Aggregatibacter actinomycetemcomitans Biofilm Reduces Gingival Epithelial Cell Keratin Expression in an Organotypic Gingival Tissue Culture Model


Abstract
1. Introduction
2. Results
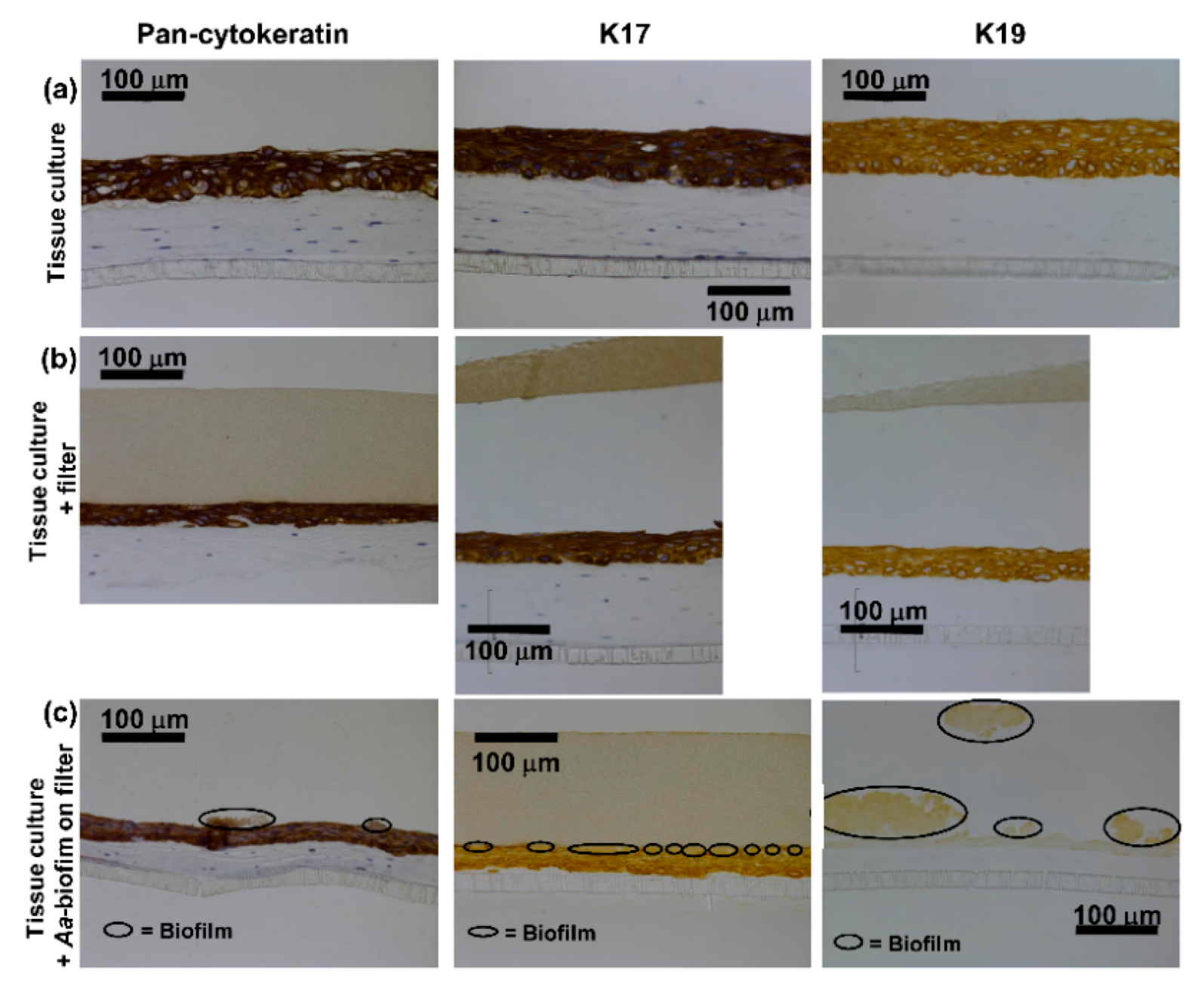
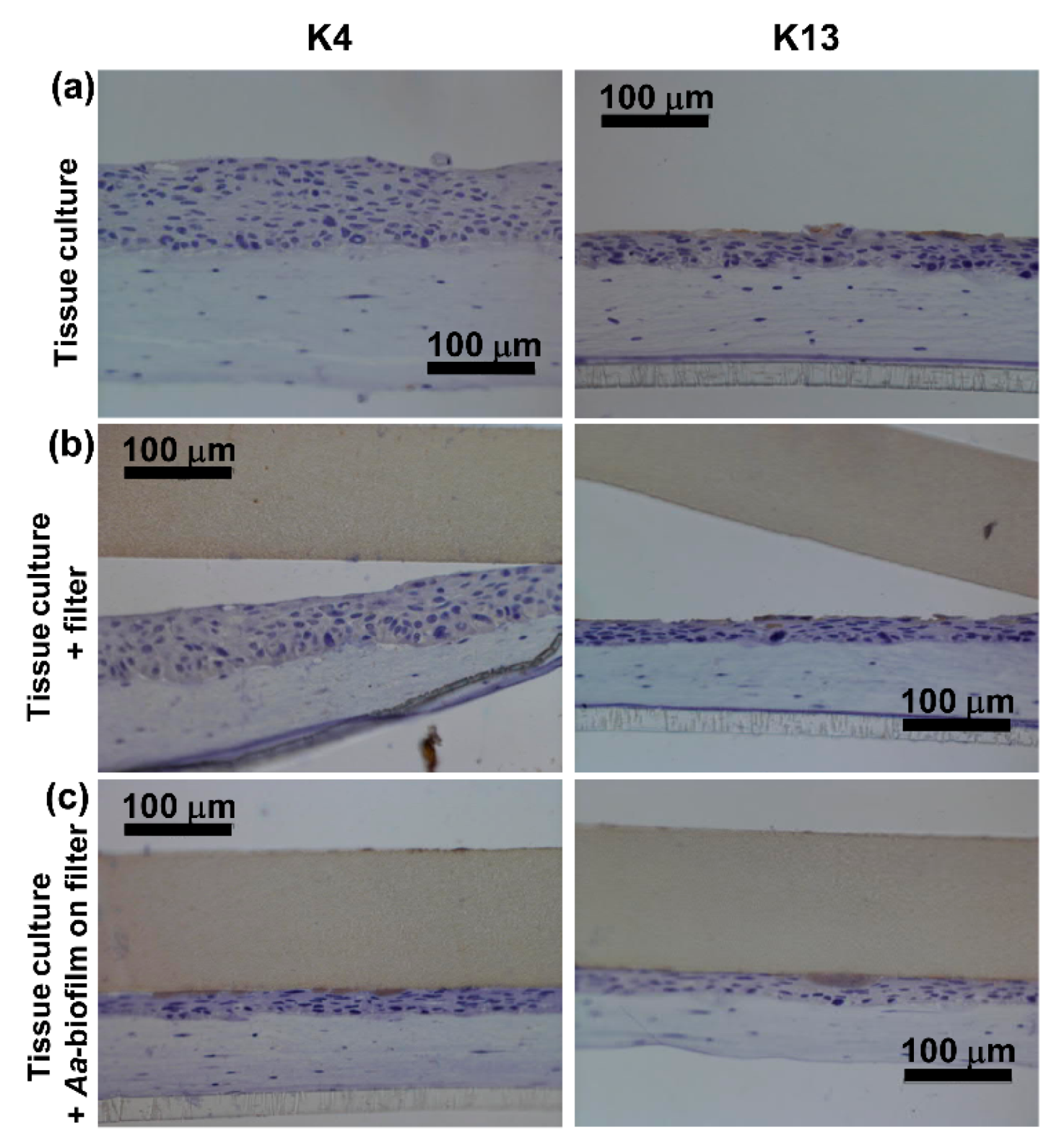
2.1. Control Cultures Showed Strong Expression of K17 and K19 and Only Weak or No Expression of K4 and K13
2.2. A. actinomycetemcomitans Biofilm Decreased the Expression of Keratin 17 and 19, Which Was in Accordance with the Decreased Expression of Total Keratin
2.3. A. actinomycetemcomitans Biofilm Caused Necrosis of the Epithelial Surface
3. Discussion
4. Materials and Methods
4.1. A. actinomycetemcomitans Biofilm Culture
4.2. Gingival Mucosa Co-Culture Models
4.3. Immunohistochemical Staining of Keratins K4, K13, K17, and K19
4.4. Transmission Electron Microscopic (TEM) Studies of the Biofilm-Epithelium Contact Site
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lamont, R.J.; Koo, H.; Hajishengallis, G. The oral microbiota: Dynamic communities and host interactions. Nat. Rev. Microbiol. 2018, 16, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Pöllänen, M.T.; Salonen, J.I. Effect of short chain fatty acids on human gingival epithelial cell keratins in vitro. Eur. J. Oral Sci. 2000, 108, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, I.C.; Gao, Z. Patterns of cytokeratin expression in the epithelia of inflamed human gingiva and periodontal pockets. J. Periodontal Res. 1993, 28, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Kebschull, M.; Guarnieri, P.; Demmer, R.T.; Boulesteix, A.L.; Pavlidis, P.; Papapanou, P.N. Molecular differences between chronic and aggressive periodontitis. J. Dent. Res. 2013, 92, 1081–1088. [Google Scholar] [CrossRef] [PubMed]
- Pöllänen, M.T.; Salonen, J.I.; Uitto, V.J. Structure and function of the tooth-epithelial interface in health and disease. Periodontol 2000 2003, 31, 12–31. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Yu, Y.; Ruan, H.; Luo, Y.; Guo, X. Morphological and functional characteristics of human gingival junctional epithelium. BMC Oral Health 2014, 14, 30. [Google Scholar] [CrossRef] [PubMed]
- Nagarakanti, S.; Ramya, S.; Babu, P.; Arun, K.V.; Sudarsan, S. Differential expression of E-cadherin and cytokeratin 19 and net proliferative rate of gingival keratinocytes in oral epithelium in periodontal health and disease. J. Periodontol. 2007, 78, 2197–2202. [Google Scholar] [CrossRef]
- Johnson, J.D.; Chen, R.; Lenton, P.A.; Zhang, G.; Hinrichs, J.E.; Rudney, J.D. Persistence of extracrevicular bacterial reservoirs after treatment of aggressive periodontitis. J. Periodontol. 2008, 79, 2305–2312. [Google Scholar] [CrossRef]
- Christersson, L.A.; Albini, B.; Zambon, J.J.; Wikesjo, U.M.; Genco, R.J. Tissue localization of Actinobacillus actinomycetemcomitans in human periodontitis. I. Light, immunofluorescence and electron microscopic studies. J. Periodontol. 1987, 58, 529–539. [Google Scholar] [CrossRef]
- Paino, A.; Lohermaa, E.; Sormunen, R.; Tuominen, H.; Korhonen, J.; Pöllänen, M.T.; Ihalin, R. Interleukin-1beta is internalised by viable Aggregatibacter actinomycetemcomitans biofilm and locates to the outer edges of nucleoids. Cytokine 2012, 60, 565–574. [Google Scholar] [CrossRef]
- Hayashi, Y.; Matsunaga, T.; Yamamoto, G.; Nishii, K.; Usui, M.; Yamamoto, M.; Tachikawa, T. Comprehensive analysis of gene expression in the junctional epithelium by laser microdissection and microarray analysis. J. Periodontal Res. 2010, 45, 618–625. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Dang, E.; Shi, X.; Jin, L.; Feng, Z.; Hu, L.; Wu, Y.; Wang, G. The pro-inflammatory cytokine IL-22 up-regulates keratin 17 expression in keratinocytes via STAT3 and ERK1/2. PLoS ONE 2012, 7, e40797. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.B.; Alimova, Y.; Myers, T.M.; Ebersole, J.L. Short- and medium-chain fatty acids exhibit antimicrobial activity for oral microorganisms. Arch. Oral Biol. 2011, 56, 650–654. [Google Scholar] [CrossRef] [PubMed]
- Braunthal, S.D.; Holt, S.C.; Tanner, A.C.; Socransky, S.S. Cellular fatty acid composition of Actinobacillus actinomycetemcomitans and Haemophilus aphrophilus. J. Clin. Microbiol. 1980, 11, 625–630. [Google Scholar]
- Pritlove-Carson, S.; Charlesworth, S.; Morgan, P.R.; Palmer, R.M. Cytokeratin phenotypes at the dento-gingival junction in relative health and inflammation, in smokers and nonsmokers. Oral Dis. 1997, 3, 19–24. [Google Scholar] [CrossRef]
- Rompikuntal, P.K.; Thay, B.; Khan, M.K.; Alanko, J.; Penttinen, A.M.; Asikainen, S.; Wai, S.N.; Oscarsson, J. Perinuclear localization of internalized outer membrane vesicles carrying active cytolethal distending toxin from Aggregatibacter actinomycetemcomitans. Infect. Immun. 2012, 80, 31–42. [Google Scholar] [CrossRef]
- Thay, B.; Damm, A.; Kufer, T.A.; Wai, S.N.; Oscarsson, J. Aggregatibacter actinomycetemcomitans outer membrane vesicles are internalized in human host cells and trigger NOD1- and NOD2-dependent NF-kappaB activation. Infect. Immun. 2014, 82, 4034–4046. [Google Scholar] [CrossRef]
- Karched, M.; Paul-Satyaseela, M.; Asikainen, S. A simple viability-maintaining method produces homogenic cell suspensions of autoaggregating wild-type Actinobacillus actinomycetemcomitans. J. Microbiol. Methods 2007, 68, 46–51. [Google Scholar] [CrossRef]
- Oksanen, J.; Hormia, M. An organotypic in vitro model that mimics the dento-epithelial junction. J. Periodontol. 2002, 73, 86–93. [Google Scholar] [CrossRef]
- Mäkelä, M.; Salo, T.; Larjava, H. MMP-9 from TNF alpha-stimulated keratinocytes binds to cell membranes and type I collagen: A cause for extended matrix degradation in inflammation? Biochem. Biophys. Res. Commun. 1998, 253, 325–335. [Google Scholar] [CrossRef]
- Wang, Y.; Shi, W.; Chen, W.; Chen, C. Type IV pilus gene homologs pilABCD are required for natural transformation in Actinobacillus actinomycetemcomitans. Gene 2003, 312, 249–255. [Google Scholar] [CrossRef]
- Paino, A.; Tuominen, H.; Jaaskelainen, M.; Alanko, J.; Nuutila, J.; Asikainen, S.E.; Pelliniemi, L.J.; Pöllönen, M.T.; Chen, C.; Ihalin, R. Trimeric form of intracellular ATP synthase subunit beta of Aggregatibacter actinomycetemcomitans binds human interleukin-1beta. PLoS ONE 2011, 6, e18929. [Google Scholar] [CrossRef]
- Karnovsky, M.J. Use of ferrocyanide-reduced osmium tetroxide in electron microscopy. In Proceedings of the 11th Annual Meeting of American Society for Cell Biology, New Orleans, LA, USA, 17–20 November 1971; p. 146. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Keratin | Dilution | Host | Type | Code | Manufacturer |
|---|---|---|---|---|---|
| Keratin 4 | 1/10 | mouse | monoclonal | MON3015-1 | Sanbio, Holland |
| Keratin 13 | 1/50 | mouse | monoclonal | MUB0340S | Nordic MUbio, Holland |
| Keratin 17 | 1/20 | mouse | monoclonal | M704601-2 | Dako, Denmark |
| Keratin 19 | 1/100 | mouse | monoclonal | M088801-2 | Dako, Denmark |
| Pan-cytokeratin 5,6,8,17,(19) | 1/50 | mouse | monoclonal | M0821 | Dako, Denmark |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beklen, A.; Torittu, A.; Ihalin, R.; Pöllänen, M. Aggregatibacter actinomycetemcomitans Biofilm Reduces Gingival Epithelial Cell Keratin Expression in an Organotypic Gingival Tissue Culture Model. Pathogens 2019, 8, 278. https://doi.org/10.3390/pathogens8040278
Beklen A, Torittu A, Ihalin R, Pöllänen M. Aggregatibacter actinomycetemcomitans Biofilm Reduces Gingival Epithelial Cell Keratin Expression in an Organotypic Gingival Tissue Culture Model. Pathogens. 2019; 8(4):278. https://doi.org/10.3390/pathogens8040278
Chicago/Turabian StyleBeklen, Arzu, Annamari Torittu, Riikka Ihalin, and Marja Pöllänen. 2019. "Aggregatibacter actinomycetemcomitans Biofilm Reduces Gingival Epithelial Cell Keratin Expression in an Organotypic Gingival Tissue Culture Model" Pathogens 8, no. 4: 278. https://doi.org/10.3390/pathogens8040278
APA StyleBeklen, A., Torittu, A., Ihalin, R., & Pöllänen, M. (2019). Aggregatibacter actinomycetemcomitans Biofilm Reduces Gingival Epithelial Cell Keratin Expression in an Organotypic Gingival Tissue Culture Model. Pathogens, 8(4), 278. https://doi.org/10.3390/pathogens8040278