Epidemiology of Noble Pen Shell (Pinna nobilis L. 1758) Mass Mortality Events in Adriatic Sea Is Characterised with Rapid Spreading and Acute Disease Progression

 , , ,
, , ,  ,
,  and
and 
Abstract
1. Introduction
2. Results
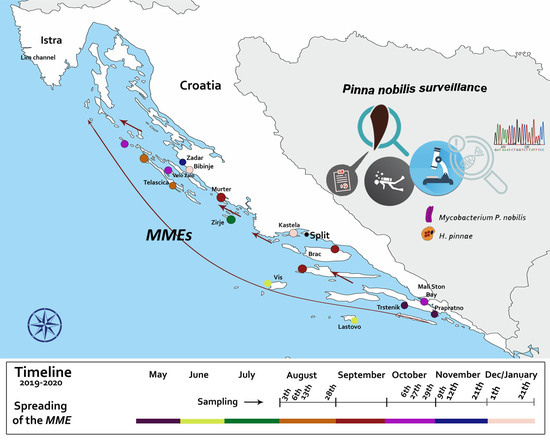
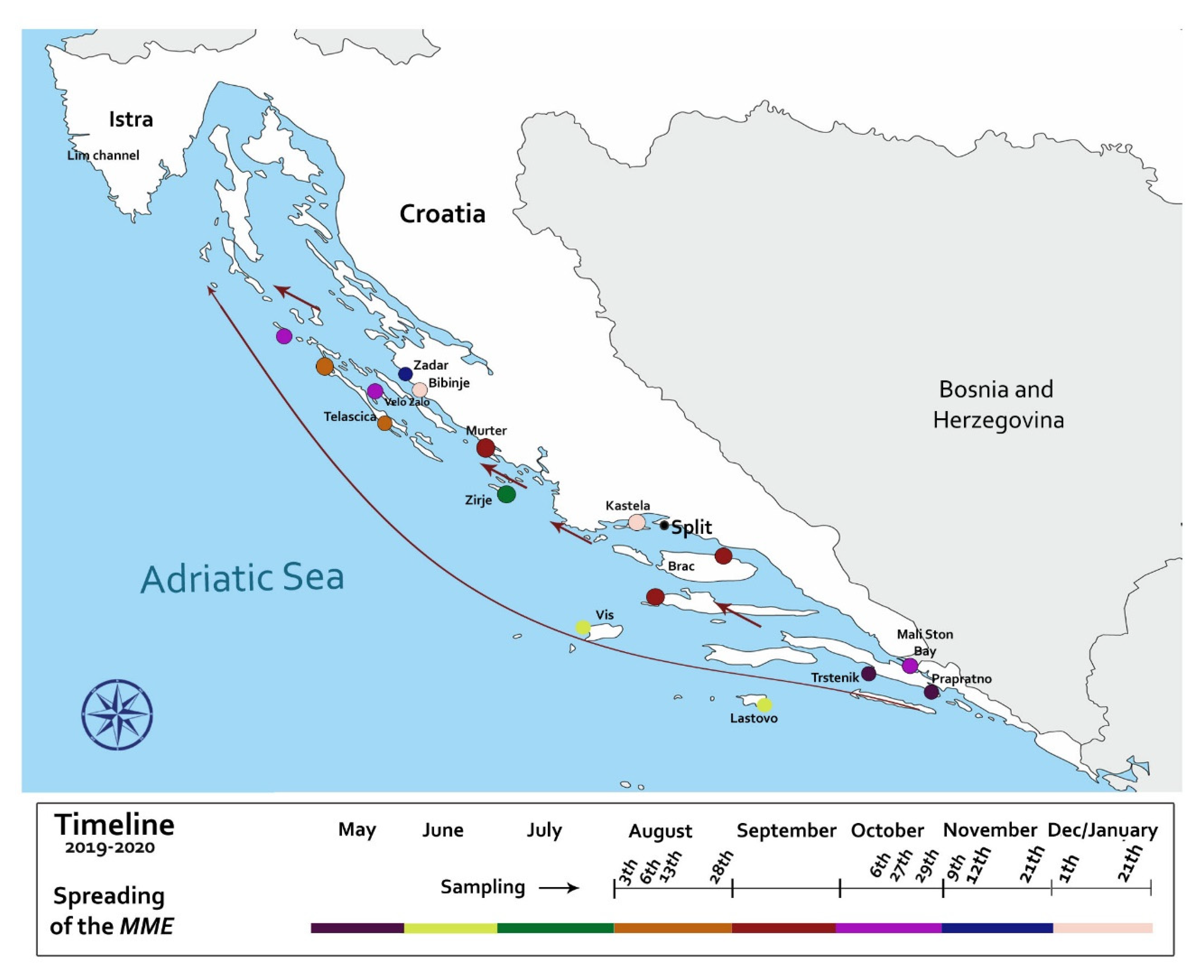
2.1. Disease Epidemiology and Pathogenesis in the Adriatic Sea
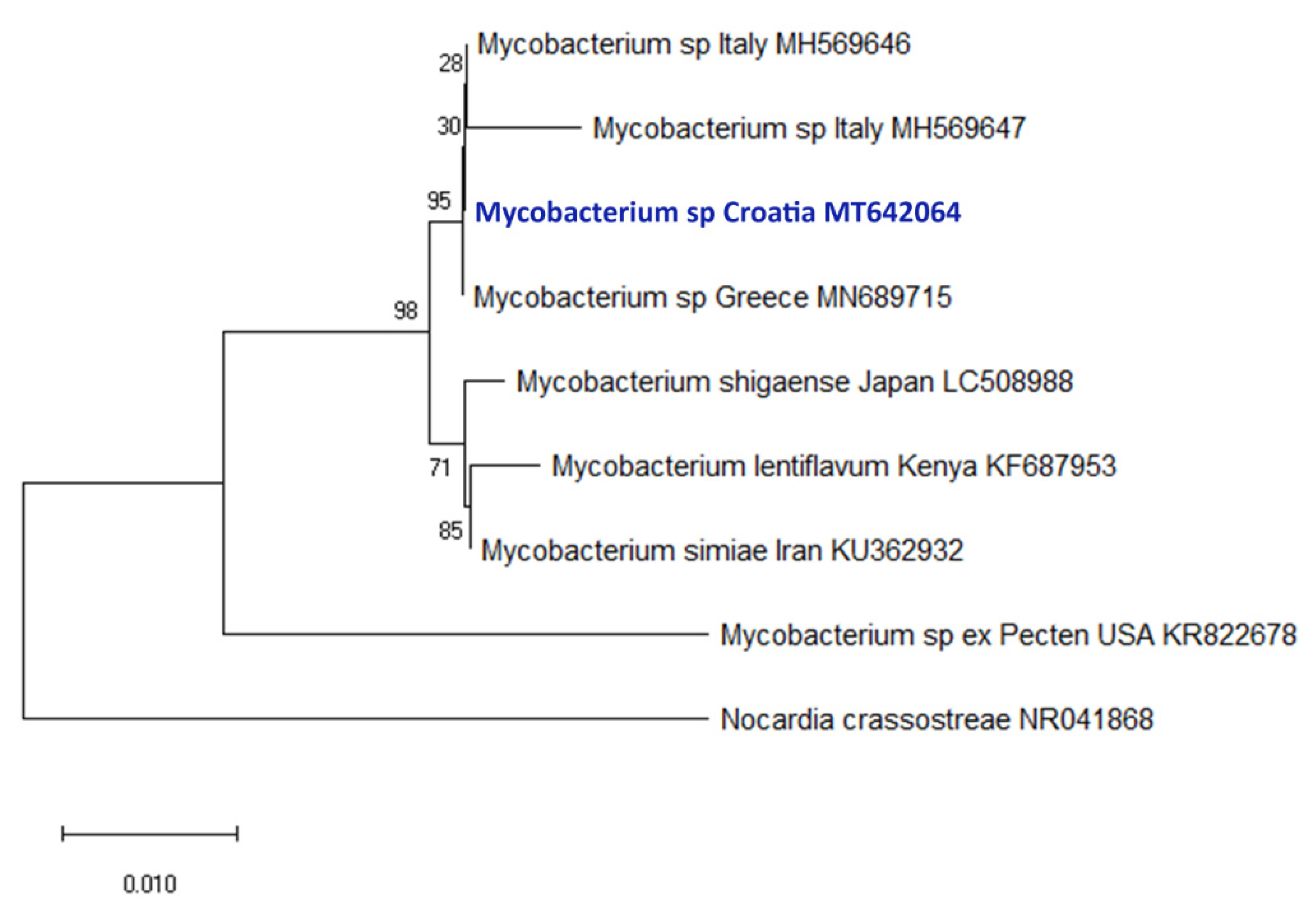
2.2. Molecular Diagnostic
3. Discussion and Conclusions
4. Materials and Methods
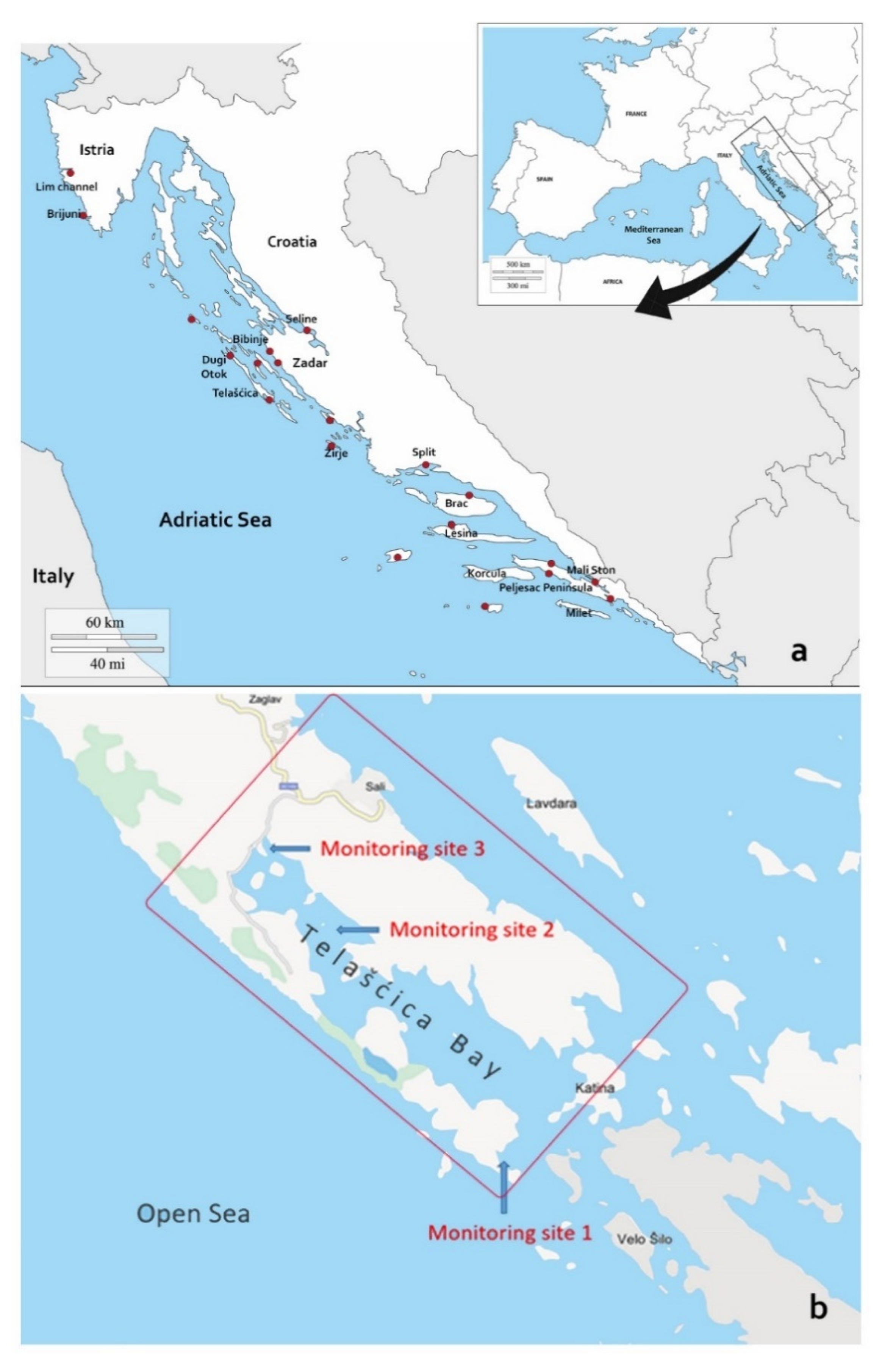
4.1. Sampling Locations and Clinical Disease Surveillance
4.2. Laboratory Diagnostic Examinations
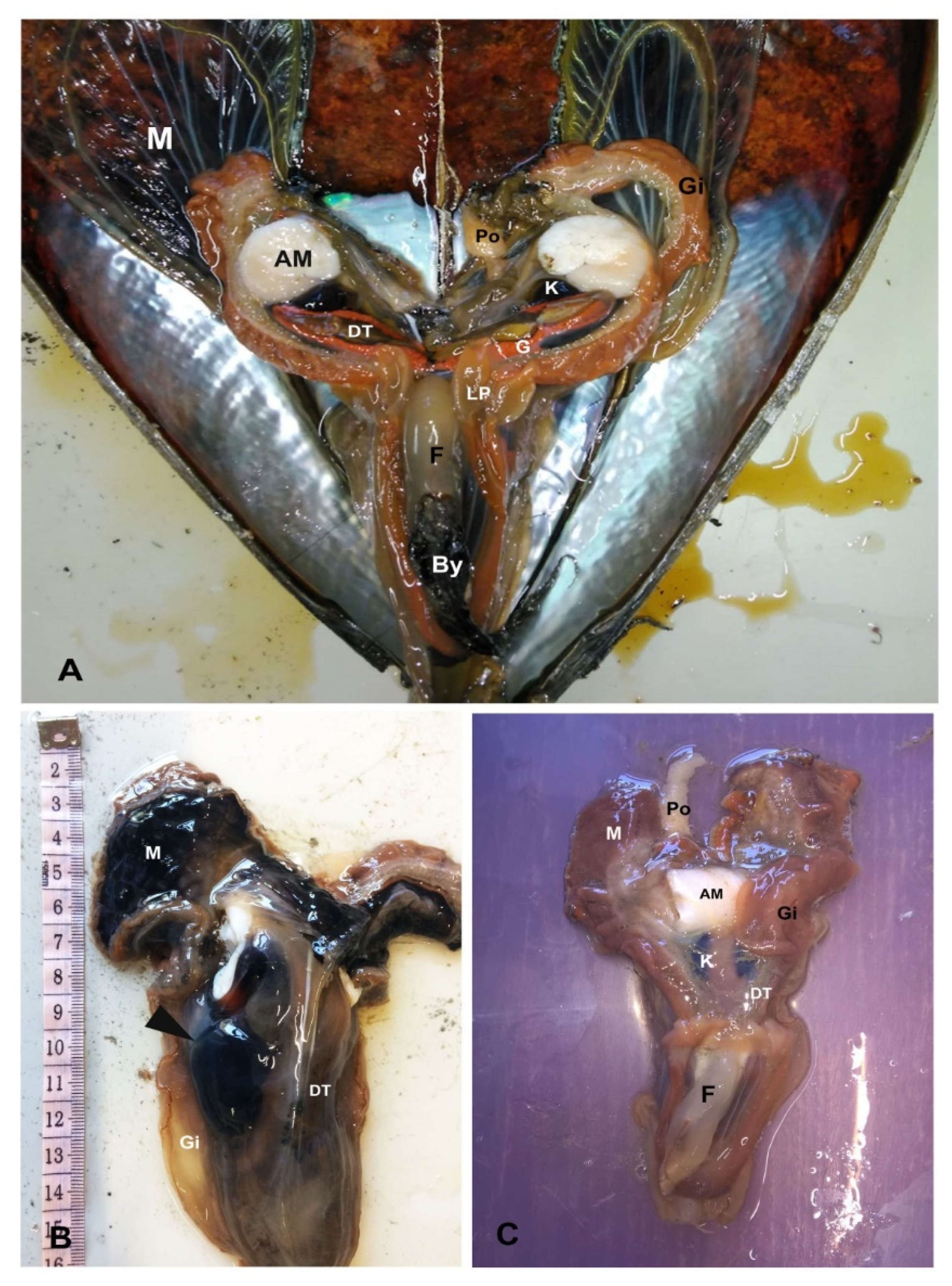
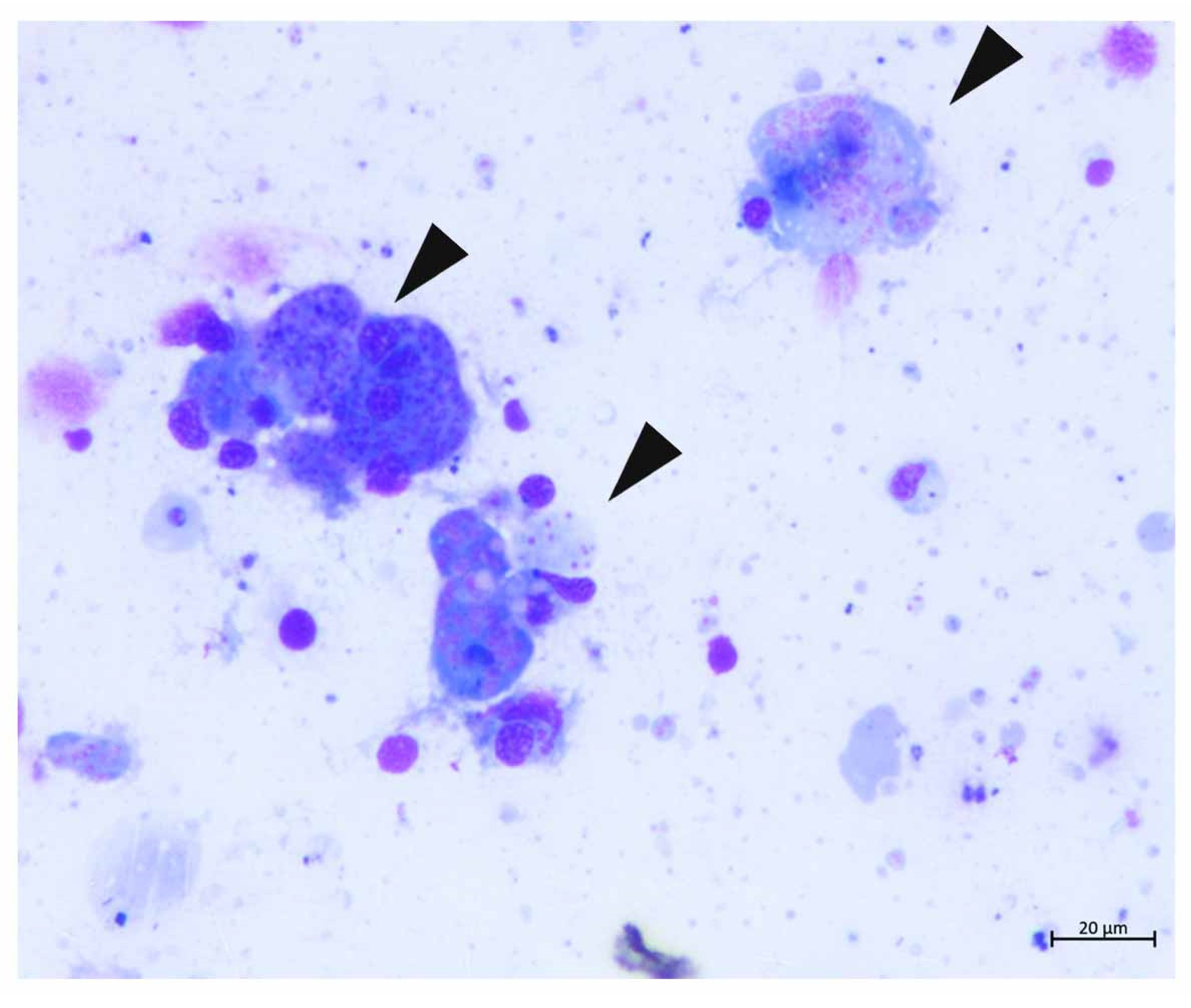
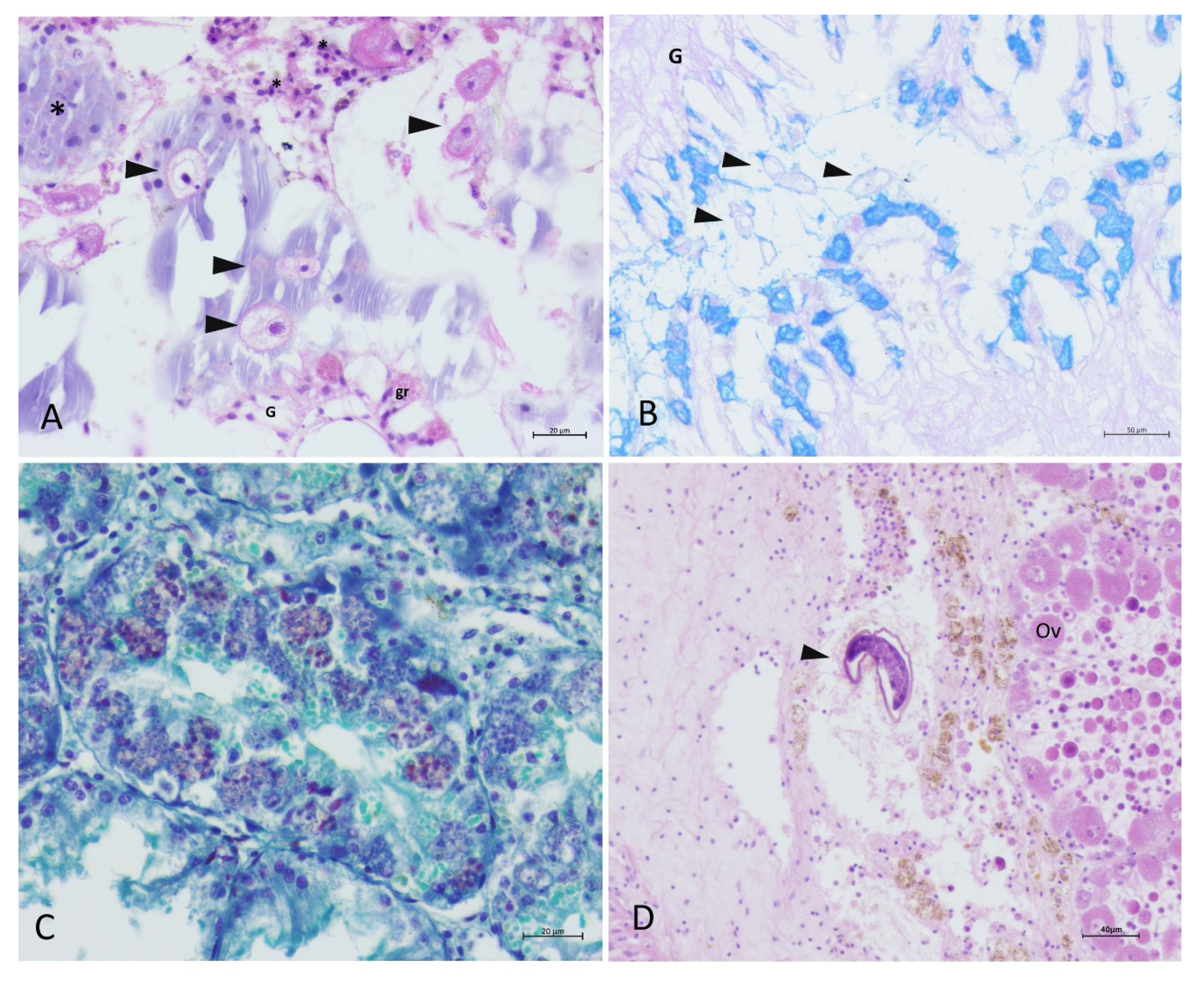
4.2.1. Gross and Microscopical Evaluation
4.2.2. DNA Isolation, PCR Diagnosis, and Phylogenetic Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fey, S.B.; Siepielski, A.M.; Nusslé, S.; Cervantes-Yoshida, K.; Hwan, J.L.; Huber, E.R.; Fey, J.M.; Catenazzi, A.; Carlson, M.S. Recent shifts in the occurrence, cause, and magnitude of animal mass mortality events. Proc. Natl. Acad. Sci. USA 2015, 112, 1083–1088. [Google Scholar] [CrossRef] [PubMed]
- Darriba, S. First haplosporidan parasite reported infecting a member of the Superfamily Pinnoidea (Pinna nobilis) during a mortality event in Alicante (Spain, Western Mediterranean). J. Invertebr. Pathol. 2017, 148, 14–19. [Google Scholar] [CrossRef]
- Catanese, G.; Grau, A.; Valencia, J.M.; Garcia-March, J.R.; Vázquez-Luis, M.; Alvarez, E.; Deudero, S.; Darriba, S.; Carballal, M.J.; Villalba, A. Haplosporidium pinnae sp. nov., a haplosporidan parasite associated with mass mortalities of the fan mussel, Pinna nobilis, in the Western Mediterranean Sea. J. Invertebr. Pathol. 2018, 157, 9–24. [Google Scholar] [CrossRef]
- Cabanellas-Reboredo, M.; Vázquez-Luis, M.; Mourre, B.; Álvarez, E.; Deudero, S.; Amores, Á.; Addis, P.; Ballesteros, E.; Barrajón, A.; Coppa, S.; et al. Tracking a mass mortality outbreak of pen shell Pinna nobilis populations: A collaborative effort of scientists and citizens. Sci. Rep. 2019, 9, 13355. [Google Scholar] [CrossRef]
- Kersting, D.; Benabdi, M.; Čižmek, H.; Grau, A.; Jimenez, C.; Katsanevakis, S.; Öztürk, B.; Tuncer, S.; Tunesi, L.; Vázquez-Luis, M.; et al. Pinna nobilis. The IUCN Red List of Threatened Species 2019; IUCN Red List: London, UK, 2019. [Google Scholar] [CrossRef]
- Carella, F.; Aceto, S.; Pollaro, F.; Miccio, A.; Iaria, C.; Carrasco, N.; Prado, P.; De Vico, G. A mycobacterial disease is associated with the silent mass mortality of the pen shell Pinna nobilis along the Tyrrhenian coastline of Italy. Sci. Rep. 2019, 9, 2725. [Google Scholar] [CrossRef] [PubMed]
- Loopez-Sanmartin, M.; Lopez Fernandez, J.R.; De La Herran, R.; Garcia-March, J.R.; Navas, J.I. Evidence of mycobacterial presence in Pinna nobilis infected by Haplosporidium pinnae maintained under quarantine conditions. In Proceedings of the II Congreso de Juvenes Investigadores del Mar, Malaga, Spain, 1–4 October 2019. [Google Scholar]
- Lattos, A.; Giantsis, I.A.; Karagiannis, D.; Michaelidis, B. First detection of the invasive Haplosporidian and Mycobacteria parasites hosting the endangered bivalve Pinna nobilis in Thermaikos Gulf, North Greece. Mar. Environ. Res. 2020, 155, 104889. [Google Scholar] [CrossRef]
- Carella, F.; Elisabetta, A.; Simone, F.; Fulvio, S.; Daniela, M.; Prado, P.; Rossella, P.; Marino, F.; Eleonora, F.; Tobia, P.; et al. In the Wake of the Ongoing Mass Mortality Events: Co-occurrence of Mycobacterium, Haplosporidium and Other Pathogens in Pinna nobilis Collected in Italy and Spain (Mediterranean Sea). Front. Mar. Sci. 2020, 7, 48. [Google Scholar] [CrossRef]
- IUCN Pinna Nobilis. Available online: https://www.iucn.org/sites/dev/files/content/documents/pinna_nobilis_iucn_final.pdf (accessed on 14 July 2020).
- Peharda, M.; Hrs-Brenko, M.; Onofri, V.; Lučić, D.; Benović, A. A visual census of bivalve distributions in the saltwater lake Malo jezero (Mljet National Park, South Adriatic Sea). Acta Adriat. 2002, 43, 65–75. [Google Scholar]
- Šiletić, T.; Peharda, M. Population study of the fan shell Pinna nobilis L. in Malo and Veliko Jezero of the Mljet National Park (Adriatic Sea). Sci. Mar. 2003, 67, 91–98. [Google Scholar] [CrossRef]
- Richardson, C.A.; Peharda, M.; Kennedy, H.; Kennedy, P.; Onofri, V. Age, growth rate and season of recruitment of Pinna nobilis (L.) in the Croatian Adriatic determined from Mg:Ca and Sr:Ca shell profiles. J. Exp. Mar. Biol. Ecol. 2004, 299, 1–16. [Google Scholar] [CrossRef]
- Sanna, D.; Cossu, P.; Dedola, G.L.; Scarpa, F.; Maltagliati, F.; Castelli, A.; Franzoi, P.; Lai, T.; Cristo, B.; Curini-Galletti, M.; et al. Mitochondrial DNA Reveals Genetic Structuring of Pinna nobilis across the Mediterranean Sea. PLoS ONE 2013, 8, e67372. [Google Scholar] [CrossRef] [PubMed]
- Ankon, P. Genetic and Morphological Variability of the Noble Pen Shell (Pinna nobilis Linnaeus, 1758) in Nature Park Telašćica and National Park Mljet. Bachelor’s Thesis, University of Zagreb, Zagreb, Croatia, 21 June 2017. [Google Scholar]
- Marrocco, V.; Zangaro, F.; Sicuro, A.; Pinna, M. A scaling down mapping of Pinna nobilis (Linnaeus, 1758) through the combination of scientific literature, NATURA 2000, grey literature and citizen science data. Nat. Conserv. 2019, 33, 21–31. [Google Scholar] [CrossRef]
- Čižmek, H.; Čolić, B.; Gračan, R.; Grau, A.; Catanese, G. An emergency situation for pen shells in the Mediterranean: The Adriatic Sea, one of the last Pinna nobilis shelters, is now affected by a mass mortality event. J. Invertebr. Pathol. 2020, 173, 107388. [Google Scholar] [CrossRef]
- Basso, L.; Hendriks, I.E.; Duarte, C.M. Juvenile Pen Shells (Pinna nobilis) Tolerate Acidification but Are Vulnerable to Warming. Estuar. Coast. 2015, 38, 1976–1985. [Google Scholar] [CrossRef]
- Vázquez-Luis, M.; Borg, J.A.; Morell, C.; Banach-Esteve, G.; Deudero, S. Influence of boat anchoring on Pinna nobilis: A field experiment using mimic units. Mar. Freshw. Res. 2015, 66, 786–794. [Google Scholar] [CrossRef]
- Cabanellas-Reboredo, M.; Blanco, A.; Deudero, S.; Tejada, S. Effects of the invasive macroalga Lophocladia lallemandii on the diet and trophism of Pinna nobilis (Mollusca: Bivalvia) and its guests Pontonia pinnophylax and Nepinnotheres pinnotheres (Crustacea: Decapoda). Sci. Mar. 2010, 74, 101–110. [Google Scholar] [CrossRef]
- Župan, I.; Šarić, T.; Kanski, D. Struktura Populacije Zajednica Školjkaša na Području Parka Prirode Telašćica (Population Structure of Bivalves in the the Telašćica Nature Park); University of Zadar: Zadar, Croatia, 2015. (In Croatian) [Google Scholar]
- Župan, I.; Šarić, T. Utjecaj Nautičkog Turizma i Koncesioniranih Sidrišta na Morski Okoliš u Zadarskoj Županiji (Impact of Nautical Tourism and Concessioned Anchorages on the Marine Environment in Zadar County); University of Zadar: Zadar, Croatia, 2017. (In Croatian) [Google Scholar]
- Župan, I.; Šarić, T. Istraživanje Trenutnog Stanja Prirodnih Zajednica Školjkaša i Puževa s Naglaskom na Vrstu Pinna Nobilis na Području Ekološke Mreže Natura 2000 HR3000171 Ušće Krke (Research of the Current Status of Bivalves and Snails Natural Communities with an Emphasis on the Species Pinna nobilis in the Area of the Natura 2000 Ecological Network HR3000171 Krka Estuary); University of Zadar: Zadar, Croatia, 2018. (In Croatian) [Google Scholar]
- Artegiani, A.; Bregant, D.; Paschini, E.; Pinardi, N.; Raicich, F.; Russo, A. The Adriatic Sea General Circulation. Part II: Baroclinic Circulation Structure. J. Phys. Oceanogr. 1997, 27, 1515–1532. [Google Scholar] [CrossRef]
- Poulain, P.M. Adriatic Sea surface circulation as derived from drifter data between 1990 and 1999. J. Mar. Syst. 2001, 29, 3–32. [Google Scholar] [CrossRef]
- Lipizer, M.; Partescano, E.; Rabitti, A.; Giorgetti, A.; Crise, A. Qualified temperature, salinity and dissolved oxygen climatologies in a changing Adriatic Sea. Ocean Sci. 2014, 10, 771–797. [Google Scholar] [CrossRef]
- Danovaro, R.; Fonda Umani, S.; Pusceddu, A. Climate Change and the Potential Spreading of Marine Mucilage and Microbial Pathogens in the Mediterranean Sea. PLoS ONE 2009, 4, e7006. [Google Scholar] [CrossRef]
- Parras-Berrocal, I.; Vazquez, R.; Cabos, W.; Sein, D.; Mañanes, R.; Perez-Sanz, J.; Izquierdo, A. The climate change signal in the Mediterranean Sea in a regionally coupled ocean-atmosphere model. Ocean Sci. 2020, 16, 743–765. [Google Scholar] [CrossRef]
- Gutierrez, Y. Diagnostic Pathology of Parasitic Infections with Clinical Correlations, 2nd ed.; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Ricciardi, A.; Ndao, M. Diagnosis of parasitic infections: What’s going on? J. Biomol. Screen. 2015, 20, 6–21. [Google Scholar] [CrossRef] [PubMed]
- Momčilović, S.; Cantacessi, C.; Arsić-Arsenijević, V.; Otranto, D.; Tasić-Otašević, S. Rapid diagnosis of parasitic diseases: Current scenario and future needs. Clin. Microbiol. Infect. 2019, 25, 290–309. [Google Scholar] [CrossRef]
- Palmer, M.V.; Welsh, M.D.; Hostetter, J.M. Mycobacterial Diseases of Animals. Vet. Med. Int. 2011, 2011, 292469. [Google Scholar] [CrossRef][Green Version]
- Hatfull, G.F.; Jacobs, W.R. Molecular Genetics of Mycobacteria, 2nd ed.; ASM Press: Washington, DC, USA, 2014; p. 832. [Google Scholar]
- Flynn, J.A.L.; Ernst, J.D. Immune responses in tuberculosis. Curr. Opin. Immunol. 2000, 12, 432–436. [Google Scholar] [CrossRef]
- Jamwal, S.V.; Mehrotra, P.; Singh, A.; Siddiqui, Z.; Basu, A.; Rao, K.V. Mycobacterial escape from macrophage phagosomes to the cytoplasm represents an alternate adaptation mechanism. Sci. Rep. 2016, 6, 23089. [Google Scholar] [CrossRef]
- Burreson, E.M.; Stokes, N.A.; Friedman, C.S. Increased virulence in an introduced pathogen: Haplosporidium nelsoni (MSX) in the eastern oyster Crassostrea virginica. J. Aquat. Anim. Health. 2000, 12, 1–8. [Google Scholar] [CrossRef]
- Chen, P.; Shi, M.; Feng, G.D.; Liu, J.Y.; Wang, B.J.; Shi, X.D.; Ma, L.; Liu, X.D.; Yang, Y.N.; Dai, W.; et al. A highly efficient Ziehl-Neelsen stain: Identifying de novo intracellular Mycobacterium tuberculosis and improving detection of extracellular M. tuberculosis in cerebrospinal fluid. J. Clin. Microbiol. 2012, 50, 1166–1170. [Google Scholar] [CrossRef]
- Bakran-Petricioli, T.; Vacelet, J.; Zibrowius, H.; Petricioli, D.; Chevaldonné, P.; Rađa, T. New data on the distribution of the ‘deep-sea’ sponges Asbestopluma hypogea and Oopsacas minuta in the Mediterranean Sea. Mar. Ecol. 2007, 28, 10–23. [Google Scholar] [CrossRef]
- OIE, 2019, Aquatic Animal Health Code—29/08/2019. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/aahc/current/chapitre_aqua_ani_surveillance.pdf (accessed on 14 July 2020).
- Mazzi, V. Manuale di Tecniche Istologiche e Istochimiche; Piccin-Nuova Libraria: Padova, Italia, 1977. [Google Scholar]
- Feng, G.D.; Shi, M.; Ma, L.; Chen, P.; Wang, B.J.; Zhang, M.; Chang, X.L.; Su, X.C.; Yang, Y.N.; Fan, X.H.; et al. Diagnostic accuracy of intracellular Mycobacterium tuberculosis detection for tuberculous meningitis. Am. J. Respir. Crit. Care Med. 2014, 189, 475–481. [Google Scholar] [CrossRef]
- Böddinghaus, B.; Rogall, T.; Flohr, T.; Blöcker, H.; Böttger, E.C. Detection and identification of mycobacteria by amplification of rRNA. J. Clin. Microbiol. 1990, 28, 1751–1759. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Location | Coordinates | Water T °C | Substrate Type | Mortality Rate % | Comments |
|---|---|---|---|---|---|---|
| 30/05/2019 | Pelješac peninsula-Prapratno | 42°48′51″ N 17°40′33″ E | 18 | Sand/Rocks | 100 | MME-south side of the peninsula, facing towards the open sea |
| 30/05/2019 | Pelješac peninsula-Trstenik | 42°54′56″ N 17°24′08″ E | 18 | Sand/Rocks | 100 | MME-south side of the peninsula, facing towards the open sea |
| 05/06/2019 | Pelješac peninsula-Mali ston | 42°52′01″ N 17°41′55″ E | 18 | Mud/Rocks | 0 | No visible sign of disease-northern side of the peninsula, more protected from the influence of the open sea |
| 05/06/2019 | Pelješac peninsula-Janjina | 42°55′47″ N 17°26′59″ E | 19 | Sand/Mud/Cymodocea nodosa meadow | 0 | No visible sign of disease-northern side of the peninsula, more protected from the influence of the open sea |
| 09/06/2019 | Bibinje | 44°03′30″ N 15°17′08″ E | 20 | Sand/C. nodosa meadow | 0 | No visible sign of disease |
| 25/06/ 2019 | Vis | 43°02′18″ N 16°13′59″ E | 22 | Sand/Rocks | >85 | MME |
| 27/06/2019 | Lastovo | 42°44′41″ N 16°48′41″ E | 22 | Sand/Rocks | 100 | MME |
| 15/07/2019 | Žirje | 43°39′58″ N 15°39′46″ E | 23 | Sand/P. oceanica meadow | >85 | MME |
| 10/08/2019 | Bibinje | 44°03′30″ N 15°17′08″ E | 24 | Sand/C. nodosa meadow | 0 | No visible sign of disease |
| 03/08/2019 | Sakarun (Dugi Otok) | 44°07′48″ N 14°52′35″ E | 23 | Sand/P. oceanica meadow | 40 | Mortality in juveniles is lower; better reaction to stimulus |
| 06/08/2019 | Žirje | 43°40′01″ N 15°39′38″ E | 24 | Sand/P. oceanica meadow | 90 | Adults alive with visible signals of disease (valve closure weakness) |
| 13/08/2019 | Seline | 44°16′20″ N 15°30′38″ E | 25 | Sand/Rocks | 0 | No visible sign of disease |
| 28/08/2019 | Telašćica Bay (Dugi otok) | 43°55′36″ N 15°08′17″ E | 25 | Sand/Mud/P. oceanica meadow | 30–100 | The mortality rate in locations closer to the open sea was 100% but in closed bays were lower (30%) |
| 22/09/2019 | Murter | 43°46′42″ N 15°37′17″ E | 23 | Sand/Rocks | >85 | MME |
| 24/09/2019 | Hvar | 43°13′31″ N 16°32′26″ E | 23 | Sand/Rocks | 30–85 | Early signs of MME |
| 25/09/2019 | Brač | 43°20′57″ N 16°44′24″ E | 23 | Sand/Mud | 30–85 | Early signs of MME |
| 06/10/2019 | Bibinje | 44°03′30″ N 15°17′08″ E | 20 | Sand/C. nodosa meadow | 0 | No visible sign of disease in moment of sampling |
| 18/10/2019 | Premuda | 44°20′07″ N 14°35′46″ E | 20 | Sand/Mud/P. oceanica meadow | >85 | MME |
| 27/10/2019 | Velo Žalo | 44°00′15″ N 15°04′1″ E | 21 | Sand/Rocks/P. oceanica meadow | 40 | Live adult specimens with visible signals of disease |
| 29/10/2019 | Lim channel (Istra) | 45°07′55″ N 13°43′55″ E | 21 | Mud/Rocks | 0 | No visible sign of disease |
| 09/11/2019 | Mali Ston Bay | 42°52′01″ N 17°41′51″ E | 20 | Mud/Rocks | 60 | Mortality in juveniles is lower, better reaction to stimulus |
| 12/11/2019 | Zadar | 44°08′05″ N 15°12′22″ E | 18 | Sand/C. nodosa meadow | 40 | Early signs of MME |
| 21/11/2019 | Brijuni | 44°55′05″ N 13°46′13″ E | 18 | Mud | 0 | No visible sign of disease |
| 01/01/2020 | Bibinje | 44°03′30″ N 15°17′08″ E | 13 | Sand/C. nodosa meadow | 80 | In period of two months mortality rate reach 80% |
| 21/01/2020 | Kaštela | 43°33′01″ N 16°21′36″ E | 13 | Sand/C. nodosa meadow | 95 | MME |
| Sample Date (day/month/yr) | Area | Animal Shell Length (cm) | Animal Mortality | Pathogens Diagnostic | Recorded Lesions | |
|---|---|---|---|---|---|---|
| Histopathology Myco/Haplo | PCR Myco/Haplo | |||||
| 03/08/2019 | Sakarun | 31 | 40% | +/+ | +/+ | Visible plasmodial phases of H. pinnae (score 1) and presence of Mycobacterium within single immune cells infiltrate digestive tissue (score 1); |
| 03/08/2019 | Sakarun | 52 | −/+ | −/+ | Visible plasmodial phases of H. pinnae (score 1) and presence of Mycobacterium within single and immune cells aggregated within digestive tissue (score 2), areas of digestive glad necrosis and nodulation. Mycobacterium and H. pinnae presence within hemolymph vessels. | |
| 03/08/2019 | Sakarun | 19 | + /+ | +/+ | Visible plasmodial phases of H. pinnae (score 1) and presence of Mycobacterium within single immune cells infiltrate digestive tissue (score 1); | |
| 03/08/2019 | Sakarun | 47 | −/+ | n.p. * | Few plasmodial phases of H. pinnae at mantle level (score 1); no other lesions visible | |
| 03/08/2019 | Sakarun | 35 | −/+ | +/+ | No lesions visible | |
| 03/08/2019 | Sakarun | 32 | +/+ | +/+ | Scarce inflammatory response; H. pinnae (score 1); Mycobacterium (score 1); | |
| 06/8/2019 | Žirje | 54 | 95% | −/− | −/− | Strong inflammation (infiltrative and nodular) at mantle and digestive gland |
| 28/08/2019 | Telašćica | 54 | 65% | +/+ | +/+ | Strong infiltrative inflammation; H. pinnae (score 2); Mycobacterium (score 1); |
| 28/08/2019 | Telašćica | 49 | +/+ | +/+ | Strong infiltrative inflammation; presence of inflammatory nodules at digestive apparatus and connective tissue level H. pinnae (score 2); Mycobacterium (score 2); | |
| 28/08/2019 | Telašćica | 41 | +/+ | +/+ | Light infiltrative inflammation; H. pinnae (score 1); Mycobacterium (score 1); | |
| 28/08/2019 | Telašćica | 23 | +/+ | +/+ | Light infiltrative inflammation; H. pinnae (score 2); Mycobacterium (score 1); | |
| 27/10/2019 | Velo Žalo | 46 | 40% | +/− | −/− | No visible lesions; Mycobacterium (score 1); |
| 27/10/2019 | Velo Žalo | 53 | +/+ | −/+ | Sporulation phases of H. pinnae (score 3) and inflammatory nodules filled with Mycobacterium in the interstitium of digestive tissue and at fibrous capsule of digestive gland (score 2). | |
| 09/11/2019 | Mali Ston | 58 | 60% | +/− | +/− | Gill inflammation, acid mucin production with presence of ciliates; Mycobacterium (score 2) |
| 09/11/2019 | Mali Ston | 55 | +/− | +/− | Gill inflammation, acid mucin production with presence of ciliates Mycobacterium (score 2) | |
| 09/11/2019 | Mali Ston | 57 | +/− | +/− | Gill inflammation, acid mucin production with presence of ciliates Mycobacterium (score 1) | |
| 12/11/2019 | Zadar | 58 | 40% | −/+ | +/− | Light infiltrative inflammation digestive tissue; H. pinnae (score 2) |
| 01/01/2020 | Bibinje | 55 | 80% | +/− | +/+ | Strong infiltrative inflammation; Mycobacterium (score 1); |
| 01/01/2020 | Bibinje | 47 | +/+ | +/+ | Strong infiltrative and nodular inflammation; H. pinnae (score 2); Mycobacterium (score 2); | |
| 01/01/020 | Bibinje | 40 | +/− | +/+ | Strong infiltrative and nodular; Mycobacterium (score 2); | |
| 21/01/2020 | Kaštela | 49 | 95% | +/+ | +/+ | Strong infiltrative inflammation; H. pinnae (score 2); Mycobacterium (score 1); |
| 21/01/2020 | Kaštela | 38 | +/+ | +/+ | Strong infiltrative inflammation; H. pinnae (score 1); Mycobacterium (score 1); | |
| 21/01/2020 | Kaštela | 30 | −/− | −/+ | Strong infiltrative inflammation; H. pinnae (score 2); Mycobacterium (score 2); | |
| Sample Date (day/month/yr) | Area | Animal Shell Length (cm) | Animal Mortality | Pathogens Diagnostic Histo M/H | Pathogens Diagnostic PCR M/H | Lesions |
|---|---|---|---|---|---|---|
| 13/08/2019 | Seline | 58 | 0% | −/− | −/− | |
| 13/08/2019 | Seline | 48 | −/− | −/− | Digestive Inflammatory nodules with Browns cells; eosinophilic granulocytes degranulation at mantle level; gill ciliates with light inflammation | |
| 13/08/2019 | Seline | 43 | −/− | −/− | light digestive glad necrosis; Bacteria intraepithelial digestive tract; gill ciliates | |
| 13/08/2019 | Seline | 25 | −/− | −/− | gill ciliates; mucous production; No visible lesions | |
| 13/08/2019 | Seline | 33 | −/− | −/− | gill ciliates; mucous production; No visible lesions | |
| 06/10/2019 | Bibinje | 22 | 0% | −/− | −/− | No visible lesions |
| 06/10/2019 | Bibinje | 43 | −/− | −/− | No visible lesions | |
| 06/10/2019 | Bibinje | 44 | −/− | −/− | No visible lesions | |
| 29/10/2019 | Istra | 47 | 0% | +/− | +/− | |
| 29/10/2019 | Istra | 50 | −/− | −/− | No visible lesions | |
| 29/10/2019 | Istra | 55 | −/− | +/− | No visible lesions | |
| 21/11/2019 | Brijuni | 60 | 0% | −/− | −/− | No visible lesions |
| 21/11/2019 | Brijuni | 55 | −/− | −/− | No visible lesions | |
| 21/11/2019 | Brijuni | 53 | −/− | −/− | No visible lesions |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šarić, T.; Župan, I.; Aceto, S.; Villari, G.; Palić, D.; De Vico, G.; Carella, F. Epidemiology of Noble Pen Shell (Pinna nobilis L. 1758) Mass Mortality Events in Adriatic Sea Is Characterised with Rapid Spreading and Acute Disease Progression. Pathogens 2020, 9, 776. https://doi.org/10.3390/pathogens9100776
Šarić T, Župan I, Aceto S, Villari G, Palić D, De Vico G, Carella F. Epidemiology of Noble Pen Shell (Pinna nobilis L. 1758) Mass Mortality Events in Adriatic Sea Is Characterised with Rapid Spreading and Acute Disease Progression. Pathogens. 2020; 9(10):776. https://doi.org/10.3390/pathogens9100776
Chicago/Turabian StyleŠarić, Tomislav, Ivan Župan, Serena Aceto, Grazia Villari, Dušan Palić, Gionata De Vico, and Francesca Carella. 2020. "Epidemiology of Noble Pen Shell (Pinna nobilis L. 1758) Mass Mortality Events in Adriatic Sea Is Characterised with Rapid Spreading and Acute Disease Progression" Pathogens 9, no. 10: 776. https://doi.org/10.3390/pathogens9100776
APA StyleŠarić, T., Župan, I., Aceto, S., Villari, G., Palić, D., De Vico, G., & Carella, F. (2020). Epidemiology of Noble Pen Shell (Pinna nobilis L. 1758) Mass Mortality Events in Adriatic Sea Is Characterised with Rapid Spreading and Acute Disease Progression. Pathogens, 9(10), 776. https://doi.org/10.3390/pathogens9100776