Phenotypic Characterization of Rhodococcus equi Biofilm Grown In Vitro and Inhibiting and Dissolving Activity of Azithromycin/Rifampicin Treatment

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Results
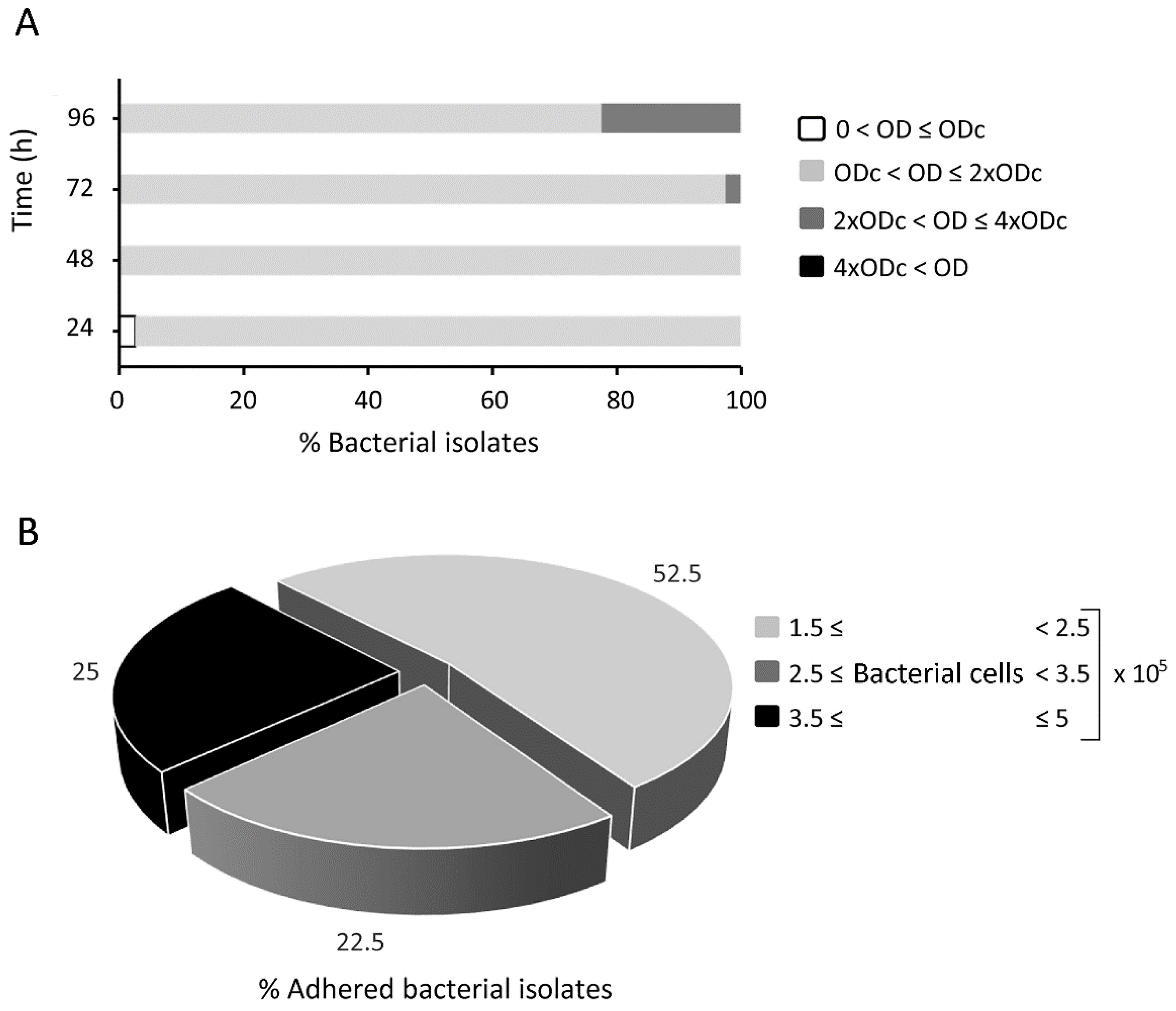
2.1. Biofilm Formation and Cellular Adhesion of Fluorescein-Labelled Rhodococcus equi
2.2. Minimum Inhibitory Concentration of Antibiotics against Rhodococcus equi Isolates
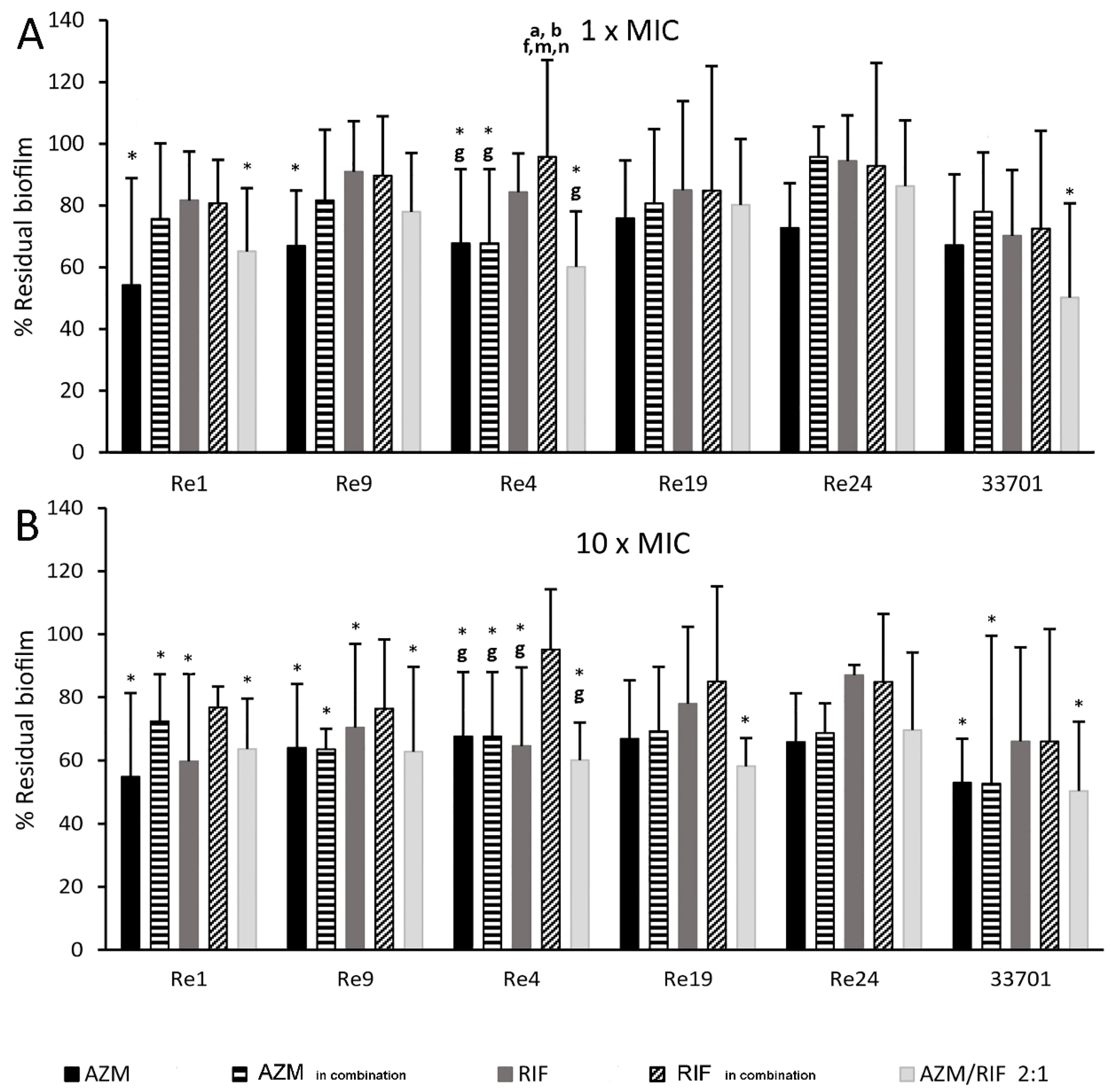
2.3. Long-Term Antimicrobial Prevention of Rhodococcus equi Biofilm
2.4. Kinetic Dissolution of Pre-Formed Rhodococcus equi Biofilm
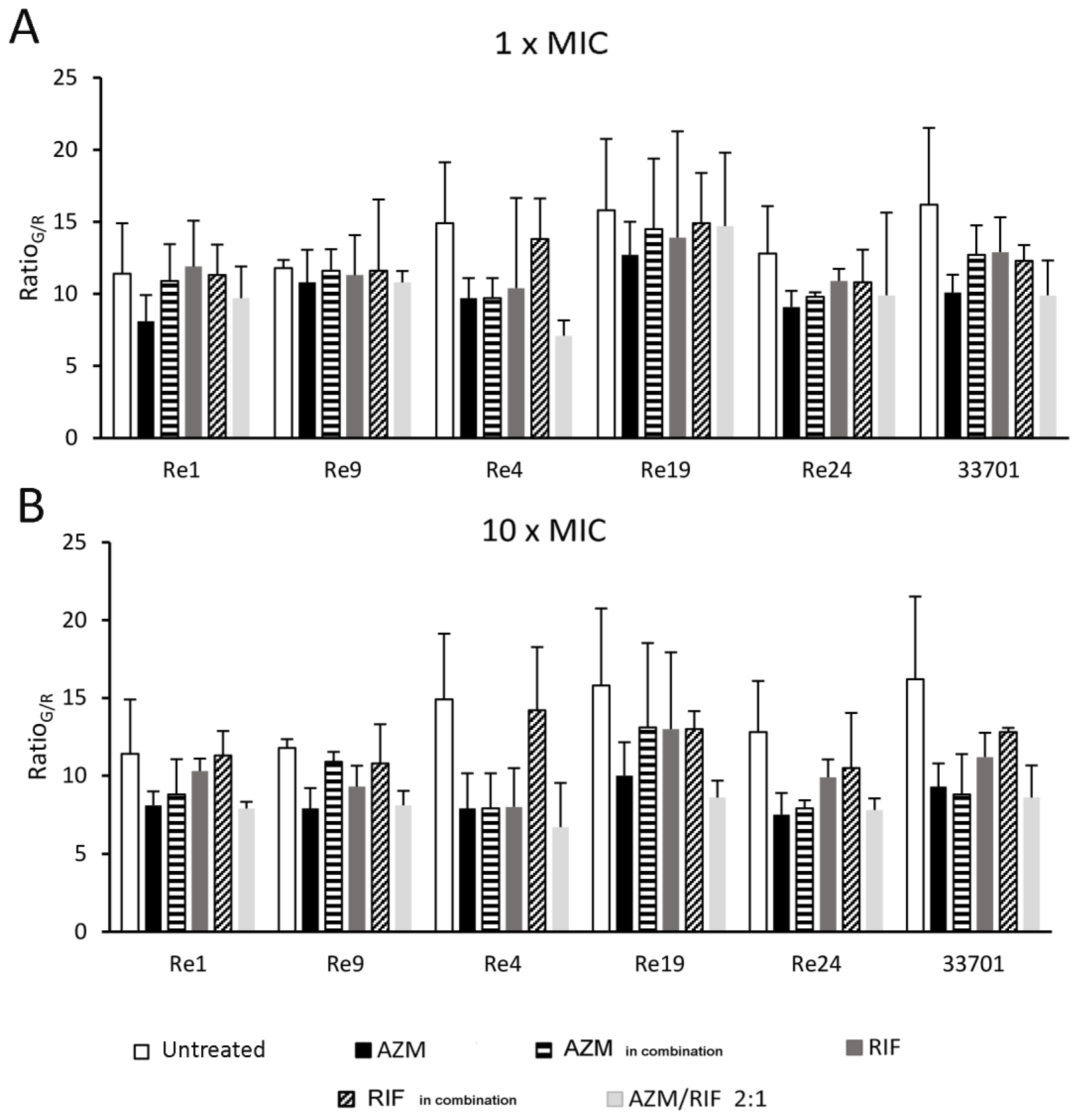
2.5. Biofilm Fluorescence Measurement
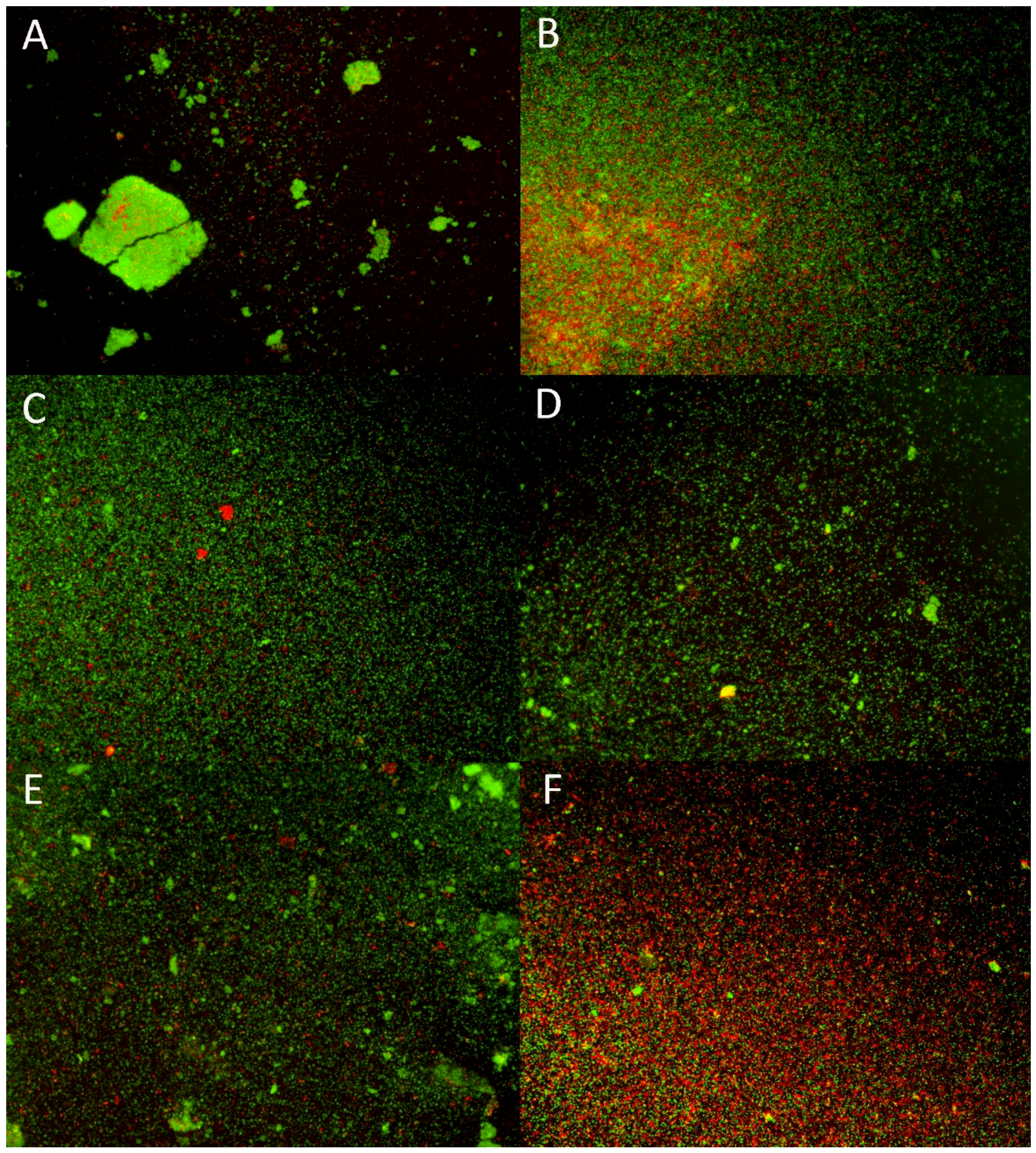
2.6. Structure of Rhodococcus equi Biofilms Captured by Scanning Electron Microscope
3. Discussion
3.1. In Vitro Model of Rhodococcus equi Biofilm Growth
3.2. Putative Pathogenic Significance of Rhodococcus equi Adhesion to Lung Alveolar Epithelium
3.3. Macrolides and Rifampicin versus Biofilm: A Controversial Issue
3.4. Ultramicroscopic Biofilm Phenotype-Based Assumptions
4. Conclusions
5. Materials and Methods
5.1. Bacteria Identification
5.2. Spray-Dried Antibiotic Microparticles
5.3. Antimicrobial Efficacy against Planktonic Bacteria
5.4. Fluorescence-Based Adhesion Assay on Pulmonary Alveolar Epithelium
5.4.1. Bacteria Labelling
5.4.2. Rhodococcus equi Adhesion Assay
5.5. Kinetic Growth of Rhodococcus equi Biofilm and Inhibiting/Dissolving Capacity of the Antimicrobial Treatments
5.6. Morphological Characterization of Rhodococcus equi Biofilm
5.6.1. LIVE/DEAD Biofilm Viability Assay and Fluorescence Microscopy
5.6.2. Scanning Electron Microscopy
5.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Costerton, W.; Veeh, R.; Shirtliff, M.; Pasmore, M.; Post, C.; Ehrlich, G. The application of biofilm science to the study and control of chronic bacterial infections. J. Clin. Investig. 2003, 112, 1466–1477. [Google Scholar] [CrossRef]
- Ojha, A.K.; Baughn, A.D.; Sambandan, D.; Hsu, T.; Trivelli, X.; Guerardel, Y.; Alahari, A.; Kremer, L.; Jacobs, W.R.; Hatfull, G.F. Growth of Mycobacterium tuberculosis biofilms containing free mycolic acids and harbouring drug-tolerant bacteria. Mol. Microbiol. 2008, 69, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Al Akhrass, F.; Al Wohoush, I.; Chaftari, A.-M.; Reitzel, R.; Jiang, Y.; Ghannoum, M.; Tarrand, J.; Hachem, R.; Raad, I. Rhodococcus bacteremia in cancer patients is mostly catheter related and associated with biofilm formation. PLoS ONE 2012, 7, e32945. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.H.; Cunha, B.A. Rhodococcus equi pneumonia. Semin. Respir. Infect. 1997, 12, 57–60. [Google Scholar] [PubMed]
- Asoh, N.; Watanabe, H.; Fines-Guyon, M.; Watanabe, K.; Oishi, K.; Kositsakulchai, W.; Sanchai, T.; Kunsuikmengrai, K.; Kahintapong, S.; Khantawa, B.; et al. Emergence of rifampin-resistant Rhodococcus equi with several types of mutations in the rpoB gene among AIDS patients in northern Thailand. J. Clin. Microbiol. 2003, 41, 2337–2340. [Google Scholar] [CrossRef]
- Gray, K.J.; French, N.; Lugada, E.; Watera, C.; Gilks, C.F. Rhodococcus equi and HIV-1 infection in Uganda. J. Infect. 2000, 41, 227–231. [Google Scholar] [CrossRef]
- Herath, S.; Lewis, C.; Nisbet, M. Increasing awareness of Rhodococcus equi pulmonary infection in the immunocompetent adult: A rare infection with poor prognosis. N. Z. Med. J. 2013, 126, 165–174. [Google Scholar] [PubMed]
- Hulsewé-Evers, H.P.; Jansveld, C.A.; Jansz, A.R.; Schneider, M.M.; Bravenboer, B. HIV-infected patient with a Rhodococcus equi pneumonia. Neth. J. Med. 2000, 57, 25–29. [Google Scholar] [CrossRef]
- Khan, M.Y.; Ali, S.; Baqi, S. Rhodococcus equi pneumonia in a live related renal transplant recipient. J. Pak. Med. Assoc. 2013, 63, 635–638. [Google Scholar]
- Mizuno, Y.; Sato, F.; Sakamoto, M.; Yoshikawa, K.; Yoshida, M.; Shiba, K.; Onodera, S.; Matsuura, R.; Takai, S. VapB-positive Rhodococcus equi infection in an HIV-infected patient in Japan. J. Infect. Chemother. 2005, 11, 37–40. [Google Scholar] [CrossRef]
- Weinstock, D.M.; Brown, A.E. Rhodococcus equi: An emerging pathogen. Clin. Infect. Dis. 2002, 34, 1379–1385. [Google Scholar] [CrossRef] [PubMed]
- von Bargen, K.; Haas, A. Molecular and infection biology of the horse pathogen Rhodococcus equi. FEMS Microbiol. Rev. 2009, 33, 870–891. [Google Scholar] [CrossRef] [PubMed]
- Muscatello, G.; Gilkerson, J.R.; Browning, G.F. Detection of virulent Rhodococcus equi in exhaled air samples from naturally infected foals. J. Clin. Microbiol. 2009, 47, 734–737. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.; Sowden, D.; Caffery, M.; Bint, M.; Broom, J. Rhodococcus equi infection: A diverse spectrum of disease. IDCases 2019, 15, e00487. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.V.; Kruse, R.L.; Yang, K.; Musher, D.M. Diagnosis and Management of Pulmonary Infection due to Rhodococcus equi. Clin. Microbiol. Infect. 2019, 25, 310–315. [Google Scholar] [CrossRef]
- Witkowski, L.; Rzewuska, M.; Takai, S.; Kizerwetter-Świda, M.; Kita, J. Molecular epidemiology of Rhodococcus equi in slaughtered swine, cattle and horses in Poland. BMC Microbiol. 2016, 16, 98. [Google Scholar] [CrossRef] [PubMed]
- Ocampo-Sosa, A.A.; Lewis, D.A.; Navas, J.; Quigley, F.; Callejo, R.; Scortti, M.; Leadon, D.P.; Fogarty, U.; Vázquez-Boland, J.A. Molecular Epidemiology of Rhodococcus equi Based on traA, vapA, and vapB Virulence Plasmid Markers. J. Infect. Dis. 2007, 196, 763–769. [Google Scholar] [CrossRef]
- Vechi, H.T.; de Oliveira, E.T.G.; de Freitas, M.R.; Rossi, F.; de Britto, M.H.M.F.; Alves, M.D.M. Chronic cavitary pneumonia by Rhodococcus equi in a highly prevalent tuberculosis country: A diagnosis challenge. Rev. Inst. Med. Trop. Sao Paulo 2018, 60, e74. [Google Scholar] [CrossRef]
- Bujold, A.R.; Lani, N.R.; Sanz, M.G. Strain-to-strain variation of Rhodococcus equi growth and biofilm formation in vitro. BMC Res. Notes 2019, 12, 519. [Google Scholar] [CrossRef]
- Gressler, L.T.; de Vargas, A.C.; da Costa, M.M.; Sutili, F.J.; Schwab, M.; Pereira, D.I.B.; Sangioni, L.A.; de A. Botton, S. Biofilm formation by Rhodococcus equi and putative association with macrolide resistance. Pesqui. Veterinária Bras. 2015, 35, 835–841. [Google Scholar] [CrossRef]
- Petrova, O.E.; Sauer, K. Sticky situations: Key components that control bacterial surface attachment. J. Bacteriol. 2012, 194, 2413–2425. [Google Scholar] [CrossRef] [PubMed]
- Orme, I.M. A new unifying theory of the pathogenesis of tuberculosis. Tuberc. (Edinb) 2014, 94, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Giguère, S. Treatment of Infections Caused by Rhodococcus equi. Vet. Clin. North Am. Equine Pr. 2017, 33, 67–85. [Google Scholar] [CrossRef] [PubMed]
- Polonio, R.E.; Mermel, L.A.; Paquette, G.E.; Sperry, J.F. Eradication of biofilm-forming Staphylococcus epidermidis (RP62A) by a combination of sodium salicylate and vancomycin. Antimicrob. Agents Chemother. 2001, 45, 3262–3266. [Google Scholar] [CrossRef] [PubMed]
- Aka, S.T.; Haji, S.H. Sub-MIC of antibiotics induced biofilm formation of Pseudomonas aeruginosa in the presence of chlorhexidine. Braz. J. Microbiol. 2015, 46, 149–154. [Google Scholar] [CrossRef]
- Rampacci, E.; Marenzoni, M.L.; Chiaradia, E.; Passamonti, F.; Ricci, M.; Pepe, M.; Coletti, M.; Giovagnoli, S. In vitro performances of novel co-spray-dried azithromycin/rifampicin microparticles for Rhodococcus equi disease treatment. Sci. Rep. 2018, 8, 12149. [Google Scholar] [CrossRef]
- Prescott, J.F. Rhodococcus equi: An animal and human pathogen. Clin. Microbiol. Rev. 1991, 4, 20–34. [Google Scholar] [CrossRef]
- Bjarnsholt, T.; Alhede, M.; Alhede, M.; Eickhardt-Sørensen, S.R.; Moser, C.; Kühl, M.; Jensen, P.Ø.; Høiby, N. The in vivo biofilm. Trends Microbiol. 2013, 21, 466–474. [Google Scholar] [CrossRef]
- Lebeaux, D.; Chauhan, A.; Rendueles, O.; Beloin, C. From in vitro to in vivo Models of Bacterial Biofilm-Related Infections. Pathogens 2013, 2, 288–356. [Google Scholar] [CrossRef]
- Sadowska, B.; Bonar, A.; von Eiff, C.; Proctor, R.A.; Chmiela, M.; Rudnicka, W.; Róźalska, B. Characteristics of Staphylococcus aureus, isolated from airways of cystic fibrosis patients, and their small colony variants. FEMS Immunol. Med. Microbiol. 2002, 32, 191–197. [Google Scholar] [CrossRef]
- Boisvert, A.-A.; Cheng, M.P.; Sheppard, D.C.; Nguyen, D. Microbial Biofilms in Pulmonary and Critical Care Diseases. Ann. Am. Thorac. Soc. 2016, 13, 1615–1623. [Google Scholar] [CrossRef] [PubMed]
- Sanz, M.; Loynachan, A.; Sun, L.; Oliveira, A.; Breheny, P.; Horohov, D.W. The effect of bacterial dose and foal age at challenge on Rhodococcus equi infection. Vet. Microbiol. 2013, 167, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Huesca, M.; Peralta, R.; Sauder, D.N.; Simor, A.E.; McGavin, M.J. Adhesion and virulence properties of epidemic Canadian methicillin-resistant Staphylococcus aureus strain 1: Identification of novel adhesion functions associated with plasmin-sensitive surface protein. J. Infect. Dis. 2002, 185, 1285–1296. [Google Scholar] [CrossRef] [PubMed]
- Piqué, N.; Berlanga, M.; Miñana-Galbis, D. Health Benefits of Heat-Killed (Tyndallized) Probiotics: An Overview. Int. J. Mol. Sci. 2019, 20, 2534. [Google Scholar] [CrossRef] [PubMed]
- Nucleo, E.; Steffanoni, L.; Fugazza, G.; Migliavacca, R.; Giacobone, E.; Navarra, A.; Pagani, L.; Landini, P. Growth in glucose-based medium and exposure to subinhibitory concentrations of imipenem induce biofilm formation in a multidrug-resistant clinical isolate of Acinetobacter baumannii. BMC Microbiol. 2009, 9, 270. [Google Scholar] [CrossRef]
- Hoffman, L.R.; D’Argenio, D.A.; MacCoss, M.J.; Zhang, Z.; Jones, R.A.; Miller, S.I. Aminoglycoside antibiotics induce bacterial biofilm formation. Nature 2005, 436, 1171–1175. [Google Scholar] [CrossRef]
- Gillis, R.J.; Iglewski, B.H. Azithromycin Retards Pseudomonas aeruginosa Biofilm Formation. J. Clin. Microbiol. 2004, 42, 5842–5845. [Google Scholar] [CrossRef]
- Lima-e-Silva, A.A.; Silva-Filho, R.G.; Fernandes, H.M.Z.; Saramago, C.S.M.; Viana, A.S.; Souza, M.J.; Nogueira, E.M. Sub-Inhibitory Concentrations of Rifampicin Strongly Stimulated Biofilm Production in S. aureus. Open Microbiol. J. 2017, 11, 142–151. [Google Scholar] [CrossRef][Green Version]
- Wu, H.; Moser, C.; Wang, H.-Z.; Høiby, N.; Song, Z.-J. Strategies for combating bacterial biofilm infections. Int. J. Oral Sci. 2015, 7, 1–7. [Google Scholar] [CrossRef]
- Presterl, E.; Hajdu, S.; Lassnigg, A.M.; Hirschl, A.M.; Holinka, J.; Graninger, W. Effects of Azithromycin in Combination with Vancomycin, Daptomycin, Fosfomycin, Tigecycline, and Ceftriaxone on Staphylococcus epidermidis Biofilms. Antimicrob. Agents Chemother. 2009, 53, 3205–3210. [Google Scholar] [CrossRef]
- Croes, S.; Beisser, P.S.; Neef, C.; Bruggeman, C.A.; Stobberingh, E.E. Unpredictable Effects of Rifampin as an Adjunctive Agent in Elimination of Rifampin-Susceptible and -Resistant Staphylococcus aureus Strains Grown in Biofilms. Antimicrob. Agents Chemother. 2010, 54, 3907–3912. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Stiefel, P.; Schmidt-Emrich, S.; Maniura-Weber, K.; Ren, Q. Critical aspects of using bacterial cell viability assays with the fluorophores SYTO9 and propidium iodide. BMC Microbiol. 2015, 15, 36. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Visurraga, J.; Cárdenas, G.; García, A. Morphological changes induced in bacteria as evaluated by electron microscopy. In Microscopy: Science, Technology, Applications and Education; Méndez-Vilas, A., Díaz, J., Eds.; 4th series; Formatex Microscopy: Badajoz, Spain, 2010; pp. 307–315. [Google Scholar]
- Justice, S.S.; Hunstad, D.A.; Cegelski, L.; Hultgren, S.J. Morphological plasticity as a bacterial survival strategy. Nat. Rev. Microbiol. 2008, 6, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Giovagnoli, S.; Pietrella, D.; Barberini, L.; Santi, C.; Carotti, A.; di Michele, A.; Ricci, M. Reshaping antibiotics through hydrophobic drug-bile acid ionic complexation enhances activity against Staphylococcus aureus biofilms. Int. J. Pharm. 2017, 528, 144–162. [Google Scholar] [CrossRef]
- Sellon, D.C.; Besser, T.E.; Vivrette, S.L.; McConnico, R.S. Comparison of nucleic acid amplification, serology, and microbiologic culture for diagnosis of Rhodococcus equi pneumonia in foals. J. Clin. Microbiol. 2001, 39, 1289–1293. [Google Scholar] [CrossRef]
- Marenzoni, M.L.; Coppola, G.; Cappelli, K.; Capomaccio, S.; Felicetti, M.; Cercone, M.; Tamantini, C.; Genga, E.; Lepri, E.; Coletti, M.; et al. Development of a nested PCR for the diagnosis of Rhodococcus equi infection. Ippologia 2011, 22, 13–18. [Google Scholar]
- Clinical and Laboratory Standards Institute. Susceptibility Testing of Mycobacteria, Nocardiae, and Other Aerobic Actinomycetes, 2nd ed.; CLSI: Wayne, PA, USA, 2011. [Google Scholar]
- Hirschhausen, N.; Block, D.; Bianconi, I.; Bragonzi, A.; Birtel, J.; Lee, J.C.; Dübbers, A.; Küster, P.; Kahl, J.; Peters, G.; et al. Extended Staphylococcus aureus persistence in cystic fibrosis is associated with bacterial adaptation. Int. J. Med. Microbiol. 2013, 303, 685–692. [Google Scholar] [CrossRef]
- Stepanovic, S.; Vukovic, D.; Dakic, I.; Savic, B.; Svabic-Vlahovic, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | AZM | RIF | AZM/RIF 2:1 | Bacteria | AZM | RIF | AZM/RIF 2:1 |
|---|---|---|---|---|---|---|---|
| Re1 | 2 | 0.125 | 0.125/0.06 | Re21 | 2 | 0.06 | 0.125/0.06 |
| Re2 | 1 | 64 | 1/0.5 | Re22 | 1 | 0.125 | 0.25/0.125 |
| Re3 | 2 | 0.06 | 0.06/0.03 | Re23 | 2 | 0.25 | 0.25/0.125 |
| Re4 | 1 | 128 | 1.0/0.5 | Re24 | 1 | 0.125 | 0.125/0.06 |
| Re5 | 1 | 0.25 | 0.25/0.125 | Re25 | 1 | 0.125 | 0.25/0.125 |
| Re6 | 1 | 0.125 | 0.25/0.125 | Re26 | 1 | 0.125 | 0.125/0.06 |
| Re7 | 1 | 0.03 | 0.06/0.03 | Re27 | 1 | 0.25 | 0.25/0.125 |
| Re8 | 1 | 0.06 | 0.06/0.03 | Re28 | 2 | 0.25 | 0.5/0.25 |
| Re9 | 1 | 0.25 | 0.25/0.125 | Re29 | 2 | 0.06 | 0.125/0.06 |
| Re10 | 2 | 0.06 | 0.125/0.06 | Re30 | 1 | 0.06 | 0.125/0.06 |
| Re11 | 1 | 0.25 | 0.25/0.125 | Re31 | 1 | 0.06 | 0.125/0.06 |
| Re12 | 2 | 0.125 | 0.25/0.125 | Re32 | 2 | 0.125 | 0.25/0.125 |
| Re13 | 2 | 0.125 | 0.25/0.125 | Re33 | 1 | 0.06 | 0.125/0.06 |
| Re14 | 2 | 0.25 | 0.25/0.125 | Re34 | 1 | 0.125 | 0.25/0.125 |
| Re15 | 2 | 0.125 | 0.25/0.125 | Re35 | 1 | 0.06 | 0.125/0.06 |
| Re16 | 1 | 0.125 | 0.25/0.125 | Re36 | 1 | 0.03 | 0.06/0.03 |
| Re17 | 1 | 0.25 | 0.25/0.125 | Re37 | 1 | 0.03 | 0.06/0.03 |
| Re18 | 2 | 0.015 | 0.03/0.015 | Re38 | 1 | 0.125 | 0.25/0.125 |
| Re19 | 1 | 0.125 | 0.125/0.06 | Re39 | 2 | 0.25 | 0.25/0.125 |
| Re20 | 1 | 0.125 | 0.125/0.06 | 33701 | 0.5 | 0.125 | 0.125/0.06 |
| Re1 | Re9 | Re4 | Re19 | Re24 | 33701 | |
|---|---|---|---|---|---|---|
| AZM MIC | 38.2 *,b,c,d | 53.6 *,b,c,d | 48.5 *,d | 49 * | 69.1 | 44.6 *,b,c,d |
| AZM MICin combination | 72.7 a | 78.6 a | 48.5 *,d | 80.7 | 81.1 | 98.9 a |
| RIF MIC | 78.6 a | 78.3 a | 49.9 *,d | 84.5 | 91.7 | 97.8 a |
| RIF MICin combination | 84.8 a,e | 94.1 a,e | 88.1 a,b,c,e | 95.6 | 94.1 | 98.9 a |
| AZM/RIF 2:1 MIC | 53.2 *,d | 69.4 *,d | 48.4 *,d | 72 | 84.4 | 59.4 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rampacci, E.; Marenzoni, M.L.; Giovagnoli, S.; Passamonti, F.; Coletti, M.; Pietrella, D. Phenotypic Characterization of Rhodococcus equi Biofilm Grown In Vitro and Inhibiting and Dissolving Activity of Azithromycin/Rifampicin Treatment. Pathogens 2019, 8, 284. https://doi.org/10.3390/pathogens8040284
Rampacci E, Marenzoni ML, Giovagnoli S, Passamonti F, Coletti M, Pietrella D. Phenotypic Characterization of Rhodococcus equi Biofilm Grown In Vitro and Inhibiting and Dissolving Activity of Azithromycin/Rifampicin Treatment. Pathogens. 2019; 8(4):284. https://doi.org/10.3390/pathogens8040284
Chicago/Turabian StyleRampacci, Elisa, Maria Luisa Marenzoni, Stefano Giovagnoli, Fabrizio Passamonti, Mauro Coletti, and Donatella Pietrella. 2019. "Phenotypic Characterization of Rhodococcus equi Biofilm Grown In Vitro and Inhibiting and Dissolving Activity of Azithromycin/Rifampicin Treatment" Pathogens 8, no. 4: 284. https://doi.org/10.3390/pathogens8040284
APA StyleRampacci, E., Marenzoni, M. L., Giovagnoli, S., Passamonti, F., Coletti, M., & Pietrella, D. (2019). Phenotypic Characterization of Rhodococcus equi Biofilm Grown In Vitro and Inhibiting and Dissolving Activity of Azithromycin/Rifampicin Treatment. Pathogens, 8(4), 284. https://doi.org/10.3390/pathogens8040284