Preliminary Study on the Use of Chitosan as an Eco-Friendly Alternative to Control Fusarium Growth and Mycotoxin Production on Maize and Wheat



Abstract
1. Introduction
2. Results
2.1. Chitosan Characterization
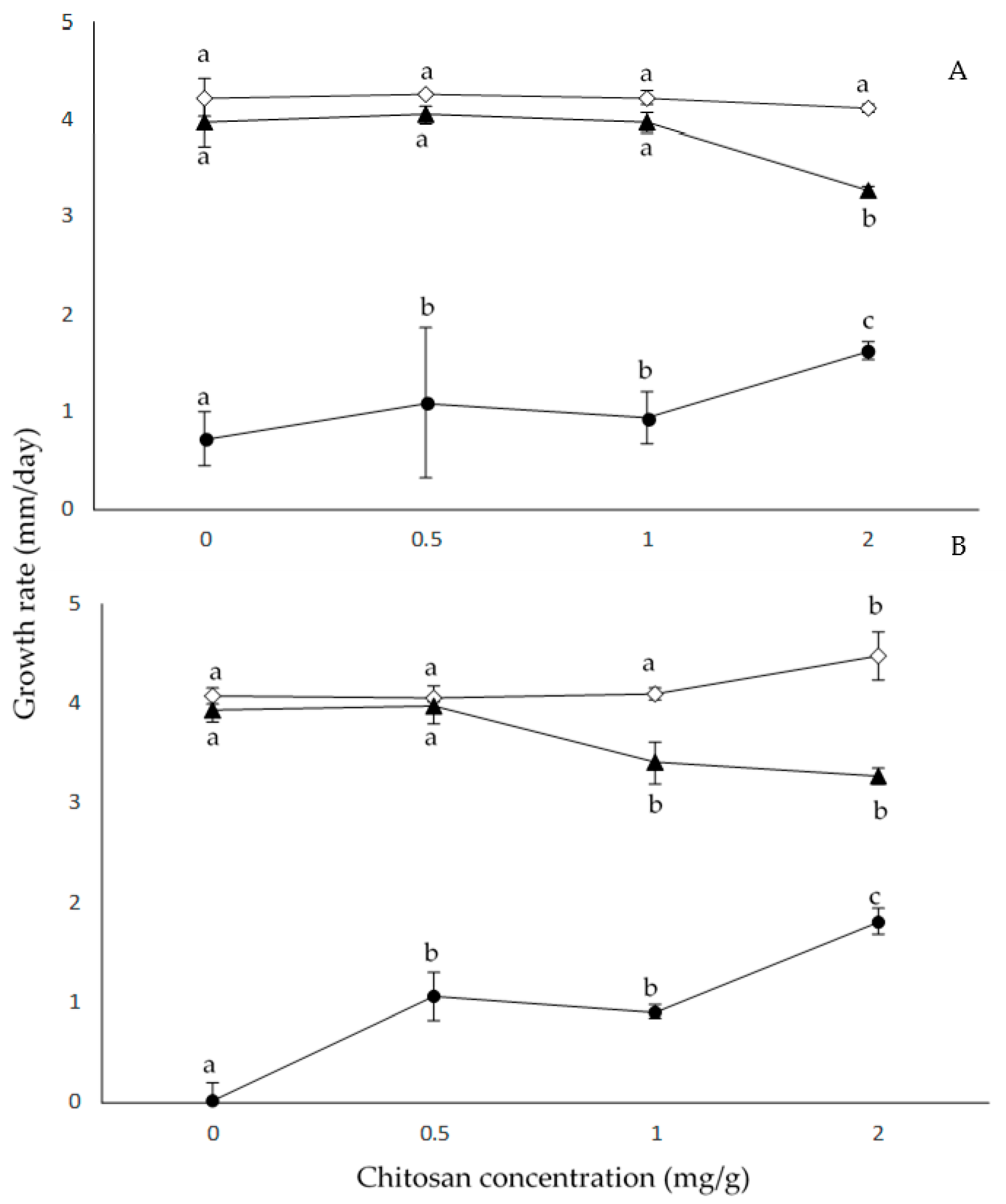
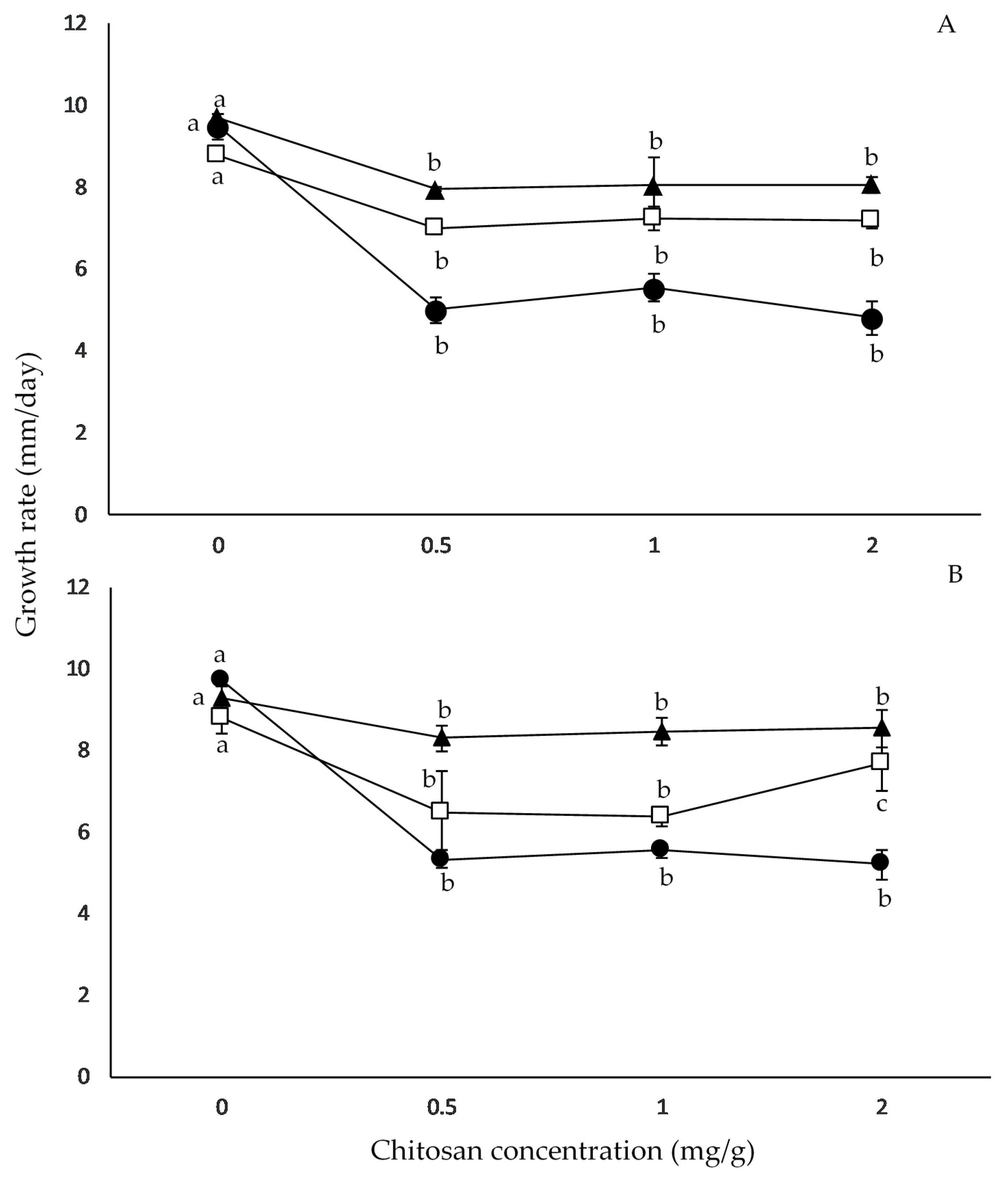
2.2. Effect of Chitosan Concentration and aW on Growth Rates
2.3. Effect of Chitosan Concentration and aW on Mycotoxin Production
3. Discussion
4. Materials and Methods
4.1. Chitosan Solution
4.2. Chitosan Characterization
4.3. Fungal Strains
4.4. Grains
4.5. Inoculation, Incubation, and Growth Assessment
4.6. Mycotoxin Determination
4.6.1. Fumonisin Determination in Maize
4.6.2. Deoxynivalenol Determination in Wheat
4.7. Statistical Treatment of Results
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Marin, S.; Sanchis, V.; Ramos, A.J.; Vinas, I.; Magan, N. Environmental factors, in vitro interactions, and niche overlap between Fusarium moniliforme, F. proliferatum, and F. graminearum, Aspergillus and Penicillium species from maize grain. Mycol. Res. 1998, 102, 831–837. [Google Scholar] [CrossRef]
- Kriek, N.P.J.; Marases, W.F.O.; Thiel, P.G. Hepato-and Cardiotoxicity of Fusarium verticillioides (F. moniliforme) isolates from southern african maize. Food Cosmet. Toxicol. 1981, 19, 447–456. [Google Scholar] [CrossRef]
- Cardwell, K.F.; Kling, J.G.; Maziya-Dixon, B.; Bosque-Pérez, N.A. Interactions between Fusarium verticillioides, Aspergillus flavus, and insect infestation in four maize genotypes in Lowland Africa. Phytopathology 2000, 90, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Marasas, W.F.O.; Kriek, N.P.J.; Wiggins, V.M.; Steyn, P.S.; Towers, D.K.; Hastie, T.J. Incidence, geographic distribution, and toxigenicity of Fusarium species in South African corn. Phytopathology 1979, 69, 1181–1185. [Google Scholar] [CrossRef]
- González, H.H.L.; Resnik, S.L.; Boca, R.T.; Marasas, W.F.O. Mycoflora of Argentinian corn harvested in the main production area in 1990. Mycopathologia 1995, 130, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Chulze, S.N.; Ramirez, M.L.; Farnochi, M.C.; Pascale, M.; Visconti, A.; March, G. Fusarium and fumonisin occurrence in Argentinian corn at different ear maturity stages. J. Agric. Food Chem. 1996, 44, 2797–2801. [Google Scholar] [CrossRef]
- Picco, M.; Nesci, A.; Barros, G.; Cavaglieri, L.; Etcheverry, M. Aflatoxin B1 and fumosin B1 in mixed cultures of Aspergillus flavus and Fusarium proliferatum on maize. Nat. Toxins 1999, 7, 331–336. [Google Scholar] [CrossRef]
- Munkvold, G.P. Fusarium species and their associated mycotoxins. In Mycotoxigenic Fungi: Methods and Protocols, Methods in Molecular Biology; Moretti, A., Susca, A., Eds.; Humana Press: New York, NY, USA, 2017; Volume 1542, pp. 51–106. ISBN 978-1-4939-6707-0. [Google Scholar]
- Marasas, W.F.O. Discovery and occurrence of the fumonisins: A historical perspective. Environ. Health Perspect. 2001, 109, 5. [Google Scholar]
- Marasas, W.F.O.; Riley, R.T.; Hendricks, K.A.; Stevens, V.L.; Sadler, T.W.; Gelineau-van Waes, J.; Missmer, S.A.; Cabrera, J.; Torres, O.; Gelderblom, W.C.A.; et al. Fumonisins disrupt sphingolipid metabolism, folate transport, and neural tube development in embryo culture and in vivo: A potential risk factor for human neural tube defects among populations consuming fumonisin-contaminated maize. J. Nutr. 2004, 134, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Missmer, S.A.; Suarez, L.; Felkner, M.; Wang, E.; Merrill, A.H.; Rothman, K.J.; Hendricks, K.A. Exposure to fumonisins and the occurrence of neural tube defects along the Texas–Mexico border. Environ. Health Perspect. 2006, 114, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Wang, S.; Hu, X.; Su, J.; Huang, T.; Yu, J.; Tang, L.; Gao, W.; Wang, J.-S. Fumonisin B1 contamination of home-grown corn in high-risk areas for esophageal and liver cancer in China. Food Addit. Contam. 2007, 24, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Wild, C.P.; Gong, Y.Y. Mycotoxins and human disease: A largely ignored global health issue. Carcinogenesis 2010, 31, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Kimanya, M.E.; De Meulenaer, B.; Roberfroid, D.; Lachat, C.; Kolsteren, P. Fumonisin exposure through maize in complementary foods is inversely associated with linear growth of infants in Tanzania. Mol. Nutr. Food Res. 2010, 54, 1659–1667. [Google Scholar] [CrossRef] [PubMed]
- Shirima, C.P.; Kimanya, M.E.; Routledge, M.N.; Srey, C.; Kinabo, J.L.; Humpf, H.-U.; Wild, C.P.; Gong, Y.Y. A prospective study of growth and biomarkers of exposure to aflatoxin and fumonisin during Early Childhood in Tanzania. Environ. Health Perspect. 2014. [Google Scholar] [CrossRef] [PubMed]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans; World Health Organization and International Agency for Research on Cancer. Some Traditional Herbal Medicines, Some Mycotoxins, Naphthalene and Styrene; IARC (International Agency for Research on Cancer): Geneva, Switzerland, 1980; Volume 82, pp. 301–366. [Google Scholar]
- World Health Organization (WHO). Safety Evaluation of Certain Mycotoxins in Food; Food & Agriculture Org.: Rome, Italy, 2001; ISBN 978-92-5-104664-7. [Google Scholar]
- Bottalico, A.; Perrone, G. Toxigenic Fusarium species and mycotoxins associated with head blight in small-grain cereals in Europe. In Mycotoxins in Plant Disease; Logrieco, A., Bailey, J.A., Corazza, L., Cooke, B.M., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 611–624. ISBN 978-94-010-3939-0. [Google Scholar]
- Lori, G.A.; Sisterna, M.N.; Haidukowski, M.; Rizzo, I. Fusarium graminearum and deoxynivalenol contamination in the durum wheat area of Argentina. Microbiol. Res. 2003, 158, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, M.L.; Chulze, S.; Magan, N. Temperature and water activity effects on growth and temporal deoxynivalenol production by two Argentinean strains of Fusarium graminearum on irradiated wheat grain. Int. J. Food Microbiol. 2006, 106, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, M.L.; Reynoso, M.M.; Farnochi, M.C.; Torres, A.M.; Leslie, J.F.; Chulze, S.N. Population genetic structure of Gibberella zeae isolated from wheat in Argentina. Food Addit. Contam. 2007, 24, 1115–1120. [Google Scholar] [CrossRef] [PubMed]
- Dalcero, A.; Torres, A.; Etcheverry, M.; Chulze, S.; Varsavsky, E. Occurrence of deoxynivalenol and fusarium graminearum in Argentinian wheat. Food Addit. Contam. 1997, 14, 11–14. [Google Scholar] [CrossRef] [PubMed]
- González, H.H.L.; Pacin, A.; Resnik, S.L.; Martinez, E.J. Deoxynivalenol and contaminant mycoflora in freshly harvested Argentinian wheat in 1993. Mycopathologia 1996, 135, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Palazzini, J.M.; Alberione, E.; Torres, A.; Donat, C.; Köhl, J.; Chulze, S. Biological control of Fusarium graminearum sensu stricto, causal agent of Fusarium head blight of wheat, using formulated antagonists under field conditions in Argentina. Biol. Control 2016, 94, 56–61. [Google Scholar] [CrossRef]
- Audenaert, K.; Vanheule, A.; Höfte, M.; Haesaert, G.; Audenaert, K.; Vanheule, A.; Höfte, M.; Haesaert, G. Deoxynivalenol: A major player in the multifaceted response of Fusarium to its environment. Toxins 2013, 6, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J.J. Deoxynivalenol: Mechanisms of action, human exposure, and toxicological relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Sevastos, A.; Markoglou, A.; Labrou, N.E.; Flouri, F.; Malandrakis, A. Molecular characterization, fitness and mycotoxin production of Fusarium graminearum laboratory strains resistant to benzimidazoles. Pest. Biochem. Physiol. 2016, 128, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Zhang, L.; Yang, L.; Zhang, F.; Norse, D.; Zhu, Z. Agricultural non-point source pollution in China: Causes and mitigation measures. Ambio 2012, 41, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Mateo, E.M.; Gómez, J.V.; Gimeno-Adelantado, J.V.; Romera, D.; Mateo-Castro, R.; Jiménez, M. Assessment of azole fungicides as a tool to control growth of Aspergillus flavus and aflatoxin B1 and B2 production in maize. Food Addit. Contam. Part A 2017, 34, 1039–1051. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, M.L.; Chulze, S.; Magan, N. Impact of environmental factors and fungicides on growth and deoxinivalenol production by Fusarium graminearum isolates from Argentinian wheat. Crop Protec. 2004, 23, 117–125. [Google Scholar] [CrossRef]
- Malerca, M.; Cerana, R. Chitosan effects on plant systems. Int. J. Mol. Sci. 2016, 17, 996. [Google Scholar] [CrossRef] [PubMed]
- Alburquenque, C.; Bucarey, S.A.; Neira-Carrillo, A.; Urza, B.; Hermosilla, G.; Tapia, C.V. Antifungal activity of low molecular weight chitosan against clinical isolates of Candida spp. Med. Mycol. 2010, 48, 1018–1023. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, R.; Liu, W. Effects of chitin and its derivative chitosan on postharvest decay of fruits: A review. Int. J. Mol. Sci. 2011, 12, 917–934. [Google Scholar] [CrossRef] [PubMed]
- Jayakumar, R.; Prabaharan, M.; Sudheesh Kumar, P.T.; Nair, S.V.; Tamura, H. Biomaterials based on chitin and chitosan in wound dressing applications. Biotechnol. Adv. 2011, 29, 322–337. [Google Scholar] [CrossRef] [PubMed]
- Kong, M.; Chen, X.G.; Xing, K.; Park, H.J. Antimicrobial properties of chitosan and mode of action: A state of the art review. Int. J. Food Microbiol. 2010, 144, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Ziani, K.; Fernández-Pan, I.; Royo, M.; Maté, J.I. Antifungal activity of films and solutions based on chitosan against typical seed fungi. Food Hydrocoll. 2009, 23, 2309–2314. [Google Scholar] [CrossRef]
- Cota-Arriola, O.; Cortez-Rocha, M.O.; Rosas-Burgos, E.C.; Burgos-Hernández, A.; López-Franco, Y.L.; Plascencia-Jatomea, M. Antifungal effect of chitosan on the growth of Aspergillus parasiticus and production of aflatoxin B1. Polym. Int. 2011, 60, 937–944. [Google Scholar] [CrossRef]
- Cuero, R.G.; Osuji, G.; Washington, A. N-carboxymethyl chitosan inhibition of aflatoxin production: Role of zinc. Biotechnol. Lett. 1991, 13, 441–444. [Google Scholar] [CrossRef]
- Fonseca Moreira da Silva, J.; Prado, G.; Gazzinelli Cruz Madeira, J.E.; Silva Oliveira, M.; Gomes Faraco, A.A.; Martins Malta, C.; Nicoli, J.R.; Sanzio Pimenta, R. Utilização de filme de quitosana para o controle de aflatoxinas em amendoim. Bragantia 2015, 74, 467–475. [Google Scholar] [CrossRef]
- Reddy, M.V.B.; Arul, J.; Ait-Barka, E.; Angers, P.; Richard, C.; Castaigne, F. EVect of Chitosan on Growth and Toxin Production by Alternaria alternata f. sp. lycopersici. Biocontrol Sci. Technol. 1998, 8, 33–43. [Google Scholar] [CrossRef]
- Mateo, E.M.; Valle-algarra, F.M.; Mateo, R.; Jiménez, M.; Magan, N. Effect of fenpropimorph, prochloraz and tebuconazole on growth and production of T-2 and HT-2 toxins by Fusarium langsethiae in oat-based medium. Int. J. Food Microbiol. 2011, 151, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Ferrochio, L.V.; Cendoya, E.; Zachetti, V.G.L.; Farnochi, M.C.; Massad, W.; Ramirez, M.L. Combined effect of chitosan and water activity on growth and fumonisin production by Fusarium verticillioides and Fusarium proliferatum on maize-based media. Int. J. Food Microbiol. 2014, 185, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.C.; Wang, H.L.; Chen, Y.M.; Li, S.L. Effect of abiotic factors on the antibacterial activity of chitosan against waterborne pathogens. Bioresour. Technol. 2003, 88, 179–184. [Google Scholar] [CrossRef]
- Marín, S.; Sanchis, V.; Teixido, A.; Saenz, R.; Ramos, A.J.; Vinas, I.; Magan, N. Water and temperature relations and microconidial germination of Fusarium moniliforme and Fusarium proliferatum from maize. Can. J. Microbiol. 1996, 42, 1045–1050. [Google Scholar] [CrossRef] [PubMed]
- Verlee, A.; Mincke, S.; Stevens, C.V. Recent developments in antibacterial and antifungal chitosan and its derivatives. Carbohydr. Polym. 2017, 164, 268–283. [Google Scholar] [CrossRef] [PubMed]
- Hosseinnejad, M.; Jafari, S.M. Evaluation of different factors affecting antimicrobial properties of chitosan. Int. J. Biol. Macromol. 2016, 85, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Sahariah, P.; Másson, M. Antimicrobial Chitosan and Chitosan Derivatives: A Review of the Structure–Activity Relationship. Biomacromolecules 2017, 18, 3846–3868. [Google Scholar] [CrossRef] [PubMed]
- Sanchis, V.; Magan, N. Environmental conditions affecting mycotoxins. In Mycotoxins in Food: Detection and Control; Magan, N., Olsen, M., Eds.; Woodhead Publishing Ltd.: Oxford, UK, 2004; pp. 174–189. ISBN 978-1-85573-733-4. [Google Scholar]
- Khan, M.R.; Doohan, F.M. Comparison of the efficacy of chitosan with that of a fluorescent pseudomonad for the control of Fusarium head blight disease of cereals and associated mycotoxin contamination of grain. Biol. Control 2009, 48, 48–54. [Google Scholar] [CrossRef]
- Kheiri, A.; Moosawi Jorf, S.A.; Mallihipour, A.; Saremi, H.; Nikkhah, M. Application of chitosan and chitosan nanoparticles for the control of Fusarium head blight of wheat (Fusarium graminearum) in vitro and greenhouse. Int. J. Biol. Macromol. 2016, 93, 1261–1272. [Google Scholar] [CrossRef] [PubMed]
- Chulze, S.N. Strategies to reduce mycotoxin levels in maize during storage: A review. Food Addit. Contam. Part A 2010, 27, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Pacin, A.M.; Ciancio Bovier, E.; González, H.H.L.; Whitechurch, E.M.; Martínez, E.J.; Resnik, S.L. Fungal and fumonisins contamination in Argentine maize (Zea mays L.) silo bags. J. Agric. Food Chem. 2009, 57, 2778–2781. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.-S.; Yang, P.; Wu, A.-B.; Zuo, D.-Y.; He, W.-J.; Guo, M.-W.; Huang, T.; Li, H.-P.; Liao, Y.-C. Variation in the microbiome, trichothecenes, and aflatoxins in stored wheat grains in Wuhan, China. Toxins 2018, 10, 171. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Du, Y.; Wang, X.; Sun, L. Chitosan kills bacteria through cell membrane damage. Int. J. Food Microbiol. 2004, 95, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Knaul, J.Z.; Kasaii, M.R.; Bui, V.T.; Creber, K.A. Characterization of deacetylated chitosan and chitosan molecular weight review. Can. J. Chem. 1998, 76, 1699–1706. [Google Scholar] [CrossRef]
- Etcheverry, M.; Torres, A.; Ramirez, M.L.; Chulze, S.; Magan, N. In vitro control of growth and fumonisin production by Fusarium verticillioides and F. proliferatum using antioxidants under different water availability and temperature regimes. J. Appl. Microbiol. 2002, 92, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Reynoso, M.M.; Torres, A.M.; Chulze, S.N. Fusaproliferin, beauvericin and fumonisin production by different mating populations among the Gibberella fujikuroi complex isolated from maize. Mycol. Res. 2004, 108, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Palacios, S.A.; Giaj Merlera, G.; Erazo, J.; Reynoso, M.M.; Farnochi, M.C.; Torres, A.M. Trichothecene genotype and genetic variability of Fusarium graminearum and F. cerealis isolated from durum wheat in Argentina. Eur. J. Plant Pathol. 2017, 149, 969–981. [Google Scholar] [CrossRef]
- Gerlach, W.; Nirenberg, H. The genus Fusarium—A pictorial atlas. Mitteilungen aus der Biologischen Bundesanstalt fur Land- und Forstwirtschaft Berlin-Dahlem 1982, 209, 1–406. [Google Scholar] [CrossRef]
- Shephard, G.S.; Sydenham, E.W.; Thiel, P.G.; Gelderblom, W.C.A. Quantitative determination of fumonisins B1 and B2 by High-Performance Liquid Chromatography with fluorescence detection. J. Liq. Chromatogr. 1990, 13, 2077–2087. [Google Scholar] [CrossRef]
- Cooney, J.M.; Lauren, D.R.; Di Menna, M.E. Impact of competitive fungi on trichothecene production by Fusarium graminearum. J. Agric. Food Chem. 2001, 49, 522–526. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Source of Variation | df a | Growth Rates | |
|---|---|---|---|
| MS b | F c | ||
| S | 1 | 0.06 | 1.2 |
| C | 3 | 0.6 | 14 * |
| aW | 2 | 66.5 | 1364 * |
| S × C | 3 | 0.2 | 5.6 |
| S × aW | 2 | 0.07 | 1.4 |
| C × aW | 6 | 1.1 | 22 * |
| S × C × aW | 6 | 0.05 | 1.1 * |
| Source of Variation | df a | Growth Rates | |
|---|---|---|---|
| MS b | F c | ||
| S | 1 | 56.4 | 1.8 |
| C | 3 | 8933.3 | 287.5 * |
| aW | 2 | 7043.5 | 226.6 * |
| S × C | 3 | 56.3 | 1.8 |
| S × aW | 2 | 95.9 | 3.1 |
| C × aW | 6 | 1095.3 | 35.2 * |
| S × C × aW | 6 | 88.4 | 2.8 |
| Strain | Chitosan Dose (mg/g) | aW | ||
|---|---|---|---|---|
| 0.99 | 0.98 | 0.95 | ||
| F. proliferatum RC2080 | 0 | 2017 ± 638 a | 5423 ± 2028 a | 819 ± 364 a |
| 0.5 | 2747 ± 651 a | 963 ± 118 b | 611 ± 489 a | |
| 1 | 1386 ± 333 b | 637 ± 98 c | 447 ± 284 b | |
| 2 | 2064 ± 537 a | 849 ± 403 b | 214 ± 219 c | |
| F. verticillioides M7075 | 0 | 4568 ± 645 a | 8582 ± 825 a | 211 ± 181 a |
| 0.5 | 1532 ± 1311 a | 2745 ± 525 b | 409 ± 81 a | |
| 1 | 9306 ± 1487 b | 1481 ± 843 b | 343 ± 44 a | |
| 2 | 3653 ± 931 a | 442 ± 57 d | 525 ± 71 a | |
| Source of Variation | Fumonisins | Deoxynivalenol | |||
|---|---|---|---|---|---|
| df a | MS b | F c | MS b | F c | |
| S | 1 | 42,955,342.1 | 27.7 * | 518.3 | 422.3 * |
| C | 3 | 18,021,595.5 | 11.6 * | 47.2 | 38.4 * |
| aW | 2 | 55,800,484.1 | 36.0 * | 43.2 | 35.2 * |
| S × C | 3 | 5,105,970.2 | 3.3 | 27.9 | 22.7 * |
| S × aW | 2 | 19,662,834.4 | 12.7 * | 7.8 | 6.4 |
| C × aW | 6 | 21,210,509.7 | 13.7 * | 12.7 | 10.4 * |
| S × C × aW | 6 | 6,420,659.2 | 4.1 | 5.9 | 4.8 * |
| Strain | Chitosan Dose (mg/g) | aW | ||
|---|---|---|---|---|
| 0.995 | 0.99 | 0.98 | ||
| RCFG6001 | 0 | 21001 ± 1024 a | 119 ± 111 a | 177 ± 46 a |
| 0.5 | 93 ± 50 b | 148 ± 99 a | Nd b | |
| 1 | Nd c | 8.4 ± 6.8 b | Nd b | |
| 2 | Nd c | Nd c | Nd b | |
| RC22-2 | 0 | 32101 ± 1866 a | 1492 ± 489 a | 1918 ± 630 a |
| 0.5 | 16455 ± 1723 b | 2951 ± 392 b | 339 ± 68 b | |
| 1 | 5892 ± 210 c | 4094 ± 646 c | 2245 ± 564 a | |
| 2 | 8323 ± 934 c | 15466 ± 1135 d | 393 ± 201 b | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zachetti, V.G.L.; Cendoya, E.; Nichea, M.J.; Chulze, S.N.; Ramirez, M.L. Preliminary Study on the Use of Chitosan as an Eco-Friendly Alternative to Control Fusarium Growth and Mycotoxin Production on Maize and Wheat. Pathogens 2019, 8, 29. https://doi.org/10.3390/pathogens8010029
Zachetti VGL, Cendoya E, Nichea MJ, Chulze SN, Ramirez ML. Preliminary Study on the Use of Chitosan as an Eco-Friendly Alternative to Control Fusarium Growth and Mycotoxin Production on Maize and Wheat. Pathogens. 2019; 8(1):29. https://doi.org/10.3390/pathogens8010029
Chicago/Turabian StyleZachetti, Vanessa G. L., Eugenia Cendoya, María J. Nichea, Sofía N. Chulze, and María L. Ramirez. 2019. "Preliminary Study on the Use of Chitosan as an Eco-Friendly Alternative to Control Fusarium Growth and Mycotoxin Production on Maize and Wheat" Pathogens 8, no. 1: 29. https://doi.org/10.3390/pathogens8010029
APA StyleZachetti, V. G. L., Cendoya, E., Nichea, M. J., Chulze, S. N., & Ramirez, M. L. (2019). Preliminary Study on the Use of Chitosan as an Eco-Friendly Alternative to Control Fusarium Growth and Mycotoxin Production on Maize and Wheat. Pathogens, 8(1), 29. https://doi.org/10.3390/pathogens8010029