Piper betel Compounds Piperidine, Eugenyl Acetate, and Chlorogenic Acid Are Broad-Spectrum Anti-Vibrio Compounds that Are Also Effective on MDR Strains of the Pathogen



 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Bacterial Culture and Antibacterial Agents
2.2. Evaluation of the Antibacterial Activity of Compounds on Vibrio spp.
2.3. Determination of Compound Minimum Inhibitory Concentrations against Vibrio spp.
2.4. Determination of the Inhibition Parameters of Piper betel Compounds in Combination with Compounds or Standard Drugs on Vibrio spp.
2.5. Determination of the Synergistic Activity of Compounds plus Antibiotics
2.6. Statistical Analysis
3. Results and Discussion
3.1. Bactericidal Activity of Piperidine on Vibrio spp.
3.2. Determination of MICs against Vibrio spp.
3.3. Antibacterial Activity of Mixtures of Piper betel-Derived Compounds against Vibrio spp.
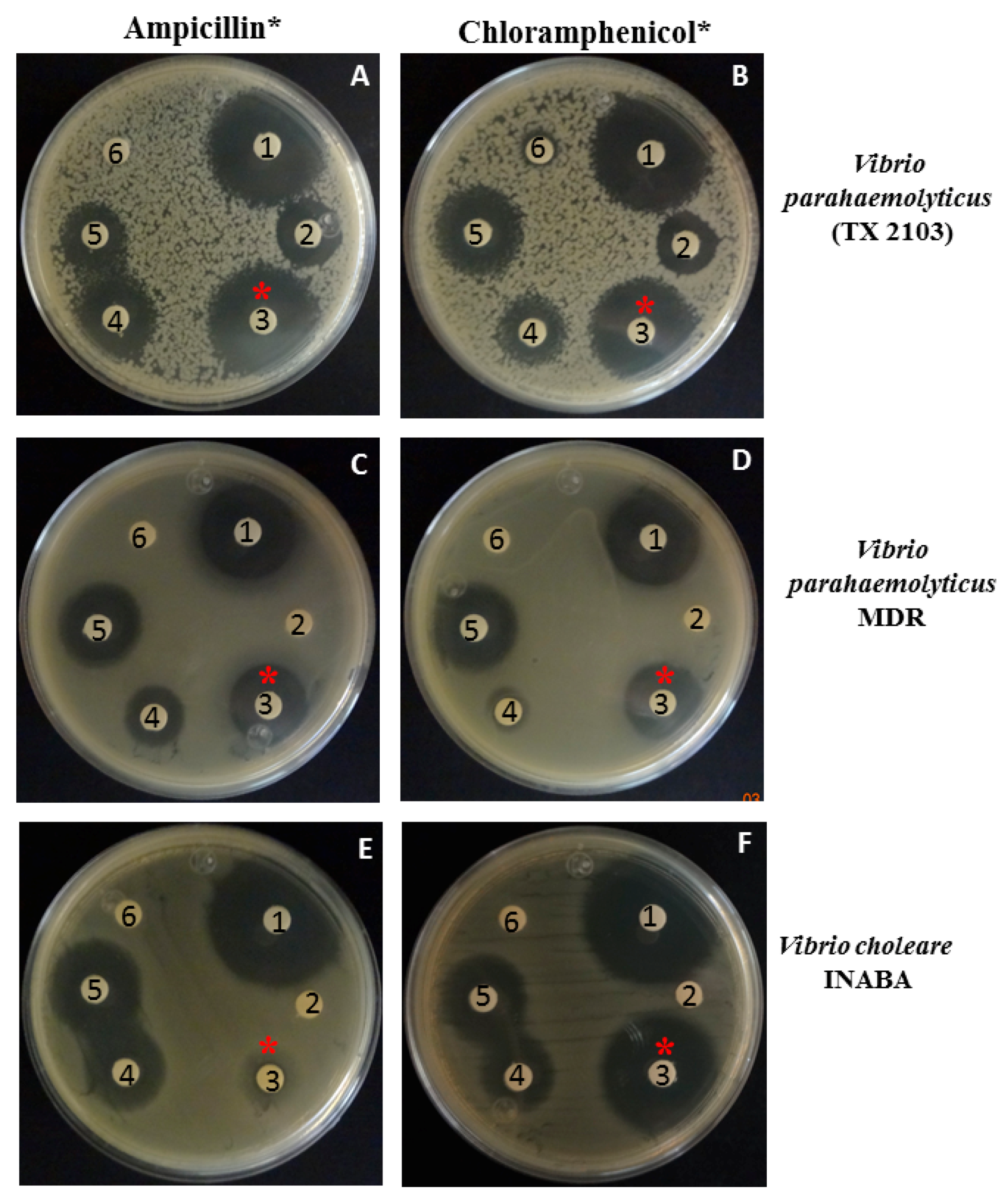
3.4. Antibacterial Activity of Piperidine in Combination with Ampicillin or Chloramphenicol against Vibrio spp.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Scharer, K.; Savioz, S.; Cernela, N.; Saegesser, G.; Stephan, R. Occurrence of vibrio spp. In fish and shellfish collected from the swiss market. J. Food Prot. 2011, 74, 1345–1347. [Google Scholar] [CrossRef]
- Thompson, F.L.; Gevers, D.; Thompson, C.C.; Dawyndt, P.; Naser, S.; Hoste, B.; Munn, C.B.; Swings, J. Phylogeny and molecular identification of vibrios on the basis of multilocus sequence analysis. Appl. Environ. Microbiol. 2005, 71, 5107–5115. [Google Scholar] [CrossRef] [PubMed]
- Janda, J.M.; Powers, C.; Bryant, R.G.; Abbott, S.L. Current perspectives on the epidemiology and pathogenesis of clinically significant vibrio spp. Clin. Microbiol. Rev. 1988, 1, 245–267. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Sakazaki, R. Additional serovars and inter-o antigenic relationships of vibrio cholerae. Jpn. J. Med. Sci. Boil. 1977, 30, 275–277. [Google Scholar] [CrossRef]
- Faruque, S.M.; Albert, M.J.; Mekalanos, J.J. Epidemiology, genetics, and ecology of toxigenic vibrio cholerae. Microbiol. Mol. Boil. Rev. 1998, 62, 1301–1314. [Google Scholar]
- Kontoyiannis, D.P.; Calia, K.E.; Basgoz, N.; Calderwood, S.B. Primary septicemia caused by vibrio cholerae non-o1 acquired on cape cod, massachusetts. Clin. Infect. Dis. 1995, 21, 1330–1333. [Google Scholar] [CrossRef]
- Anderson, A.M.; Varkey, J.B.; Petti, C.A.; Liddle, R.A.; Frothingham, R.; Woods, C.W. Non-o1 vibrio cholerae septicemia: Case report, discussion of literature, and relevance to bioterrorism. Diagn. Microbiol. Infect. Dis. 2004, 49, 295–297. [Google Scholar] [CrossRef]
- Sack, D.A.; Sack, R.B.; Nair, G.B.; Siddique, A.K. Cholera. Lancet 2004, 363, 223–233. [Google Scholar] [CrossRef]
- Pal, B.B.; Khuntia, H.K.; Samal, S.K.; Kar, S.K.; Patnaik, B. Epidemics of severe cholera caused by el tor vibrio cholerae o1 ogawa possessing the ctxb gene of the classical biotype in Orissa, India. Int. J. Infect. Dis. 2010, 14, e384–e389. [Google Scholar] [CrossRef]
- Safa, A.; Sultana, J.; Dac Cam, P.; Mwansa, J.C.; Kong, R.Y. Vibrio cholerae o1 hybrid el tor strains, asia and africa. Emerg. Infect. Dis. 2008, 14, 987–988. [Google Scholar] [CrossRef]
- Fernando, R.R.; Krishnan, S.; Fairweather, M.G.; Ericsson, C.D. Vibrio parahemolyticus septicaemia in a liver transplant patient: A case report. JMCR 2011, 5, 171. [Google Scholar] [CrossRef] [PubMed]
- Baffone, W.; Casaroli, A.; Campana, R.; Citterio, B.; Vittoria, E.; Pierfelici, L.; Donelli, G. ’In vivo’ studies on the pathophysiological mechanism of vibrio parahaemolyticus tdh(+)−induced secretion. Microb. Pathog. 2005, 38, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Ham, H.; Orth, K. The role of type iii secretion system 2 in vibrio parahaemolyticus pathogenicity. J. Microbiol. 2012, 50, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Pezzlo, M.; Valter, P.J.; Burns, M.J. Wound infection associated with vibrio alginolyticus. Am. J. Clin. Pathol. 1979, 71, 476–478. [Google Scholar] [CrossRef] [PubMed]
- Dalsgaard, A.; Glerup, P.; Hoybye, L.L.; Paarup, A.M.; Meza, R.; Bernal, M.; Shimada, T.; Taylor, D.N. Vibrio furnissii isolated from humans in Peru: A possible human pathogen? Epidemiology Infect. 1997, 119, 143–149. [Google Scholar] [CrossRef]
- Thekdi, R.; Lakhani, A.G.; Vachha, S.M.; Chandrakapure, M.R. Vibrio fluvialis (group f vibrio) in Maharashtra. Indian J. Med. Res. 1982, 76, 80–85. [Google Scholar]
- Igbinosa, E.O.; Okoh, A.I. Vibrio fluvialis: An unusual enteric pathogen of increasing public health concern. Int. J. Environ. Res. Public Health 2010, 7, 3628–3643. [Google Scholar] [CrossRef]
- Valentao, P.; Goncalves, R.F.; Belo, C.; de Pinho, P.G.; Andrade, P.B.; Ferreres, F. Improving the knowledge on piper betle: Targeted metabolite analysis and effect on acetylcholinesterase. JSS 2010, 33, 3168–3176. [Google Scholar] [CrossRef]
- Majumdar, B.; Ray Chaudhuri, S.G.; Ray, A.; Bandyopadhyay, S.K. Effect of ethanol extract of piper betle linn leaf on healing of nsaid-induced experimental ulcer--a novel role of free radical scavenging action. Indian. J. Exp. Biol. 2003, 41, 311–315. [Google Scholar]
- Ali, I.; Khan, F.G.; Suri, K.A.; Gupta, B.D.; Satti, N.K.; Dutt, P.; Afrin, F.; Qazi, G.N.; Khan, I.A. In vitro antifungal activity of hydroxychavicol isolated from piper betle l. Ann. Clin. Microbiol. Antimicrob. 2010, 9, 7. [Google Scholar] [CrossRef]
- Trakranrungsie, N.; Chatchawanchonteera, A.; Khunkitti, W. Ethnoveterinary study for antidermatophytic activity of piper betle, alpinia galanga and allium ascalonicum extracts in vitro. Res. Vet. Sci. 2008, 84, 80–84. [Google Scholar] [CrossRef]
- Ganguly, S.; Mula, S.; Chattopadhyay, S.; Chatterjee, M. An ethanol extract of piper betle linn. Mediates its anti-inflammatory activity via down-regulation of nitric oxide. J. Pharm. Pharmacol. 2007, 59, 711–718. [Google Scholar] [CrossRef]
- Barh, D.; Gupta, K.; Jain, N.; Khatri, G.; Leon-Sicairos, N.; Canizalez-Roman, A.; Tiwari, S.; Verma, A.; Rahangdale, S.; Shah Hassan, S.; et al. Conserved host-pathogen ppis. Globally conserved inter-species bacterial ppis based conserved host-pathogen interactome derived novel target in c. Pseudotuberculosis, c. Diphtheriae, m. Tuberculosis, c. Ulcerans, y. Pestis, and e. Coli targeted by piper betel compounds. Integr. Biol. (Camb). 2013, 5, 495–509. [Google Scholar]
- Tiwari, S.; Barh, D.; Imchen, M.; Rao, E.; Kumavath, R.K.; Seenivasan, S.P.; Jaiswal, A.K.; Jamal, S.B.; Kumar, V.; Ghosh, P.; et al. Acetate kinase (ack) is essential for microbial growth and betel-derived compounds potentially target ack, phop and mdr proteins in m. Tuberculosis, v. Cholerae and pathogenic e. Coli: An in silico and in vitro study. Curr. Top. Med. Chem. 2018, 18, 2731–2740. [Google Scholar] [CrossRef]
- DePaola, A.; Kaysner, C.A.; Bowers, J.; Cook, D.W. Environmental investigations of vibrio parahaemolyticus in oysters after outbreaks in Washington, Texas, and New York (1997 and 1998). Appl. Environ. Microb. 2000, 66, 4649–4654. [Google Scholar] [CrossRef]
- Myers, M.L.; Panicker, G.; Bej, A.K. Pcr detection of a newly emerged pandemic vibrio parahaemolyticus o3:K6 pathogen in pure cultures and seeded waters from the gulf of Mexico. Appl. Environ. Microb. 2003, 69, 2194–2200. [Google Scholar] [CrossRef]
- Velazquez-Roman, J.; Leon-Sicairos, N.; de Jesus Hernandez-Diaz, L.; Canizalez-Roman, A. Pandemic vibrio parahaemolyticus o3:K6 on the american continent. Front. Microbiol. 2014, 3, 110. [Google Scholar] [CrossRef]
- Velazquez-Roman, J.; Leon-Sicairos, N.; Flores-Villasenor, H.; Villafana-Rauda, S.; Canizalez-Roman, A. Association of pandemic vibrio parahaemolyticus o3:K6 present in the coastal environment of northwest Mexico with cases of recurrent diarrhea between 2004 and 2010. Appl. Environ. Microb. 2012, 78, 1794–1803. [Google Scholar] [CrossRef]
- De Jesus Hernandez-Diaz, L.; Leon-Sicairos, N.; Velazquez-Roman, J.; Flores-Villasenor, H.; Guadron-Llanos, A.M.; Martinez-Garcia, J.J.; Vidal, J.E.; Canizalez-Roman, A. A pandemic vibrio parahaemolyticus o3:K6 clone causing most associated diarrhea cases in the pacific northwest coast of mexico. Front. Microbiol. 2015, 6, 221. [Google Scholar] [CrossRef]
- Zavala-Norzagaray, A.A.; Aguirre, A.A.; Velazquez-Roman, J.; Flores-Villasenor, H.; Leon-Sicairos, N.; Ley-Quinonez, C.P.; Hernandez-Diaz Lde, J.; Canizalez-Roman, A. Isolation, characterization, and antibiotic resistance of vibrio spp. In sea turtles from northwestern Mexico. Front. Microbiol. 2015, 6, 635. [Google Scholar] [CrossRef]
- Balaji, K.; Okonjo, P.A.; Thenmozhi, R.; Karutha Pandian, S. Virulence and multidrug resistance patterns of vibrio cholerae o1 isolates from diarrheal outbreaks of south india during 2006-2009. Microb. Drug Resist. 2013, 19, 198–203. [Google Scholar] [CrossRef]
- Tran, H.D.; Alam, M.; Trung, N.V.; Kinh, N.V.; Nguyen, H.H.; Pham, V.C.; Ansaruzzaman, M.; Rashed, S.M.; Bhuiyan, N.A.; Dao, T.T.; et al. Multi-drug resistant vibrio cholerae o1 variant el tor isolated in northern vietnam between 2007 and 2010. J. Med. Microbiol. 2012, 61, 431–437. [Google Scholar] [CrossRef]
- Krishna, B.V.; Patil, A.B.; Chandrasekhar, M.R. Fluoroquinolone-resistant vibrio cholerae isolated during a cholera outbreak in India. Trans. R. Soc. Trop. Med. Hyg. 2006, 100, 224–226. [Google Scholar] [CrossRef]
- Sudha, S.; Divya, P.S.; Francis, B.; Hatha, A.A. Prevalence and distribution of vibrio parahaemolyticus in finfish from cochin (south india). Vet. Ital. 2012, 48, 269–281. [Google Scholar] [PubMed]
- Oh, E.G.; Son, K.T.; Yu, H.; Lee, T.S.; Lee, H.J.; Shin, S.; Kwon, J.Y.; Park, K.; Kim, J. Antimicrobial resistance of vibrio parahaemolyticus and vibrio alginolyticus strains isolated from farmed fish in korea from 2005 through 2007. J. Food Prot. 2011, 74, 380–386. [Google Scholar] [CrossRef]
- Rajpara, N.; Patel, A.; Tiwari, N.; Bahuguna, J.; Antony, A.; Choudhury, I.; Ghosh, A.; Jain, R.; Bhardwaj, A.K. Mechanism of drug resistance in a clinical isolate of vibrio fluvialis: Involvement of multiple plasmids and integrons. Int. J. Antimicrob. Agents 2009, 34, 220–225. [Google Scholar] [CrossRef]
- Mohanty, P.; Patel, A.; Kushwaha Bhardwaj, A. Role of h- and d- mate-type transporters from multidrug resistant clinical isolates of vibrio fluvialis in conferring fluoroquinolone resistance. PLoS ONE 2012, 7, e35752. [Google Scholar] [CrossRef]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Breed, R.S.; Dotterrer, W.D. The number of colonies allowable on satisfactory agar plates. J. Bacteriol. 1916, 1, 321–331. [Google Scholar]
- Moody, J.A.; Moody, J. Synergism Testing: Broth Microdilution Checkerboard and Broth Macrodilution Methods, 2nd ed.; ASM Press: Washington, DC, USA, 2007; Volume 1. [Google Scholar]
- Odds, F.C. Synergy, antagonism, and what the chequerboard puts between them. J. Antimicrob. Chemother. 2003, 52, 1. [Google Scholar] [CrossRef]
- Rathee, J.S.; Patro, B.S.; Mula, S.; Gamre, S.; Chattopadhyay, S. Antioxidant activity of piper betel leaf extract and its constituents. J. Agric. Food Chem. 2006, 54, 9046–9054. [Google Scholar] [CrossRef]
- Nair, R.; Chanda, S. Antimicrobial activity of terminalia catappa, manilkara zapota and piper betel leaf extract. Indian. J. Pharm. Sci. 2008, 70, 390–393. [Google Scholar] [PubMed]
- Wiart, C.; Hannah, N.A.; Yassim, M.; Hamimah, H.; Sulaiman, M. Antimicrobial activity of tiger’s betel (piper porphyrophyllum n.E. Br., piperaceae). Phytother. Res. 2004, 18, 783–784. [Google Scholar] [CrossRef] [PubMed]
- Worthington, R.J.; Melander, C. Combination approaches to combat multidrug-resistant bacteria. Trend. Biotechnol. 2013, 31, 177–184. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Vibrio Strains | Resistance Pattern | |||||||
|---|---|---|---|---|---|---|---|---|
| Tetracycline | Chloramphenicol | Ampicillin | SXT | Cefotaxime | Gentamicin | Ciprofloxacin | Nalidixic Acid | |
| V. cholerae O1 Inaba | S | S | S | S | S | R | R | R |
| V. cholerae O1 Ogawa | R | S | S | S | R | S | S | R |
| Vibrio fuvialis | I | S | R | I | R | S | S | I |
| Vibrio furnissii | S | S | S | S | S | S | S | S |
| V. parahaemolyticus MDR | R | S | S | R | R | S | R | S |
| V. parahaemolyticus TX2103 | S | S | R | S | S | S | S | S |
| V. vulnificus | R | S | S | S | I | I | R | S |
| V. alginolyticus | R | S | R | S | R | S | S | R |
| V. cholerae non-O1 serotype and toxigenic | S | S | S | S | S | S | R | S |
| Vibrio Species | MDR Drugs | References |
|---|---|---|
| Vibrio cholerae O1 (Inaba and Ogawa serotype) | Ampicillin, polymyxin B, nalidixic acid, co-trimoxazole, norfloxacin, ciprofloxacin, doxycycline, gentamicin, chloramphenicol | Balaji et al. 2013 |
| V. cholerae serogroup O1 Ogawa and El Tor | Co-trimoxazole, nalidixic acid, tetracycline, azithromycin, fluoroquinolones | Tran et al. 2012 |
| V. cholerae non-O1, non-O139 serogroups | Norfloxacin and ciprofloxacin | Krishna et al. 2006 |
| V. parahaemolyticus | Ampicillin and streptomycin, followed by carbenicillin, cefpodoxime, cephalothin, colistin, amoxycillin, nalidixic acid, tetracycline, chloramphenicol, and ciprofloxacin | Sudha et al. 2012 |
| V. alginolyticus | Ampicillin, tetracycline, trimethoprim, and rifampin | Oh et al. 2011 |
| Vibrio fluvialis | 14 antibiotics including neomycin, co-trimoxazole, nalidixic acid, trimethoprim, ampicillin, kanamycin, ciprofloxacin, streptomycin, sulfisoxazole, chloramphenicol, norfloxacin | Rajpara et al. 2009; Mohanty et al. 2012 |
| Compounds | MIC (mg/mL) a | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| MICS of Each Compound Incubated in the Cultures | MICS of Each Compound When All Were Incubated in the Cultures | |||||||||
| Vibrio cholerae INABA | Vibrio parahaemolyticus TX 2103 | Vibrio parahaemolyticus O3:K6 | Vibrio furnisii | Vibrio fluvialis | Vibrio cholerae INABA | Vibrio parahaemolyticus TX 2103 | Vibrio parahaemolyticus O3:K6 | Vibrio furnisii | Vibrio fluvialis | |
| Piperidine mg/mL | 2 ± 0.5 | 2 ± 0.5 | 6.5 ± 0.5 | 4 ± 0.5 | 2 ± 0.8 | 0.6 ± 0.4 | 0.6 ± 0.3 | 1.7 ± 0.5 | 1 ± 0.6 | 1 ± 0.4 |
| Chlorogenic acid mg/mL | 5.5 ± 0.5 | 5.5 ± 1 | 16 ± 4 | 2 ± 0.5 | 6.5 ± 0.5 | 1.8 ± 0.2 | 1.8 ± 0.2 | 2 ± 0.8 | 1 ± 0.4 | 2 ± 0.4 |
| Eugenyl acetate mg/mL | 20 ± 4 | 5.5 ± 0.5 | ≥16 ± 6 | 6.5 ± 0.5 | 6.5 ± 1 | 2 ± 0.25 | 0.5 ± 0.25 | 2 ± 0.8 | 2 ± 0.6 | 2 ± 0.8 |
| Pinoresinol mg/mL | ≥30 | ≥30 | ≥30 | - | - | - | - | - | - | - |
| Strains | Agent | MIC | FIC Index * | Outcome * | |
|---|---|---|---|---|---|
| Alone | Combination | ||||
| Vibrio parahaemolyticus MDR | Chloramphenicol (µg/mL) | 30 | 22.5 | 0.75 | Partial synergy |
| Piperidine (mg/mL) | 4 | 3 | |||
| Ampicillin (µg/mL) | 50 | 10 | 0.45 | Synergy | |
| Piperidine (mg/mL) | 4 | 1 | |||
| Chloramphenicol (µg/mL) | 30 | 15 | 1 | Partial synergy | |
| Chlorogenic acid (mg/mL) | 20 | 10 | |||
| Ampicillin (µg/mL) | 50 | 10 | 0.35 | Synergy | |
| Chlorogenic acid (mg/mL) | 20 | 3 | |||
| Chloramphenicol (µg/mL) | 30 | 10 | 0.83 | Partial synergy | |
| Eugenyl acetate (mg/mL) | 20 | 10 | |||
| Ampicillin (µg/mL) | 50 | 10 | 0.7 | Partial synergy | |
| Eugenyl acetate (mg/mL) | 20 | 10 | |||
| Vibrio parahaemolyticus TX2103 | Chloramphenicol (µg/mL) | 30 | 11.25 | 0.75 | Partial synergy |
| Piperidine (mg/mL) | 4 | 1.5 | |||
| Ampicillin (µg/mL) | ≥100 | 10 | 0.83 | Partial synergy | |
| Piperidine (mg/mL) | 4 | 2 | |||
| Chloramphenicol (µg) | 30 | 15 | 1 | Partial synergy | |
| Chlorogenic acid (mg/mL) | 6 | 3 | |||
| Ampicillin (µg/mL) | ≥100 | 10 | 0.6 | Partial synergy | |
| Chlorogenic acid (mg/mL) | 6 | 3 | |||
| Chloramphenicol (µg/mL) | 30 | 15 | 1 | Partial synergy | |
| Eugenyl acetate (mg/mL) | 6 | 3 | |||
| Ampicillin (µg/mL) | ≥100 | 10 | 0.6 | Partial synergy | |
| Eugenyl acetate (mg/mL) | 6 | 3 | |||
| Vibrio cholerae O1 INABA | Chloramphenicol (µg/mL) | 30 | 7.5 | 0.5 | Synergy |
| Piperidine (mg/mL) | 4 | 1 | |||
| Ampicillin (µg/mL) | 50 | 10 | 0.45 | Synergy | |
| Piperidine (mg/mL) | 4 | 1 | |||
| Chloramphenicol (µg/mL) | 30 | 10 | 0.83 | Partial synergy | |
| Chlorogenic acid (mg/mL) | 6 | 3 | |||
| Ampicillin (µg/mL) | 50 | 10 | 0.7 | Partial synergy | |
| Chlorogenic acid (mg/mL) | 6 | 3 | |||
| Chloramphenicol (µg/mL) | 30 | 10 | 0.58 | Partial synergy | |
| Eugenyl acetate (mg/mL) | 20 | 5 | |||
| Ampicillin (µg/mL) | 50 | 10 | 0.45 | Synergy | |
| Eugenyl acetate (mg/mL) | 20 | 5 | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acosta-Smith, E.; Leon-Sicairos, N.; Tiwari, S.; Flores-Villaseñor, H.; Canizalez-Roman, A.; Kumavath, R.; Ghosh, P.; Azevedo, V.; Barh, D. Piper betel Compounds Piperidine, Eugenyl Acetate, and Chlorogenic Acid Are Broad-Spectrum Anti-Vibrio Compounds that Are Also Effective on MDR Strains of the Pathogen. Pathogens 2019, 8, 64. https://doi.org/10.3390/pathogens8020064
Acosta-Smith E, Leon-Sicairos N, Tiwari S, Flores-Villaseñor H, Canizalez-Roman A, Kumavath R, Ghosh P, Azevedo V, Barh D. Piper betel Compounds Piperidine, Eugenyl Acetate, and Chlorogenic Acid Are Broad-Spectrum Anti-Vibrio Compounds that Are Also Effective on MDR Strains of the Pathogen. Pathogens. 2019; 8(2):64. https://doi.org/10.3390/pathogens8020064
Chicago/Turabian StyleAcosta-Smith, Erika, Nidia Leon-Sicairos, Sandeep Tiwari, Hector Flores-Villaseñor, Adrian Canizalez-Roman, Ranjith Kumavath, Preetam Ghosh, Vasco Azevedo, and Debmalya Barh. 2019. "Piper betel Compounds Piperidine, Eugenyl Acetate, and Chlorogenic Acid Are Broad-Spectrum Anti-Vibrio Compounds that Are Also Effective on MDR Strains of the Pathogen" Pathogens 8, no. 2: 64. https://doi.org/10.3390/pathogens8020064
APA StyleAcosta-Smith, E., Leon-Sicairos, N., Tiwari, S., Flores-Villaseñor, H., Canizalez-Roman, A., Kumavath, R., Ghosh, P., Azevedo, V., & Barh, D. (2019). Piper betel Compounds Piperidine, Eugenyl Acetate, and Chlorogenic Acid Are Broad-Spectrum Anti-Vibrio Compounds that Are Also Effective on MDR Strains of the Pathogen. Pathogens, 8(2), 64. https://doi.org/10.3390/pathogens8020064