Genetic Basis and Physiological Effects of Lipid A Hydroxylation in Pseudomonas aeruginosa PAO1

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Generation of Deletion Mutants
2.3. Generation of Plasmids
2.4. Lipid A Analysis
2.5. Detergent Sensitivity Assay
2.6. Antibiotics Sensitivity Assays
2.7. Biofilm Assay
2.8. Whole Blood Killing Assay
2.9. Galleria Mellonella Infection and Persistence Assays
2.10. Statistical Analyses
3. Results and Discussion
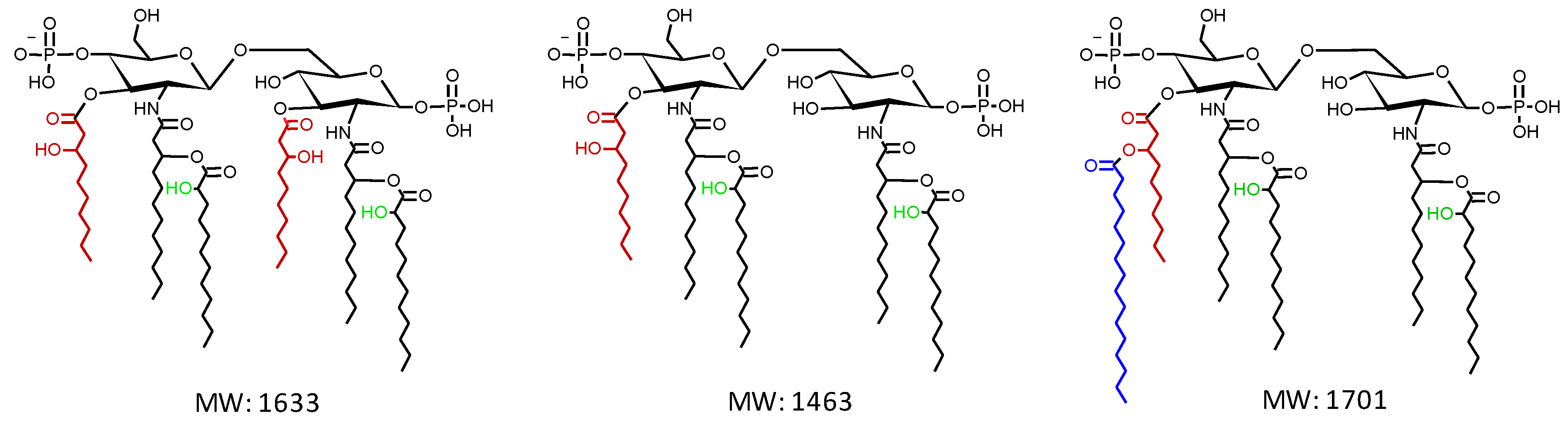
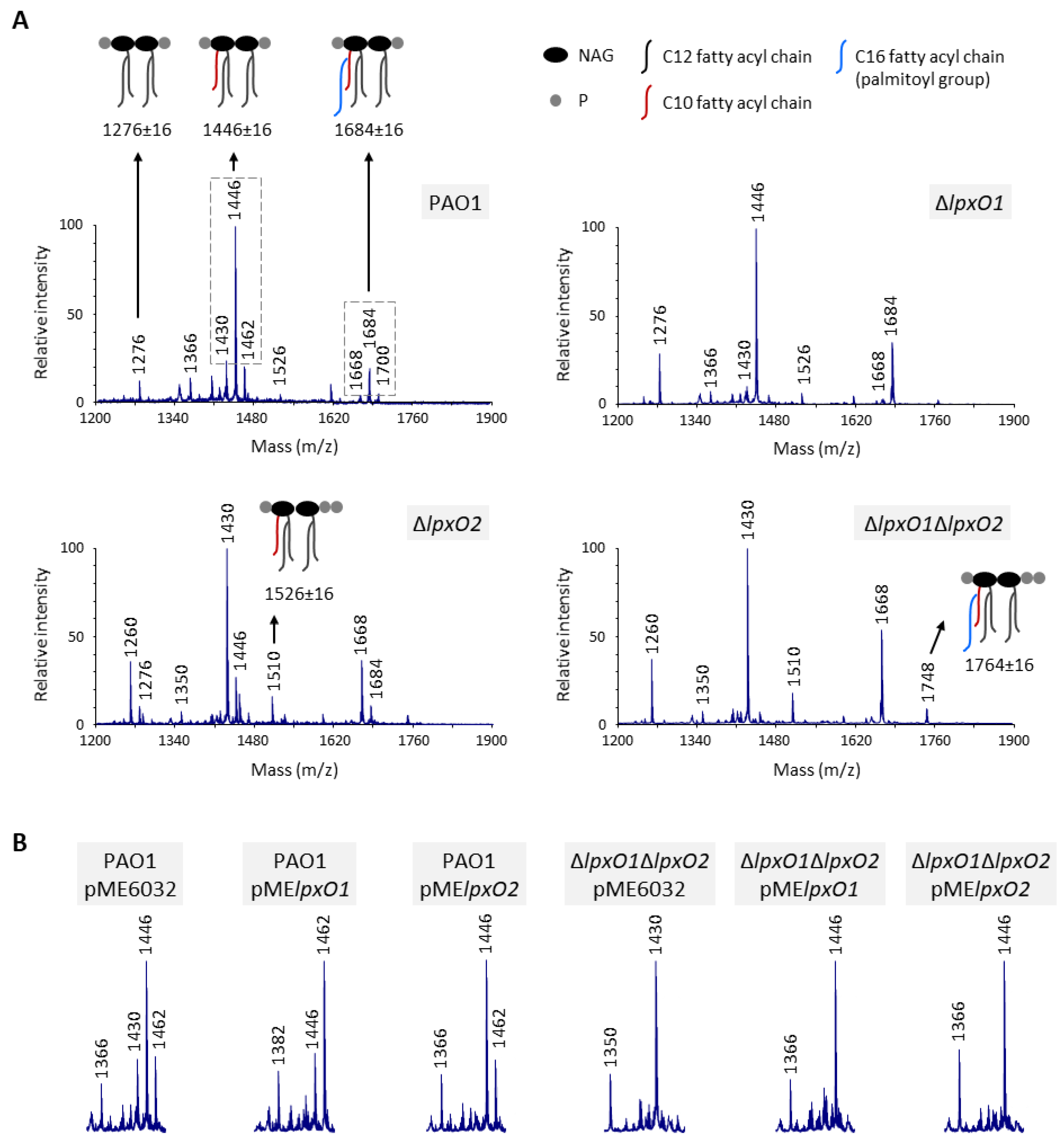
3.1. Both LpxO1 and LpxO2 are Responsible for Lipid A Hydroxylation
3.2. Lipid A Hydroxylation Does Not Affect in vitro Growth, Cell Envelope Integrity and Resistance
3.3. Lipid A Hydroxylation Contributes to P. aeruginosa Virulence in an Insect Model of Infection
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Silhavy, T.J.; Kahne, D.; Walker, S. The bacterial cell envelope. Cold Spring Harb. Perspect. Biol. 2010, 2, a000414. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef] [PubMed]
- Tümmler, B. Emerging therapies against infections with Pseudomonas aeruginosa. F1000Research 2019, 8, F1000. [Google Scholar] [CrossRef] [PubMed]
- King, J.D.; Kocíncová, D.; Westman, E.L.; Lam, J.S. Review: Lipopolysaccharide biosynthesis in Pseudomonas aeruginosa. Innate Immun. 2009, 15, 261–312. [Google Scholar] [CrossRef]
- Mdluli, K.E.; Witte, P.R.; Kline, T.; Barb, A.W.; Erwin, A.L.; Mansfield, B.E.; McClerren, A.L.; Pirrung, M.C.; Tumey, L.N.; Warrener, P.; et al. Molecular validation of LpxC as an antibacterial drug target in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2006, 50, 2178–2184. [Google Scholar] [CrossRef]
- Srinivas, N.; Jetter, P.; Ueberbacher, B.J.; Werneburg, M.; Zerbe, K.; Steinmann, J.; Van der Meijden, B.; Bernardini, F.; Lederer, A.; Dias, R.L.; et al. Peptidomimetic antibiotics target outer-membrane biogenesis in Pseudomonas aeruginosa. Science 2010, 327, 1010–1013. [Google Scholar] [CrossRef]
- Fernández-Piñar, R.; Lo Sciuto, A.; Rossi, A.; Ranucci, S.; Bragonzi, A.; Imperi, F. In vitro and in vivo screening for novel essential cell-envelope proteins in Pseudomonas aeruginosa. Sci. Rep. 2015, 5, 17593. [Google Scholar] [CrossRef]
- Knirel, Y.A.; Bystrova, O.V.; Kocharova, N.A.; Zähringer, U.; Pier, G.B. Conserved and variable structural features in the lipopolysaccharide of Pseudomonas aeruginosa. J. Endotoxin Res. 2006, 12, 324–336. [Google Scholar] [CrossRef]
- Ernst, R.K.; Adams, K.N.; Moskowitz, S.M.; Kraig, G.M.; Kawasaki, K.; Stead, C.M.; Trent, M.S.; Miller, S.I. The Pseudomonas aeruginosa lipid A deacylase: Selection for expression and loss within the cystic fibrosis airway. J. Bacteriol. 2006, 188, 191–201. [Google Scholar] [CrossRef]
- Lo Sciuto, A.; Martorana, A.M.; Fernández-Piñar, R.; Mancone, C.; Polissi, A.; Imperi, F. Pseudomonas aeruginosa LptE is crucial for LptD assembly, cell envelope integrity, antibiotic resistance and virulence. Virulence 2018, 9, 1718–1733. [Google Scholar] [CrossRef]
- Gibbons, H.S.; Lin, S.; Cotter, R.J.; Raetz, C.R. Oxygen requirement for the biosynthesis of the S-2-hydroxymyristate moiety in Salmonella typhimurium lipid A. Function of LpxO, A new Fe2+/alpha-ketoglutarate-dependent dioxygenase homologue. J. Biol. Chem. 2000, 275, 32940–32949. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, H.S.; Reynolds, C.M.; Guan, Z.; Raetz, C.R. An inner membrane dioxygenase that generates the 2-hydroxymyristate moiety of Salmonella lipid A. Biochemistry 2008, 47, 2814–2825. [Google Scholar] [CrossRef] [PubMed]
- Mills, G.; Dumigan, A.; Kidd, T.; Hobley, L.; Bengoechea, J.A. Identification and Characterization of Two Klebsiella pneumoniae lpxL Lipid A Late Acyltransferases and Their Role in Virulence. Infect. Immun. 2017, 85, e00068-17. [Google Scholar] [CrossRef] [PubMed]
- Bartholomew, T.L.; Kidd, T.J.; Sá Pessoa, J.; Conde Álvarez, R.; Bengoechea, J.A. 2-Hydroxylation of Acinetobacter baumannii Lipid A Contributes to Virulence. Infect. Immun. 2019, 87, e00066-19. [Google Scholar] [CrossRef] [PubMed]
- Norris, M.H.; Somprasong, N.; Schweizer, H.P.; Tuanyok, A. Lipid A Remodeling Is a Pathoadaptive Mechanism That Impacts Lipopolysaccharide Recognition and Intracellular Survival of Burkholderia pseudomallei. Infect. Immun. 2018, 86, e00360-18. [Google Scholar] [CrossRef] [PubMed]
- Llobet, E.; Martínez-Moliner, V.; Moranta, D.; Dahlström, K.M.; Regueiro, V.; Tomás, A.; Cano, V.; Pérez-Gutiérrez, C.; Frank, C.G.; Fernández-Carrasco, H.; et al. Deciphering tissue-induced Klebsiella pneumoniae lipid A structure. Proc. Natl. Acad. Sci. USA 2015, 112, E6369–E6378. [Google Scholar] [CrossRef] [PubMed]
- Rocchio, S.; Santorelli, D.; Rinaldo, S.; Franceschini, M.; Malatesta, F.; Imperi, F.; Federici, L.; Travaglini-Allocatelli, C.; Di Matteo, A. Structural and functional investigation of the Small Ribosomal Subunit Biogenesis GTPase A (RsgA) from Pseudomonas aeruginosa. FEBS J. 2019. [Google Scholar] [CrossRef]
- Milton, D.L.; O’Toole, R.; Horstedt, P.; Wolf-Watz, H. Flagellin A is essential for the virulence of Vibrio anguillarum. J. Bacteriol. 1996, 178, 1310–1319. [Google Scholar] [CrossRef]
- Hoang, T.T.; Karkhoff-Schweizer, R.R.; Kutchma, A.J.; Schweizer, H.P. A broad-host-range Flp-FRT recombination system for site-specific excision of chromosomally-located DNA sequences: Application for isolation of unmarked Pseudomonas aeruginosa mutants. Gene 1998, 212, 77–86. [Google Scholar] [CrossRef]
- Heeb, S.; Blumer, C.; Haas, D. Regulatory RNA as mediator in GacA/RsmA-dependent global control of exoproduct formation in Pseudomonas fluorescens CHA0. J Bacteriol. 2002, 184, 1046–1056. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Chandler, C.E.; Leung, L.M.; McElheny, C.L.; Mettus, R.T.; Shanks, R.M.Q.; Liu, J.H.; Goodlett, D.R.; Ernst, R.K.; Doi, Y. Structural Modification of Lipopolysaccharide Conferred by mcr-1 in Gram-Negative ESKAPE Pathogens. Antimicrob. Agents Chemother. 2017, 61, e00580-17. [Google Scholar] [CrossRef] [PubMed]
- Lo Sciuto, A.; Fernández-Piñar, R.; Bertuccini, L.; Iosi, F.; Superti, F.; Imperi, F. The periplasmic protein TolB as a potential drug target in Pseudomonas aeruginosa. PLoS ONE 2014, 9, e103784. [Google Scholar] [CrossRef] [PubMed]
- Jander, G.; Rahme, L.G.; Ausubel, F.M. Positive correlation between virulence of Pseudomonas aeruginosa mutants in mice and insects. J. Bacteriol. 2000, 182, 3843–3845. [Google Scholar] [CrossRef] [PubMed]
- Antunes, L.C.; Imperi, F.; Carattoli, A.; Visca, P. Deciphering the multifactorial nature of Acinetobacter baumannii pathogenicity. PLoS ONE 2011, 6, e22674. [Google Scholar] [CrossRef] [PubMed]
- Cigana, C.; Curcurù, L.; Leone, M.R.; Ieranò, T.; Lorè, N.I.; Bianconi, I.; Silipo, A.; Cozzolino, F.; Lanzetta, R.; Molinaro, A.; et al. Pseudomonas aeruginosa exploits lipid A and muropeptides modification as a strategy to lower innate immunity during cystic fibrosis lung infection. PLoS ONE 2009, 4, e8439. [Google Scholar] [CrossRef]
- Korneev, K.V.; Arbatsky, N.P.; Molinaro, A.; Palmigiano, A.; Shaikhutdinova, R.Z.; Shneider, M.M.; Pier, G.B.; Kondakova, A.N.; Sviriaeva, E.N.; Sturiale, L.; et al. Structural Relationship of the Lipid A Acyl Groups to Activation of Murine Toll-Like Receptor 4 by Lipopolysaccharides from Pathogenic Strains of Burkholderia mallei, Acinetobacter baumannii, and Pseudomonas aeruginosa. Front. Immunol. 2015, 6, 595. [Google Scholar] [CrossRef]
- Latino, L.; Caroff, M.; Pourcel, C. Fine structure analysis of lipopolysaccharides in bacteriophage-resistant Pseudomonas aeruginosa PAO1 mutants. Microbiology 2017, 163, 848–855. [Google Scholar] [CrossRef]
- Galani, I.; Kontopidou, F.; Souli, M.; Rekatsina, P.D.; Koratzanis, E.; Deliolanis, J.; Giamarellou, H. Colistin susceptibility testing by Etest and disk diffusion methods. Int. J. Antimicrob. Agents 2008, 31, 434–439. [Google Scholar] [CrossRef]
- Hankins, J.V.; Madsen, J.A.; Giles, D.K.; Childers, B.M.; Klose, K.E.; Brodbelt, J.S.; Trent, M.S. Elucidation of a novel Vibrio cholerae lipid A secondary hydroxy-acyltransferase and its role in innate immune recognition. Mol. Microbiol. 2011, 81, 1313–1329. [Google Scholar] [CrossRef]
- Termine, E.; Michel, G.P. Transcriptome and secretome analyses of the adaptive response of Pseudomonas aeruginosa to suboptimal growth temperature. Int. Microbiol. 2009, 12, 7–12. [Google Scholar]
- Redfern, J.; Wallace, J.; van Belkum, A.; Jaillard, M.; Whittard, E.; Ragupathy, R.; Verran, J.; Kelly, P.; Enright, M.C. Biofilm Associated Genotypes of Multidrug-Resistant Pseudomonas aeruginosa. bioRxiv 2019, 713453. [Google Scholar] [CrossRef]
- Tsai, C.J.; Loh, J.M.; Proft, T. Galleria mellonella infection models for the study of bacterial diseases and for antimicrobial drug testing. Virulence 2016, 7, 214–229. [Google Scholar] [CrossRef]
- Raetz, C.R.; Reynolds, C.M.; Trent, M.S.; Bishop, R.E. Lipid A modification systems in gram-negative bacteria. Annu. Rev. Biochem. 2007, 76, 295–329. [Google Scholar] [CrossRef] [PubMed]
- Steimle, A.; Autenrieth, I.B.; Frick, J.S. Structure and function: Lipid A modifications in commensals and pathogens. Int. J. Med. Microbiol. 2016, 306, 290–301. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, R.F.; Sá-Correia, I.; Valvano, M.A. Lipopolysaccharide modification in Gram-negative bacteria during chronic infection. FEMS Microbiol. Rev. 2016, 40, 480–493. [Google Scholar] [CrossRef] [PubMed]
- Anandan, A.; Vrielink, A. Structure and function of lipid A-modifying enzymes. Ann. N. Y. Acad. Sci. 2019. [Google Scholar] [CrossRef]
- Thaipisuttikul, I.; Hittle, L.E.; Chandra, R.; Zangari, D.; Dixon, C.L.; Garrett, T.A.; Rasko, D.A.; Dasgupta, N.; Moskowitz, S.M.; Malmström, L.; et al. A divergent Pseudomonas aeruginosa palmitoyltransferase essential for cystic fibrosis-specific lipid A. Mol. Microbiol. 2014, 91, 158–174. [Google Scholar] [CrossRef]
- Nowicki, E.M.; O’Brien, J.P.; Brodbelt, J.S.; Trent, M.S. Characterization of Pseudomonas aeruginosa LpxT reveals dual positional lipid A kinase activity and co-ordinated control of outer membrane modification. Mol. Microbiol. 2014, 94, 728–741. [Google Scholar] [CrossRef]
- Hittle, L.E.; Powell, D.A.; Jones, J.W.; Tofigh, M.; Goodlett, D.R.; Moskowitz, S.M.; Ernst, R.K. Site-specific activity of the acyltransferases HtrB1 and HtrB2 in Pseudomonas aeruginosa lipid A biosynthesis. Pathog. Dis. 2015, 73, ftv053. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Inhibition Halo (mm) 2 | MIC (μg/ml) 3 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Cip | Novo | Rif | Ery | Sm | Tob | Imp | Ct | Ct | |
| PAO1 | 30.6 (±0.5) | 6.0 (±0.0) | 6.0 (±0.0) | 6.0 (±0.0) | 12.9 (±1.3) | 22.2 (±1.7) | 25.0 (±1.4) | 12.5 (±0.5) | 0.5 |
| ΔlpxO1 | 30.8 (±1.6) | 6.0 (±0.0) | 6.0 (±0.0) | 6.0 (±0.0) | 12.6 (±0.7) | 23.2 (±3.1) | 24.2 (±0.5) | 12.5 (±0.5) | 0.5 |
| ΔlpxO2 | 30.9 (±1.1) | 6.0 (±0.0) | 6.0 (±0.0) | 6.0 (±0.0) | 11.9 (±1.5) | 22.0 (±0.9) | 25.2 (±0.8) | 12.5 (±0.5) | 0.5 |
| ΔlpxO1ΔlpxO2 | 30.8 (±2.2) | 6.0 (±0.0) | 6.0 (±0.0) | 6.0 (±0.0) | 12.4 (±1.8) | 23.0 (±2.8) | 24.4 (±1.2) | 12.5 (±0.5) | 0.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lo Sciuto, A.; Cervoni, M.; Stefanelli, R.; Spinnato, M.C.; Di Giamberardino, A.; Mancone, C.; Imperi, F. Genetic Basis and Physiological Effects of Lipid A Hydroxylation in Pseudomonas aeruginosa PAO1. Pathogens 2019, 8, 291. https://doi.org/10.3390/pathogens8040291
Lo Sciuto A, Cervoni M, Stefanelli R, Spinnato MC, Di Giamberardino A, Mancone C, Imperi F. Genetic Basis and Physiological Effects of Lipid A Hydroxylation in Pseudomonas aeruginosa PAO1. Pathogens. 2019; 8(4):291. https://doi.org/10.3390/pathogens8040291
Chicago/Turabian StyleLo Sciuto, Alessandra, Matteo Cervoni, Roberta Stefanelli, Maria Concetta Spinnato, Alessandra Di Giamberardino, Carmine Mancone, and Francesco Imperi. 2019. "Genetic Basis and Physiological Effects of Lipid A Hydroxylation in Pseudomonas aeruginosa PAO1" Pathogens 8, no. 4: 291. https://doi.org/10.3390/pathogens8040291
APA StyleLo Sciuto, A., Cervoni, M., Stefanelli, R., Spinnato, M. C., Di Giamberardino, A., Mancone, C., & Imperi, F. (2019). Genetic Basis and Physiological Effects of Lipid A Hydroxylation in Pseudomonas aeruginosa PAO1. Pathogens, 8(4), 291. https://doi.org/10.3390/pathogens8040291