E. Bacillary necrosis, a disease of larval and juvenile bivalve mollusks I. Etiology and epizootiology. J. Bacteriol. 1965, 90, 1036–1044. [Google Scholar]
- Elston, R.A.; Hasegawa, H.; Humphrey, K.L.; Polyak, I.K.; Häse, C.C. Re-emergence of Vibrio tubiashii in bivalve shellfish aquaculture: Severity, environmental drivers, geographic extent and management. Dis. Aquat. Org. 2008, 82, 119–134. [Google Scholar]
- Kim, H.J.; Jun, J.W.; Giri, S.S.; Chi, C.; Yun, S.; Kim, S.G.; Kim, S.W.; Han, S.J.; Kwon, J.; Oh, W.T.; et al. Identification and genome analysis of Vibrio coralliilyticus causing mortality of Pacific oyster (Crassostrea gigas) larvae. Pathogens 2020, 9, 206. [Google Scholar]
- Sugumar, G.; Nakai, T.; Hirata, Y.; Matsubara, D.; Muroga, K. Vibrio splendidus biovar II as the causative agent of bacillary necrosis of Japanese oyster Crassostrea gigas larvae. Dis. Aquat. Org. 1998, 33, 111–118. [Google Scholar]
- Travers, M.-A.; Achour, R.M.; Haffner, P.; Tourbiez, D.; Cassone, A.-L.; Morga, B.; Doghri, I.; Garcia, C.; Renault, T.; Fruitier-Arnaudin, I.; et al. First description of French V. tubiashii strains pathogenic to mollusk: I. Characterization of isolates and detection during mortality events. J. Invertebr. Pathol. 2014, 123, 38–48. [Google Scholar]
- Arzul, I.; Renault, T.; Lipart, C. Experimental herpes-like viral infections in marine bivalves: Demonstration of interspecies transmission. Dis. Aquat. Org. 2001, 46, 1–6. [Google Scholar]
- Kim, H.J.; Jun, J.W.; Giri, S.S.; Yun, S.; Kim, S.G.; Kim, S.W.; Kang, J.W.; Han, S.J.; Kwon, J.; Oh, W.T.; et al. Mass mortality in Korean bay scallop (Argopecten irradians) associated with Ostreid Herpesvirus-1 uVar. Transbound. Emerg. Dis. 2019, 66, 1442–1448. [Google Scholar]
- Farley, C.A.; Banfield, W.G.; Kasnic, G.; Foster, W.S. Oyster herpes-type virus. Scinece 1972, 178, 759–760. [Google Scholar]
- Gittenberger, A.; Voorbergen-Laarman, M.; Engelsma, M. Ostreid herpesvirus OsHV-1 µVar in Pacific oyster Crassostrea gigas (Thunberg 1793) of the Wadden Sea, a UNESCO world heritage site. J. Fish Dis. 2016, 39, 105–109. [Google Scholar]
- Moss, J.; Burreson, E.; Cordes, J.; Dungan, C.; Brown, G.; Wang, A.; Reece, K. Pathogens in Crassostrea ariakensis and other Asian oyster species: Implications for non-native oyster introduction to Chesapeake Bay. Dis. Aquat. Org. 2007, 77, 207–223. [Google Scholar]
- Renault, T.; Arzul, I. Herpes-like virus infection in hatchery-reared bivalve larvae in Europe: Specific viral DNA detection by PCR. J. Fish Dis. 2001, 24, 161–167. [Google Scholar]
- Richards, G.P.; Watson, M.A.; Needleman, D.S.; Church, K.M.; Häse, C.C. Mortalities of Eastern and Pacific oyster larvae caused by the pathogens Vibrio coralliilyticus and Vibrio tubiashii. Appl. Environ. Microbiol. 2015, 81, 292–297. [Google Scholar]
- Dubert, J.; Osorio, C.R.; Prado, S.; Barja, J.L. Persistence of antibiotic resistant Vibrio spp. in shellfish hatchery environment. Microb. Ecol. 2016, 72, 851–860. [Google Scholar]
- Nicolas, J.L.; Corre, S.; Gauthier, G.; Robert, R.; Ansquer, D. Bacterial problems associated with scallop Pecten maximus larval culture. Dis. Aquat. Org. 1996, 27, 67–76. [Google Scholar]
- Kim, H.J.; Jun, J.W.; Giri, S.S.; Chi, C.; Yun, S.; Kim, S.G.; Kim, S.W.; Kang, J.W.; Han, S.J.; Kwon, J.; et al. Application of the bacteriophage pVco-14 to prevent Vibrio coralliilyticus infection in Pacific oyster (Crassostrea gigas) larvae. J. Invertebr. Pathol. 2019, 167, 107244. [Google Scholar]
- Kim, H.J.; Giri, S.S.; Kim, S.G.; Kim, S.W.; Kwon, J.; Lee, S.B.; Park, S.C. Isolation and characterization of two bacteriophages and their preventive effects against pathogenic Vibrio coralliilyticus causing mortality of Pacific oyster (Crassostrea gigas) larvae. Microorganisms 2020, 8, 926. [Google Scholar]
- Cohen, Y.; Joseph Pollock, F.; Rosenberg, E.; Bourne, D.G. Phage therapy treatment of the coral pathogen Vibrio coralliilyticus. Microbiol. Open 2013, 2, 64–74. [Google Scholar]
- Jacquemot, L.; Bettarel, Y.; Monjol, J.; Corre, E.; Halary, S.; Desnues, C.; Bouvier, T.; Ferrier-Pages, C.; Baudoux, A.-C. Therapeutic potential of a new jumbo phage that infects Vibrio coralliilyticus, a widespread coral pathogen. Front. Microbiol. 2018, 9, 2501. [Google Scholar]
- Li, Z.; Li, X.; Zhang, J.; Wang, X.; Wang, L.; Cao, Z.; Xu, Y. Use of phages to control Vibrio splendidus infection in the juvenile sea cucumber Apostichopus japonicus. Fish. Shellfish Immunol. 2016, 54, 302–311. [Google Scholar]
- Patil, J.R.; Desai, S.N.; Roy, P.; Durgaiah, M.; Saravanan, R.S.; Vipra, A. Simulated hatchery system to assess bacteriophage efficacy against Vibrio harveyi. Dis. Aquat. Org. 2014, 112, 113–119. [Google Scholar]
- Hoai, T.D.; Mitomi, K.; Nishiki, I.; Yoshida, T. A lytic bacteriophage of the newly emerging rainbow trout pathogen Weissella ceti. Virus Res. 2018, 247, 34–39. [Google Scholar]
- Jun, J.W.; Kim, J.H.; Shin, S.P.; Han, J.E.; Chai, J.Y.; Park, S.C. Protective effects of the Aeromonas phages pAh1-C and pAh6-C against mass mortality of the cyprinid loach (Misgurnus anguillicaudatus) caused by Aeromonas hydrophila. Aquaculture 2013, 416, 289–295. [Google Scholar]
- Park, S.C.; Shimamura, I.; Fukunaga, M.; Mori, K.I.; Nakai, T. Isolation of bacteriophages specific to a fish pathogen, Pseudomonas plecoglossicida, as a candidate for disease control. Appl. Environ. Microbiol. 2000, 66, 1416–1422. [Google Scholar]
- Park, S.C.; Nakai, T. Bacteriophage control of Pseudomonas plecoglossicida infection in ayu Plecoglossus altivelis. Dis. Aquat. Org. 2003, 53, 33–39. [Google Scholar]
- Gu, J.; Liu, X.; Li, Y.; Han, W.; Lei, L.; Yang, Y.; Zhao, H.; Gao, Y.; Song, J.; Lu, R.; et al. A method for generation phage cocktail with great therapeutic potential. PLoS ONE 2012, 7, e31698. [Google Scholar]
- Chan, B.K.; Abedon, S.T.; Loc-Carrillo, C. Phage cocktails and the future of phage therapy. Future Microbiol. 2013, 8, 769–783. [Google Scholar]
- Nakai, T.; Sabour, P.; Griffiths, M. Application of bacteriophages for control of infectious diseases in aquaculture. In Bacteriophages in the Control of Food and Waterborne Pathogens; Sabour, P.M., Griffiths, M.W., Eds.; American Society for Microbiology Press: Washington, WA, USA, 2010; pp. 257–272. [Google Scholar]
- Mateus, L.; Costa, L.; Silva, Y.J.; Pereira, C.; Cunha, A.; Almeida, A. Efficiency of phage cocktails in the inactivation of Vibrio in aquaculture. Aquaculture 2014, 424, 167–173. [Google Scholar]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage resistance mechanisms. Nat. Rev. Microbiol. 2010, 8, 317–327. [Google Scholar]
- Filippov, A.A.; Sergueev, K.V.; He, Y.; Huang, X.Z.; Gnade, B.T.; Mueller, A.J.; Fernandez-Prada, C.M.; Nikolich, M.P. Bacteriophage-resistant mutants in Yersinia pestis: Identification of phage receptors and attenuation for mice. PLoS ONE 2011, 6, e25486. [Google Scholar]
- Costa, P.; Pereira, C.; Gomes, A.T.P.C.; Almeida, A. Efficiency of single phage suspensions and phage cocktail in the inactiovation of Escherichia coli and Salmonella Typhimurium: An in vitro preliminary study. Microorganisms 2019, 7, 94. [Google Scholar]
- Ma, J.; Bruce, T.J.; Jones, E.M.; Cain, K.D. A review of fish vaccine development strategies: Conventional methods and modern biotechnological approaches. Microorganisms 2019, 7, 569. [Google Scholar]
- Nakai, T.; Park, S.C. Bacteriophage therapy of infectious diseases in aquaculture. Res. Microbiol. 2002, 153, 13–18. [Google Scholar]
- Irianto, A.; Austin, B. Probiotics in aquaculture. J. Fish Dis. 2002, 25, 633–642. [Google Scholar]
- Wang, W.; Sun, J.; Liu, C.; Xue, Z. Application of immunostimulants in aquaculture: Current knowledge and future perspectives. Aquac. Res. 2016, 48, 1–13. [Google Scholar]
- Kim, H.J.; Jun, J.W.; Giri, S.S.; Chi, C.; Yun, S.; Kim, S.G.; Kim, S.W.; Kang, J.W.; Han, S.J.; Park, S.C. Complete genome sequence of a bacteriophage, pVco-5, that infects Vibrio coralliilyticus, which causes bacillary necrosis in Pacific oyster (Crassostrea gigas) larvae. Genome Announc. 2018, 6, e01143-17. [Google Scholar]
- Sambrook, V.; Russell, D.W. Molecular cloning: A laboratory manual (3-volume set). Immunology. 2001, 49, 895–909. [Google Scholar]
- Clinical and Laboratory Standards Institute. Methods for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria M45, 3rd ed.; CLSI Guideline M45; CLSI: Wayne, PA, USA, 2015. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antimicrobial Agent | Zone Diameter (mm)Interpretive Criteria | VcoR-7 | Antimicrobial Agent | Zone Diameter (mm)Interpretive Criteria | VcoR-7 | ||||
|---|---|---|---|---|---|---|---|---|---|
| S * | I * | R * | Zone Diameter/Criteria | S | I | R | Zone Diameter/Criteria | ||
| Ampicillin | ≥17 | 14–16 | ≤13 | 0/R | Imipenem | ≥23 | 20–22 | ≤19 | 26/S |
| Amoxicillin-clavulanate | ≥18 | 14–17 | ≤13 | 16/I | Meropenem | ≥23 | 20–22 | ≤19 | 18/R |
| Ampicillin-sulbactam | ≥15 | 12–14 | ≤11 | 11/R | Amikacin | ≥17 | 15–16 | ≤14 | 15/I |
| Piperacillin | ≥21 | 18–20 | ≤17 | 0/R | Gentamicin | ≥15 | 13–14 | ≤12 | 15/S |
| Piperacillin-tazobactam | ≥21 | 18–20 | ≤17 | 20/I | Tetracycline | ≥15 | 12–14 | ≤11 | 20/S |
| Cefepime | ≥25 | 19–24 | ≤18 | 18/R | Ciprofloxacin | ≥21 | 16–20 | ≤15 | 25/S |
| Cefotaxime | ≥26 | 23–25 | ≤22 | 21/R | Levofloxacin | ≥17 | 14–16 | ≤13 | 25/S |
| Cefoxitin | ≥18 | 15–17 | ≤14 | 14/R | Ofloxacin | ≥16 | 13–15 | ≤12 | 22/S |
| Ceftazidime | ≥21 | 18–20 | ≤17 | 15/R | Trimethoprim-sulfamethoxazole | ≥16 | 11–15 | ≤10 | 27/S |
| Cefuroxime sodium | ≥18 | 15–17 | ≤14 | 19/S | |||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.J.; Jun, J.W.; Giri, S.S.; Kim, S.G.; Kim, S.W.; Kwon, J.; Lee, S.B.; Chi, C.; Park, S.C. Bacteriophage Cocktail for the Prevention of Multiple-Antibiotic-Resistant and Mono-Phage-Resistant Vibrio coralliilyticus Infection in Pacific Oyster (Crassostrea gigas) Larvae. Pathogens 2020, 9, 831. https://doi.org/10.3390/pathogens9100831
Kim HJ, Jun JW, Giri SS, Kim SG, Kim SW, Kwon J, Lee SB, Chi C, Park SC. Bacteriophage Cocktail for the Prevention of Multiple-Antibiotic-Resistant and Mono-Phage-Resistant Vibrio coralliilyticus Infection in Pacific Oyster (Crassostrea gigas) Larvae. Pathogens. 2020; 9(10):831. https://doi.org/10.3390/pathogens9100831
Chicago/Turabian StyleKim, Hyoun Joong, Jin Woo Jun, Sib Sankar Giri, Sang Guen Kim, Sang Wha Kim, Jun Kwon, Sung Bin Lee, Cheng Chi, and Se Chang Park. 2020. "Bacteriophage Cocktail for the Prevention of Multiple-Antibiotic-Resistant and Mono-Phage-Resistant Vibrio coralliilyticus Infection in Pacific Oyster (Crassostrea gigas) Larvae" Pathogens 9, no. 10: 831. https://doi.org/10.3390/pathogens9100831
APA StyleKim, H. J., Jun, J. W., Giri, S. S., Kim, S. G., Kim, S. W., Kwon, J., Lee, S. B., Chi, C., & Park, S. C. (2020). Bacteriophage Cocktail for the Prevention of Multiple-Antibiotic-Resistant and Mono-Phage-Resistant Vibrio coralliilyticus Infection in Pacific Oyster (Crassostrea gigas) Larvae. Pathogens, 9(10), 831. https://doi.org/10.3390/pathogens9100831